
Marialice P.C. Silvestre
Universidade Federal de Minas Gerais (UFMG), Belo Horizonte, Minas Gerais, Brazil
Mariana W.S. de Souza
Universidade Federal de Minas Gerais (UFMG), Belo Horizonte, Minas Gerais, Brazil
Carlos O. Lopes Junior
EDETEC Ind�stria Aliment�cia S/A, Belo Horizonte, Minas Gerais, Brazil
Viviane D.M. Silva
EDETEC Ind�stria Aliment�cia S/A, Belo Horizonte, Minas Gerais, Brazil
Marcos J.B. Aguiar
Universidade Federal de Minas Gerais (UFMG), Belo Horizonte, Minas Gerais, Brazil
Wendel O. Afonso
EDETEC Ind�stria Aliment�cia S/A, Belo Horizonte, Minas Gerais, Brazil
Mauro R. Silva
EDETEC Ind�stria Aliment�cia S/A, Belo Horizonte, Minas Gerais, Brazil
American Journal of Food Technology
Year: 2012 | Volume: 7 | Issue: 3 | Page No.: 123-132







ABSTRACT
Phenylketonuria (PKU) is a metabolic disease in which the untreated patients can show irreversible mental retardation and the use of milk by phenylketonurics patients is forbidden. With the aim of preparing low-phenylalanine milk hydrolysates, to be used by phenylketonurics patients with no restriction, the Activated Carbon (AC) was used as the adsorbent support. Also, the effects of certain parameters were analyzed, such as enzyme type (proteases from Aspergillus sojae and from Bacillus subtilis), temperature (30 and 50°C) and enzyme:substrate (E:S) ratio (1:100 and 2:100). The efficiency of Phenylalanine (Phe) removal was evaluated by second derivative spectrophotometry, measuring the Phe content of milk as well as in its hydrolysates after AC treatment. AC showed to be an efficient Phe adsorbent because a Phe removal from 16.28 to 58.76% was achieved, yielding certain hydrolysates with less than 100 mg Phe 100 mL-1, that could be used in the diet of phenylketonurics. The highest Phe removal (58.26%, in average) was achieved using the protease from B. subtilis at 50°C and E:S ratio of 1:100 or 2:100. At 50oC, the use of B. subtilis protease was more efficient than A. sojae protease whereas at 30°C the trend was inversed. The E:S ratio of 2:100 yielded greater Phe removal than 1:100 for B. subtilis protease at 30°C and for A. sojae at 30 and 50°C. It can be inferred that the conditions used in the study showed to be efficient in obtaining milk with reduced Phe content, appropriate for the diet of phenylketonurics.
PDF Abstract XML References Citation
Received: October 18, 2011;
Accepted: December 10, 2011;
Published: January 21, 2012
How to cite this article
Marialice P.C. Silvestre, Mariana W.S. de Souza, Carlos O. Lopes Junior, Viviane D.M. Silva, Marcos J.B. Aguiar, Wendel O. Afonso and Mauro R. Silva, 2012. Effect of Enzyme Type, Enzyme Substrate Ratio and Temperature on Phenylalanine Removal from Milk. American Journal of Food Technology, 7: 123-132.
DOI: 10.3923/ajft.2012.123.132
URL: https://scialert.net/abstract/?doi=ajft.2012.123.132
DOI: 10.3923/ajft.2012.123.132
URL: https://scialert.net/abstract/?doi=ajft.2012.123.132
INTRODUCTION
Phenylketonuria (PKU) is a metabolic disease associated with the metabolism disorder of Phenylalanine (Phe) and its treatment is mainly dietetic and based on reducing the ingestion of Phe to the lowest possible level, achieved by limitation of the protein intake (Wasserstein et al., 2006). The introduction of milk in the phenylketonurics diet is restricted and in some cases prohibited, since it is some of the food having severe controlled use for these patients (Sitta et al., 2009). Considering that milk plays an important role in the diet, one can conclude that the development of milk with low-Phe content is of great interest.
The method for removing Phe from milk involves two processes, i.e., protein hydrolysis and the use of an adsorbent. Firstly, the proteins are subjected to an enzymatic treatment aiming to expose or release Phe residues. In this way, various hydrolytic parameters and proteases had already been tested by the same group as the present work (Morato et al., 2000; Lopes et al., 2005; Morais et al., 2005; Silva et al., 2007; Soares et al., 2007; Carreira et al., 2009).
In this work two proteases were used for hydrolyzing milk proteins, one from Bacillus subtilis and the other from Aspergillus sojae. The first protease is one of subtilisins which are serine proteases. These enzymes are endopeptidases that act at neutral or alkaline pH and show a preference for cleaving esters of aromatic amino acids (Reed, 1975; Rao et al., 1998; Yang et al., 2000; Urano et al., 2001; Choi et al., 2004). The second protease has large specificity and acts at pH from 4 to 11. In addition, the protease from A. sojae is a metaloprotease activated by Zn and Co. This enzyme acts as an exopeptidase and hydrolyzes preferentially leucine bonds followed by Phe, Lys and Arg derivatives (Chien et al., 2002).
Following the enzymatic breakdown of protein molecules, Phe is removed with the aid of absorbent supports, such as resins (Delvivo et al., 2006) and the activated carbon (Lopes et al., 2005; Soares et al., 2006; Capobiango et al., 2007; Silva et al., 2007; Lopes et al., 2008; Carreira et al., 2008).
With the aim of evaluating the performance of Phe removal, this amino acid is quantified in the intact protein as well as in hydrolyzed protein after AC treatment. For this purpose, the second derivative spectrophotometry was the method chosen in the current work, because it has been previously used by the same group as the present work (Lopes et al., 2005; Capobiango et al., 2007; Lopes et al., 2007; Silva et al., 2007; Soares et al., 2007; Carreira et al., 2008; Lopes et al., 2008; Carreira et al., 2009; Silvestre et al., 2009; Silva et al., 2010; Silvestre et al., 2011a).
Aiming at the preparation of milk with low-Phe content, this work involved the study of protein hydrolysis using two enzymes (B. subtilis and A. sojae proteases) and evaluating the effect of E:S ratio and temperature.
MATERIALS AND METHODS
This study was conducted in March 2009 until June 2010 in Federal University of Minas Gerais (UFMG, Belo Horizonte, Brazil).
Materials: The UHT whole milk was acquired in a supermarket of Belo Horizonte, MG, Brazil. The proteases from Bacillus subtilis (Corolase® N, metallo-protease-EC 3.4.24.28, activity of 3.6 U mL-1, optimum pH between 6.5 and 7.5, optimum temperature between 45 and 55°C) and from Aspergillus sojae (Corolase® LAP, amino-exopeptidase-EC 3.4.11.1, activity of 63.9 U mL-1, optimum pH between 6.0 and 9.0, optimum temperature between 55 e 70°C) were kindly provided by AB Enzymes Ltd. Brazil (Barueri, SP, Brazil). One unit of activity is defined as the amount of enzyme that liberates 1 μg of tyrosine in one min at 37°C. Activated Carbon (AC) with three different particle sizes (20x50, 12x25, 6x12 mesh) was purchased from Carbomafra S.A. (Curitiba, Brazil). The other reagents were of analytical grade.
Methods
Preparation of protein hydrolysates: Eight protein hydrolysates from milk were prepared and the following parameters were varied: enzyme type, enzyme:substrate (E:S) ratio and temperature (Table 1). Initially, a volume of 50 mL of milk was placed in a flask and the pH measured (6.7).
| Table 1: | Hydrolytic conditions used for preparing milk hydrolysates |
 | |
| E:S: Enzyme substrate ratio | |
Next, it was heated at 30 and 50°C in a vaseline bath and the enzyme was added in an appropriate quantity to obtain the desired E:S ratio. After 5 h, the reaction was stopped by heating the flask in a water bath at 75°C for 15 sec in order to inactivate the enzyme which was confirmed by measuring the enzyme activity before and after the enzymatic treatment using the method described by Dias et al. (2008) without the addition of casein.
Removal of phenylalanine from protein hydrolysates: The removal of Phe from protein hydrolysates using activated carbon was described before by the same group of the present work (Silvestre et al., 2009).
Evaluation of the effect of some parameters on Phe removal: The effects of enzyme type (proteases from Aspergillus sojae and Bacillus subtilis), enzyme:substrate (E:S) ratio (1:100 and 2:100) and temperature (30 and 50°C) were evaluated in this study.
Evaluation of the efficiency of Phe removal: The efficiency of Phe removal was evaluated by measuring the free Phe in milk and in milk hydrolysates after AC treatment. Second Derivative Spectrophotometry (SDS) was used as described previously by the same group of the present work (Silvestre et al., 2009).
To generate the standard curve, stock solutions of Phe (6.05x10-4 mol L-1), Tyr (5.52x10-4 mol L-1) and Trp (4.90x10-4 mol L-1) were prepared in 0.01 mol L-1 sodium phosphate (pH 6.0). Next, 10 mL of each solution was mixed and successive dilutions of this mixture were prepared, with Phe concentrations ranging from 0.067 to 2.018x10-4 mol L-1. Spectra of these diluted solutions were recorded from 250 to 280 nm and the areas of the third negative peaks of the Phe spectra were plotted as a function of concentration. The efficiency of Phe removal was calculated according to Eq. 1:
| (1) |
Where:
| Initial Phe content | = | Phe content of milk |
| Final Phe content | = | Phe content of protein hydrolysate, after AC treatment |
Statistical analysis: All experiments were replicated three times and all measurements were carried out in triplicate. Differences between the mean areas values were evaluated by analysis of variance (ANOVA) and the Duncan test. Differences were considered to be significant at p<0.05 throughout this study. The least squares method was used to fit the standard curve and the adequacy of the linear model (y = ax+b) was tested at p<0.05. A factorial analysis was used to evaluate the effects of some parameters on Phe removal. Analysis of variance was performed for each condition to test for significant effects among treatments (p<0.05) and when significant effects were detected, the Duncan test was applied to establish the difference between the means. All the statistical analysis was conducted using the software Statistica for Windows.
RESULTS AND DISCUSSION
Efficiency of phenylalanine removal: The results of Phe removal from milk hydrolysates are shown in Table 2. They show that activated carbon was effective in removing phenylalanine from milk and the values changed from 16.28 to 58.76% and the final Phe content from 63.1 to 128.1 mg 100 mL-1 of milk. The Phe content of milk was 153 mg 100 mL-1, which is quite similar to values found in the literature, i.e., 150 mg 100 mL-1, as reported by Mapekula et al. (2011), for whole milk.
Among all tested hydrolytic conditions, six conditions (H3 to H8) resulted in milk with a low-Phe content that would be suitable for the diet of phenylketonurics, because according to Brazilian legislation the maximum amount of Phe allowed in the diet of patients is 100 mg Phe 100 mL-1 of product. It was also apparent that using the Bacillus subtilis protease with E:S ratios of 1:100 and 2:100, at 50°C resulted in the lowest Phe content. It is also worth noting that the hydrolysis of milk proteins allows them to be more quickly absorbed by the body. The hydrolyzed proteins have a lower osmolarity and are better tolerated and accepted than the mixtures of free amino acids that are commonly used in the diet of PKU patients (Frenhani and Burini, 1999).
In previous studies carried out at the same laboratory as the present study, Phe was removed from raw and skim powdered milk (Lopes et al., 2006; Soares et al., 2006). Using experimental conditions different from those of this study (raw material concentration of 0.35 g 100 mL-1), it had studied the effects of enzyme type and the method of enzyme treatment (protease from Aspergillus oryzae-AO, associated with papain-PA), as well as the E: S ratio (1:100 AO+2:100 PA, 10:100 AO+20:100 PA, for Lopes et al., 2006; Soares et al., 2006, respectively) and protein: AC ratio (1:118 and 1:90 for Lopes et al., 2006 and Soares et al., 2006, respectively). Moreover, in the study of Lopes et al. (2006), the AC was used in solution and this made it possible to remove 99% of the Phe, whereas, in the study of Soares et al. (2006), the highest value obtained was 98%.
| Table 2: | Phe removal and final Phe content of milk hydrolysates |
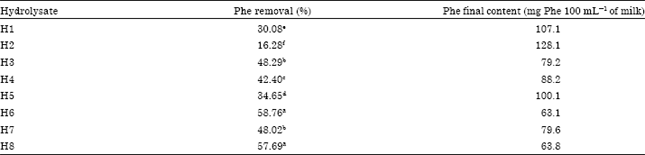 | |
Phe:Phenylalanine. Phe final content, Phe content after treatment with activated carbon. Different letters are significantly different (p<0.05) | |
These results were well above the maximum obtained in this study. It should be noted, however, that the use of a very dilute solution of raw material (24 times less concentrated than that used here) as well as large amount of AC (up to 5 times greater than the amount used here) and high E:S ratios (10 times higher than those used here) would make it economically unviable to scale up the process.
No studies were found in the literature concerning Phe removal from milk hydrolysates. Instead, certain milk proteins or milk products were reported and they were produced by using activated carbon as an adsorbent. Kitagawa et al. (1987), after hydrolyzing whey proteins with actinase at pH 6.5 and 37°C, treated these preparations with activated carbon and removed 97% of Phe. However, the conditions for treatment with this adsorbent were not described. Bajonero et al. (1991) reduced the level of Phe by 92% in hydrolysates of skim milk or sodium caseinate that were obtained by the action of papain and a protease from Aspergillus orysae. Using a mixture of three enzymes (chymotrypsin, carboxypeptidase A and leucine aminopeptidase), Moszczynski and Idziak (1993) removed 95% of Phe from casein hydrolysates. However, these authors employed more extreme conditions than those used by our group, with a very long hydrolysis time (72 h). The differences observed between these previously reported results and those obtained here may also be attributable to factors such as the type of proteolytic enzymes used, the protein: AC ratios and sources of proteins used as substrates for hydrolysis.
Activated carbon has been previously used by the same group of the present study to remove Phe from hydrolysates prepared using different protein sources, such as milk (93.6 to 99%) (Soares et al., 2006; Lopes et al., 2006), whey (75 to 99%) (Silva et al., 2007; Delvivo et al., 2006), rice grains (85 to 100%) (Lopes et al., 2008), rice flour (25.7 to 94.1%) (Silvestre et al., 2009), corn flour (68.63 to 97.55%) (Capobiango et al., 2007) and beans (25.4 to 81.5%) (Silvestre et al., 2011a). The values listed above refer to percentages of Phe content that were removed.
Effect of certain parameters on phenylalanine removal: The effects of certain parameters were evaluated based on two considerations; the ability to produce low-Phe hydrolysates and the ability to maintain low costs for scaling up the process (smaller E:S ratios and temperatures). The goal is to use the best hydrolysates obtained in the current study for the development of low-Phe milk for phenylketonurics.
Effect of enzyme type: To evaluate the influence of enzyme type on Phe removal from the hydrolysates, the samples were divided into four groups, each corresponding to different reaction conditions (E:S ratio and temperature). As shown in Fig. 1, the greatest Phe removal (58.76% in Group 2 and 57.69% in Group 4) was achieved using the protease from Bacillus subtilis at 50°C and E:S ratio of 1:100 or 2:100. This protease was much more efficient than the protease from Aspergillus sojae for two groups (2 and 4) whereas no significant difference was observed for the other two groups between the results obtained for these two enzymes.
The effect of the enzyme type on Phe removal from protein hydrolysates has not been studied by other authors. Nevertheless, the same group of the present study has carried out six studies using various protein sources and enzymes, as well as activated carbon, for removing Phe. In most of these studies, the enzyme type influenced Phe removal, as found in this study. For example, whey was hydrolyzed by a papain and a pancreatin, both immobilized on AC or alumina and it was shown that the first enzyme removed more Phe when the immobilization support was alumina (96.3 and 92.6%, respectively), whereas the opposite was observed when AC was the support (91 and 89%, respectively) (Silva et al., 2007).
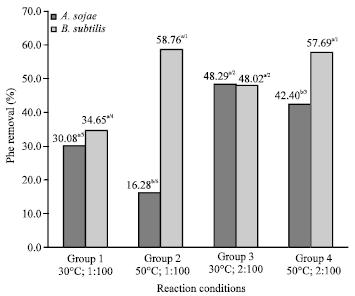 |
| Fig. 1: | Effect of enzyme type on Phe removal. Each value represents the mean of triplicate measurements. Within each group, different letters represent significantly different values (p<0.05) for different enzymes. Different numbers represent different values (p<0.05) among all samples. For all groups, the reaction time was 5 h and the Protein:Activated Carbon ration was 1:22 |
Among the seven enzymes (pancreatin, papain, proteases from Bacillus licheniformis, Aspergillus sojae, Bacillus stearothermophilus and two different ones from Bacillus subtilis used for hydrolyzing rice flour, papain was the most efficient giving rise to the greatest Phe removal (94.1%). The poorest result was obtained using B. stearothermophilus protease (69.1%; Silvestre et al., 2009). Six enzymes (the same used in the above study with rice flour, except the pancreatin) were used for the hydrolysis of bean proteins and, in this case, papain also removed the highest amount of Phe (81.5%), whereas B. subtilis protease gave the poorest results (60.8%) (Silvestre et al., 2011a).
In contrast to the results presented here, two previous studies from our group reported that the enzyme type had no influence on Phe removal. No difference was observed in the amount of Phe removed when using either a papain or a pepsin for hydrolyzing skim milk (97.6 and 97.1%, respectively) (Soares et al., 2006). Using an enzymatic extract from pineapple peel prepared in the same laboratory as the present work and a pancreatin for hydrolyzing wheat flour, no significant difference between the results obtained with both enzymes was observed (41.9 and 42.5% of Phe removal, respectively) (Carreira et al., 2009).
Effect of E:S ratio: The action of E:S ratio (1:100 and 2:100) on Phe removal from the hydrolysates is shown in Fig. 2. Samples were divided into four groups, each corresponding to different reaction conditions. As shown in Fig. 2, the E:S ratio affected Phe removal for three groups (1,2 and 3) the greatest values were achieved using an E:S of 2:100. For group 4, no significant difference was observed between the results obtained with E:S ratios of 1:100 and 2:100 which were the highest ones. It can be inferred that the advantage of using lower E:S ratio was not observed.
No other authors have reported the effect of E:S ratio on Phe removal of protein hydrolysates. However, in six studies of the same of group of the present study, this parameter was considered. In two of these studies, the results were similar to those presented here, because the advantage of using a lower E:S ratio was not observed.
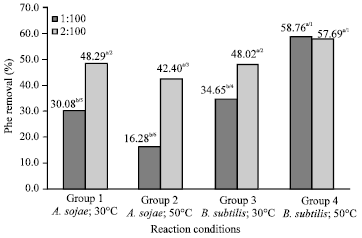 |
| Fig. 2: | Effect of E:S ratio on Phe removal. Each value represents the mean of triplicate measurements. Within each group, different letters represent significantly different values (p<0.05) for different Enzyme:Substrate ratios (E:S). Different numbers represent different values (p<0.05) among all samples. For all groups, the reaction time was 5 h and the Protein:Activated Carbon ration was 1:22 |
For example, in the study of bean protein hydrolysis using six different enzymes, the advantage of using a lower E:S ratio was not observed; the highest Phe removal was achieved with an E:S ratio of 10:100 rather than 4:100, 5:100 or 7:100 (Silvestre et al., 2011a). This was also found in case of Whey Protein Concentrate (WPC) hydrolysis using a pancreatin; the highest E:S ratio (4:100) gave the greatest Phe removal (83.1%) (Silvestre et al., 2011b).
In contrast to this study, four previous studies showed that, in some cases, there is an advantage to use a lower E:S ratio. When using a protease from Aspergillus oryzae (AO) together with a papain for hydrolyzing skim milk, it was found that reducing the E:S ratios of both enzymes from 10:100 to 1:100 for AO and from 20:100 to 2:100 for papain led to a slight improvement in Phe removal, which increased from 96 to 99% (Lopes et al., 2005).
In another example, a pancreatin was immobilized on Activated Carbon (AC) and a resin prior to the hydrolysis of whey. The advantage of using an E:S of 0.1:100 instead of 1:100 was observed only when using AC, in which case the observed Phe removal was 97 and 84% for the two E:S ratios, respectively (Delvivo et al., 2006).
In another study of whey hydrolysis, a pancreatin and a papain were immobilized on activated carbon (AC) and alumina prior to the hydrolytic reaction. The beneficial effect of E:S reduction was only observed for papain immobilized on alumina, for which an E:S of 2:100 gave better results (95% Phe removal) than an E:S of 1:100 (84% Phe removal; Silva et al., 2007). Corn flour was used as a raw material for hydrolysis by a pancreatin and in some cases it was observed the advantage of using a lower E:S ratio. Notably, when comparing E:S ratios of 1:100 and 2:100, the extents of Phe removal were 86.68 and 79.01%, respectively (Capobiango et al., 2007).
Finally, in another study, the E:S ratio had no effect on Phe removal. In this case, the E:S ratios (1:100 and 2:100) of two different pancreatins had no effect on Phe removal from rice protein hydrolysates (Lopes et al., 2008).
Effect of the temperature: The effect of the temperature on Phe removal from hydrolysates is shown in Fig. 3. Samples were divided into four groups, each corresponding to different reaction conditions (enzyme type and E:S ratio).
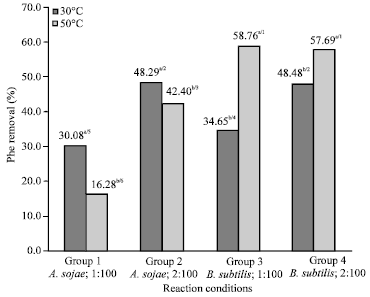 |
| Fig. 3: | Effect of the temperature on Phe removal. Each value represents the mean of triplicate measurements. Within each group, different letters represent significantly different values (p<0.05) for different temperatures. Different numbers represent different values (p<0.05) among all samples. For all groups, the reaction time was 5 h and the Protein:Activated Carbon ration was 1:22 |
The greatest Phe removal occurred at the temperature of 50°C using the B. subtillis protease and no significant difference between the percentages of Phe removal obtained for group 3 (58.76%) and 4 (57.69%) was observed. In contrast, the temperature of 30°C was more advantageous when using the A. sojae protease. In addition, the amounts of Phe removed when using these two enzymes at 30°C with an E:S of 2:100 were similar. Phe removal increased with the temperature when the B. subtillis protease was used. However, when the A. sojae protease was used, the trend was reversed, Phe removal was greater when shorter temperature was used.
No data from other authors concerning the effect of the temperature on Phe removal from protein hydrolysates were found in the literature, so the results of this study were compared with previous studies from the same group of the present work. Using a pancreatin for hydrolyzing whey proteins, it was shown that shorter temperature improved Phe removal with the best results obtained using 25°C instead of 50°C, which resulted in 83.25% Phe removal (Delvivo et al., 2006). In the study of wheat flour hydrolysis using a crude enzymatic extract (CE) from pineapple peel in association with a pancreatin, the advantage of using a shorter temperature was observed; the highest Phe removal was achieved with a temperature of 50°C (66.3%) rather than 70°C (56.9%) (Carreira et al., 2009).
CONCLUSION
The enzyme type, E:S ratio and temperature affected the Phe removal from milk. In general, the most efficient enzyme was B. subtilis protease, the E:S ratio of 2:100 was more advantageous than 1:100 and the temperature of 50°C rather than 30°C yielded the highest Phe removal. In addition, it was possible to prepare certain milk hydrolysates with reduced Phe content to be introduced in the diet of phenylketonurics.
ACKNOWLEDGMENTS
The authors thank CNPq, CAPES and FAPEMIG for their financial support.
REFERENCES
- Capobiango, M., D.C.F. Lopes, R.L. Carreira, W.O. Afonso, S.D. Segall and M.P.C. Silvestre, 2007. Optimization of enzyme assisted processes for extracting and hydrolysing corn proteins aiming phenylalanine removal. Int. J. Food Eng., 3: 1-19.
CrossRefDirect Link - Carreira, R.L., M.R. Silva, A.L.P. Starling, M.J.B. Aguiar, J.N. Januario and M.P.C. Silvestre, 2008. Association of two enzymes for obtaining low phenylalanine protein hydrolysates from wheat flour. Int. J. Food Eng., 4: 1-9.
CrossRef - Carreira, R.L., C.S. Ramos, L.A. Mundim, M.R. Silva, V.D.M. Silva and M.P.C. Silvestre, 2009. Effect of enzyme type, mode of enzyme action and temperature on the obtention of low phenylalanine hydrolysates from wheat flour. Am. J. Food Technol., 4: 71-78.
CrossRefDirect Link - Chien, H.C., L.L. Lin, S.H. Chao, C.C. Chen and W.C. Wang et al., 2002. Purification, characterization and genetic analysis of a leucine aminopeptidase from Aspergillus sojae. Biochim. Biophys. Acta (BBA): Gene Struct. Expression, 1576: 119-126.
CrossRefPubMedDirect Link - Choi, N.S., K.T. Chang, P. Jae Maeng and S.H. Kim, 2004. Cloning, expression and fibrin (ogen)olytic properties of a subtilisin DJ-4 gene from Bacillus sp. DJ-4. FEMS Microbiol. Lett., 236: 325-331.
CrossRefPubMedDirect Link - Delvivo, F.M., C.R. Vieira, E.A.R. Biasutti, M. Capobiango, V.D.M. Silva, W.O. Afonso and M.P.C. Silvestre, 2006. Evaluating the effect of adsorption medium, hydrolytic parameters and ultrafiltration on the phenylalanine removal from pancreatic whey hydrolysates. Am. J. Food. Technol., 1: 94-104.
CrossRefDirect Link - Dias, D.R., D.M. Vilela, M.P.C. Silvestre and R.F. Schwan, 2008. Alkaline protease from Bacillus sp. isolated from coffee bean grown on cheese whey. World J. Microbiol. Biotechnol., 24: 2027-2034.
CrossRefDirect Link - Frenhani, P.B. and R.B. Burini, 1999. [Mechanisms of absorption of amino acids and oligopeptides. Control and implications in human diet therapy]. Arq. Gastroenterol., 36: 227-237.
CrossRefDirect Link - Kitagawa, T., M. Owada, K. Aoki, S. Arai and T. Oura et al., 1987. Treatment of phenylketonuria with a formula consisting of low-phenylalanine peptide. Enzym, 38: 321-327.
PubMed - Lopes, D.C.F., F.M. Delvivo and M.P.C. Silvestre, 2005. Use of activated carbon for removing phenylalanine from skim milk powder. Food Sci. Technol., 38: 447-453.
CrossRefDirect Link - Lopes, D.C.F., F.M. Delvivo, J.N. Januario, M.J.B. Aguiar, A.L.P. Starling and M.P.C. Silvestre, 2007. Phenylalanine removal from whey hydrolysates. J. Food Technol., 5: 191-197.
Direct Link - Lopes, D.C.F., C.S. Bizzotto, R.L. Carreira, W.O. Afonso, C.O. Lopes Jr. and M.P.C. Silvestre, 2008. Removal of phenylalanine from protein hydrolysates prepared with rice. J. Food Technol., 6: 57-65.
Direct Link - Bajonero, L.J.L., P.L. Calderon, A.G. Mariscal, A.V. Arellano and A.L. Munguia, 1991. Enzymatic production of a low-phenylalanine product from skim milk powder and caseinate. J. Food Sci., 56: 938-942.
Direct Link - Mapekula, M., M. Chimonyo, C. Mapiye and K. Dzama, 2011. Fatty acid, amino acid and mineral composition from Nguni and local crossbred cows in South Africa. J. Food Comp. Anal., 24: 529-536.
CrossRefDirect Link - Morato, A.F., R.L. Carreira, R.G. Junqueira and M.P.C. Silvestre, 2000. Optimization of casein hydrolysis for obtaining high contents of small peptides: Use of subtilisin and trypsin. J. Food Compos. Anal., 13: 843-857.
Direct Link - Morais, H.A., L.M. De Marco, M.C. Oliveira and M.P.C. Silvestre, 2005. Casein hydrolysates using papain: Peptide profile and encapsulation in liposomes. Acta Alimentaria, 34: 59-69.
CrossRefDirect Link - Rao, M.B., A.M. Tanksale, M.S. Ghatge and V.V. Deshpande, 1998. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev., 62: 597-635.
Direct Link - Silva, V.D.M., L.M. De Marco, W.O. Afonso, D.C.F. Lopes and J.N. Januario et al., 2007. Preparation of low-phenylalanine whey hydrolysates, using papain and pancreatin immobilized on activated carbon and alumina. Am. J. Food Technol., 2: 327-341.
CrossRefDirect Link - Silva, M.C., V.D.M. Silva, T.F. Fernandes and M.P.C. Silvestre, 2010. Preparation of whey protein concentrate hydrolysates with low phenylalanine content. Nutrire: J. Brazilian Soc. Food Nutr., 35: 29-46.
Direct Link - Silvestre, M.P.C., C.O. Lopes, M.R. Silva, V.D.M. Silva, M.J.B. Aguiar and L.L. Amorin, 2011. Study of brazilian beans: Extraction and hydrolysis followed by phenylalanine removal. Am. J. Food Technol., 6: 893-903.
CrossRefDirect Link - Silvestre, M.P.C., M.C. Silva, V.D.M, Silva, M.R Silva and L.L. Amorin, 2011. WPC hydrolysates obtained by the action of pancreatin: Preparation, analysis and phelyalanine removal. Asian J. Sci. Res., 4: 302-314.
CrossRefDirect Link - Sitta, A., A.G. Barschak, M. Deon, A.T. Barden and G.B. Biancini et al., 2009. Effect of short- and long-term exposition to high phenylalanine blood levels on oxidative damages in phenylketonurics patients. Int. J. Dev. Neurosci., 27: 243-247.
CrossRefDirect Link - Soares, R.D.L., E.A.R. Biasutti, M. Capobiango, C.R. Vieira and V.D.M. Silva et al., 2006. Preparation of enzymatic skim milk hydrolysates with low phenylalanine content. Acta Farm. Bom., 25: 325-332.
Direct Link - Soares, R.D.L., M. Capobiango, E.A.R. Biasutti and M.P.C. Silvestre, 2007. Enzyme-catalyzed production of oligopeptides from skim milk. Food Biotechnol., 21: 45-56.
CrossRefDirect Link - Urano, T., H. Ihara, K. Umenara, Y. Suzuki and M. Oike et al., 2001. The profibrinolytic enzyme subtilisin NAT purified from Bacillus subtilis cleaves and inactivates plasminogen activator inhibitor type 1. J. Biol. Chem., 276: 24690-24696.
Direct Link - Wasserstein, M.P., S.E. Snyderman, C. Sansaricq and M.S. Buchsbaum, 2006. Cerebral glucose metabolism in adults with early treated classic phenylketonuria. Mol. Genet. Metabol., 87: 272-277.
CrossRef - Yang, Y., L. Jiang, L. Zhu, Y. Wu and S. Yang, 2000. Thermal stable and oxidant-resistant variant of subtilisis E. J. Biotechnol., 81: 113-118.
CrossRefDirect Link