
A. A. Sherief
Department of Botany, Faculty of Science, Mansoura University, Egypt
A. B. El-Tanash
Department of Botany, Faculty of Science, Mansoura University, Egypt
A. M. Temraz
Department of Botany, Faculty of Science, Mansoura University, Egypt
Journal of Environmental Science and Technology
Year: 2010 | Volume: 3 | Issue: 1 | Page No.: 18-34







ABSTRACT
A commercial strain of Pleurotus ostreatus was cultivated on rice straw and sawdust in plastic bags. Lignin biodegradation appeared to be doubled in case of sawdust (36%) compared with rice straw (18%). However, hemicellulose and cellulose degradation were faster in case of rice straw (66.6%), (31.2%) than saw dust (60.0%), (18.2%), respectively. Free sugars appeared to be used variably during Pleurotus ostreatus growth and residual sugars as well as total protein were considerably accumulated after fruiting. In spite of fruiting of Pleurotus ostreatus on rice straw was earlier than sawdust; however, the levels of enzyme production and activities including; exoglucanase, endoglucanase, CMCase and pectinase appeared to be higher in case of using sawdust than rice straw in both of two stages namely; mycelial and fruiting stages. On contrary, xylanase showed early rate of higher production of enzyme activity on sawdust (21.0I U g-1) during mycelial growth then sharply decreased at fruiting stage (11.0I U g-1). In addition Mn-peroxidase (MnPase) activity was more or less equal on both of the two wastes.
PDF Abstract XML References Citation
How to cite this article
A. A. Sherief, A. B. El-Tanash and A. M. Temraz, 2010. Lignocellulolytic Enzymes and Substrate Utilization During Growth and Fruiting of Pleurotus ostreatus on Some Solid Wastes. Journal of Environmental Science and Technology, 3: 18-34.
DOI: 10.3923/jest.2010.18.34
URL: https://scialert.net/abstract/?doi=jest.2010.18.34
DOI: 10.3923/jest.2010.18.34
URL: https://scialert.net/abstract/?doi=jest.2010.18.34
INTRODUCTION
Pleurotus species, commonly known as oyster mushroom, are edible cultivated world wide fungi. It characterized by its high protein content and is the easiest, fastest and cheapest to grow, require less preparation time and production technology compared with other mushrooms. Bioconversion of lignocellulosic residues through cultivation of Pleurotus species offers the opportunity to utilize renewable resources in the production of edible protein rich food (Sanchez et al., 2002). Various agricultural byproducts are being used as substrates for oyster mushroom cultivation. Some of these wastes include rice straw, wheat straw, banana leaves, peanut hull, corn leaves, mango fruits and seeds and sugarcane leaves (Cangy and Peerally, 1995; Mandeel et al., 2005). However, widely used substrates for cultivation of oyster mushroom are rice straw and sawdust (Thomas et al., 1998). Bioconversions of hard lignocellulosic components by oyster mushrooms are due to their ability to produce high levels of cellulases, hemicellulases (Vishal et al., 2006) and liginases (Tellez-Tellez et al., 2005). In parallel bioconversion technology can increase the nutritional values of wastes to use as animal fodder and increasing high values of reducing sugars able to ferment producing other sources of energy.
In Egypt, rice cultivation and wood industries results in the accumulation of large quantities of agricultural wastes rich in cellulose, hemicellulose and lignin are available with free cost throughout the year not only in farms but also in private homes. The present study aims to follow the substrate utilization and production of hydrolytic enzymes associated with mushroom development using of Pleurotus ostreatus on two different local substrates namely; rice straw and sawdust.
MATERIALS AND METHODS
Mushroom Spawns
Stock culture of P. ostreatus on millet grains was obtained from Mushroom Unit, Faculty of Agriculture, Mansoura University, Egypt. The spawn was kept at 4.0°C until used for inoculation at different experiments.
Mushroom Cultivation
Two local lignocellulosic wastes; rice straw (collected from countryside side farms of Mansoura, Dakhlia district) and yellow sawdust (Swedish wood; collected locally) were used as substrates for cultivation of P. ostreatus. A mixture of air-dried (5.0 kg) solid substrate, wheat bran (250 g) and CaCO3 (250 g) was soaked in hot tap water (70-80°C) along 3.0 h for pasteuralization and slightly decrease the level of lignin. The excess water was allowed to drain off over night. Then, CaCO3 and wheat bran as nitrogen source was added. 0.5 kg of the mixture was transferred to sterile polyethylene bag (30 cm lengthx25 cm width); however, the content occupies 20 cm length. A multilayered spawning method was applied using 15 g spawn for each bag. In this method, a layer of pasteuralized substrate was spread to a height of about 5.0 cm at the bottom of polyethylene bag. Later, a layer of spawn of about 3.0 cm was spread on the substrate until the final consisted of spawn.
Substrate Inoculation
The spawn of P. ostrearus was used to inoculate the sterile substrate under conditions (treatment were done in triplicates). Spawning was done in layers by first putting the substrate in a plastic bag and inoculating it with spawn. Another layer of substrate was place on top of that and was also inoculated with spawn. The procedure was followed until the plastic bag was full. The spawned bags were sealed and placed in a well ventilated dark room at 20±3.0°C, as mycelia growth requires no light.
After three days incubation, 25 fine holes were made into each spawned bag for proper aeration. Three replicates were harvested after 5.0 days intervals along 50 days incubation for each used substrate.
Fruiting
When the bags were fully invaded by the mycelia, they were taken to the fruiting room where temperature and humidity were controlled. Spraying the fruiting room with water raised the humidity. In addition to controlled temperature and humidity, light was also switched on to induce fruiting. Cuts were made on the bags where mushrooms were to sprout.
Determination of Lignin Content
Gravimetric determination of lignin in cultivated substrates was estimated according to Sun et al. (1996) and Adsul et al. (2005). A considerable weight of air-dried moldy substrate was fragmented into small pieces and suspended in 200 mL 1.0% (wt vol-1) aqueous solution of NaOH. The mixture was autoclaved at 121°C for 1.0 h in 500 mL Erlenmeyer flask. The residues were collected and extremely washed by tap water until neutral pH; then dried at 80°C for 48 h and weighted. The loss of weight is corresponded to lignin content.
Determination of Cellulose and Hemicellulose Content
According to (Sun et al., 1996; Badal et al., 2005) considerable weight of air dried delignified substrate (after lignin determination) was milled and screened to about 0.1 cm and suspended in 100 mL sulfuric acid 1.0% (vol vol-1). The mixture was then autoclaved at 121°C for 1.0 h in 250 mL Erlenmeyer flask. The residues were collected and washed extensively with tap water until neutral pH, dried at 80°C for 48 h and then weighted. The difference between started weight and residual weight was corresponding to the hemicellulose fractions; while, residual weight after acid hydrolysis is corresponded to cellulose content.
Extraction of Enzymes
The crude enzyme solution was obtained by soaking moldy substrate with considerable volume of 0.01 M acetate buffer (pH 5.5). The mixture was shaken for 2.0 h and centrifuged for 10.0 min at 5000 rpm to remove cells and residual substrate. The clarified extract representing as crude enzyme was used for assaying different enzyme activities.
Determination of Free Reducing Sugars
The amount of free reducing sugars in a known volume (0.1 mL) of extract media was determined as reported by Nelson (1944) and Somogyi (1952). Cultures without inoculation were used as blank to determine initial reducing sugar concentration in time zero. Free reducing sugars concentration (mM g-1 substrate) was calculated by mM standard curve of D-glucose at 700 nm using Spectro UV-VIS RS spectrophotometer (Serial No. UV-VIS 0478; Labomed Inc., USA).
Protein Determination
Protein was determined by method of Bradford (1976) by measuring optical density at 595 nm using Spectro UV-VIS RS Spectrophotometer (Serial No. UV-VIS 0478; Labomed Inc., USA). The amount of protein was calculated using μg standard curve of Bovine Serum Albumin (BSA).
Endoglucanase, CMCase and Exoglucanase Assay
The 0.1 mL of enzyme solution was incubated for 30 min at 40°C with 0.1 mL of 1.0% (wt/v) of amorphous cellulose (for endoglucanase), CMC (for CMCase) or Avicel pH 101 (for exoglucanase) dissolved in 0.1 M pH 5.5 acetate buffers. The amount of liberating free sugars was determined by Nelson (1944) and Somogyi (1952) method against boiled enzyme as control with D-glucose as standard. One unit of Endoglucanase, CMCase and Exoglucanase was defined as the amount of enzyme release of one μmole of glucose per minute per μg protein per used substrate under assay conditions.
Xylanase Assay
The 0.1 mL of crude enzyme solution was incubated with 0.1 mL of 0.5% (wt v-1) Oat-splet xylan (Biochem Co., chemicals) freshly suspended in 0.1 M pH 5.5 acetate buffer. Reaction mixture was incubated at 40°C for 30 min and then completed to 1.0 mL by adding 0.8 mL distilled water. The amount of reducing sugar released was determined by Nelson (1944) and Somogyi (1952) method against boiled enzyme using D-xylose as standard. One unit of xylanase was defined as the amount of enzyme which releases one μ mole of D-xylose per minute per μg protein per used substrate under assay conditions.
Pectinase Assay
The 0.1 mL of enzyme solution was incubated with 0.1 mL of 0.5% pectin (Citrus pectic acid; Sigma chemicals) freshly prepared by dissolving in 0.1 M pH 5.5 acetate buffer. The mixture was incubated at 40°C for 30 min. The amount of reducing sugars released was determined by Nelson (1944) and Somogyi (1952) method with D-galacturonic acid as standard. One unit of pectinase was defined as the amount of enzyme release one μ mole of reducing sugars per minute per μg protein per used substrate under assay conditions.
Manganese Peroxidase (MnPase) Assay
According to Kuwahara et al. (1984), MnPase (EC 1.11.1.13) activity was assayed using phenol red as substrate by measuring optical density at 610 nm using Spectro UV-VIS RS Spectrophotometer (Serial No. UV-VIS 0478; Labomed Inc., USA). The reaction mixture (1.0 mL) contained 500 μL enzyme extract, 100 μL phenol red solution (1.0 g L-1), 100 μL sodium lactate pH 4.5 (250 mmol L-1), 200 μL bovine serum albumin solution (0.5%), 50 μL manganese sulfate (2 mmol L-1) and 50 μL H2O2 (2 mmol L-1) in sodium succinate buffer pH 4.5 (20 mmol L-1). One unit of enzyme activity was defined as the amount of enzyme able to oxidize 1.0 μmol of substrate per minute. One control sample was tested using all the method conditions without manganese in order to demonstrate the manganese peroxides.
Statistical Analysis
Statistical software; Graph-Pad Prism 4.0 (San Diego, USA) is used in regression analysis and plotting all figures shown in our study.
RESULTS AND DISCUSSION
Cultivation of mushroom presents an economically important biotechnological industry that has markedly developed all over the world. It is estimated that more than 10 million metric ton of edible and medicinal mushrooms were produced in 2004 in various countries (Royse, 2005). Mushroom can convert the huge lignocellulosic wastes into a wide diversity of products (edible or medicinal food, feed and fertilizers), protecting and regenerating the environment.
Many literatures were previously studied the growth and the yield of different edible mushrooms; however, scattered researches studied their ability for different lignocellulases production. Great potential for using different local lignocelluloses as raw materials was used for the cultivation and production of P. ostreatus (Pankow, 1984; Zadrazil, 1987; Gapinski and Ziombra, 1988).
Pleurotus ostreatus was cultivated on considerable weight (0.5 kg bag -1) of natural rice straws and saw dust at 23°C along 50 days. Three sets of cultivated bags were harvested after 5.0 days intervals. Biodegradable lignin, cellulose and hemi-cellulose were estimated. Furthermore, relative lignocelluloses patterns associated with mushroom development were also detected. Practically, the incubation time was divided into two main periods; the first (5-20 days) is mycelial growth period; while the second period (20-50 days) considered as fruiting period.
The results showed that the rate of growth, mushroom yield and the diameter of its fruiting bodies were varied from rice straw to saw dust. In addition; Leatham (1982) detected considerable out-layers mycelial growth of P. ostreatus on wood after 3.0 days, while; complete mycelium network was highly permeated the medium after 15 days incubation; then, rapid mushroom growth and maximum lignocellulases pattern was occurred in the range of 15-45 days incubation.
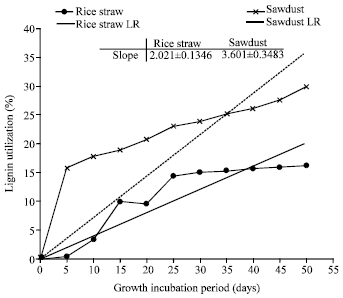 | |
| Fig. 1: | Utilization of lignin by P. ostreatus grown on rice straw and sawdust |
According to lignin pattern Fig. 1, results indicated that lignin degradation by Pleurotus ostreatus was gradually increased with incubation time on both rice straw and saw dust. The 30 and 16.3% loss of lignin was estimated after 50 days for sawdust and rice straw, respectively. However, linear regression analysis of data indicated that the rate of lignin degradation in case of sawdust (3.60±0.35% h-1) was higher than that of rice straw (2.02±0.14% h-1). These results are in agreement with those obtained by Albores et al. (2006), who reported a decrease in substrate dry weight with incubation time. The loss of initial lignin could be used as indication for substrate degradation caused by extra-cellular enzymes produced from P. ostreatus. Similar patterns of lignocellulosic components utilization was reported by Mukherjee and Nandi (2004) during degradation of water hyacinth biomass by P. citrinopileatus after 48 days. Furthermore, Lechner and Papinutti (2006) reported reduction of lignin and cellulose content of wheat straw to 21.49 and 53.26%, respectively.
Lignin degradation could be attributed to the ability of P. ostreatus to bulk of ligninases production such as laccases and peroxidases (Leonowicz et al., 1999; Baldrian et al., 2005; Hoegger et al., 2007). In addition, Ortega et al. (1992) refereed that strong ligninolytic activity by Pleurotus on sugarcane and wheat straw. Furthermore, the rate of lignin degradation was highly depending on used substrate and the value of Mn+2 in used substrate or other additive nutrients. About 20% of lignin from cotton stalks was estimated by P. ostreatus after 30 days incubation; while, 50-56% lignin of cotton stalks amended with Mn+2 was degraded by P. ostreatus after the same fermentation period (Kerem and Hadar, 1995). Mn+2 ions are a crucial substrate for Manganese Peroxidase (MnP) which considered the most abundant extra-cellular ligninolytic enzyme (Hataka, 1994). Ruhl et al. (2008) found that P. ostreatus may use as industrial strain for MnP production when cultivated on wheat straw. Furthermore, the preferring properties of lignin utilization by white rot fungi were industrially used in delignification and animal feed (Reddy, 2001).
The rates of cellulose degradation Fig. 2 are directly proportional with fermentation periods on both used substrates. However, the rate of cellulose utilization by P. ostreatus grown on rice straw (3.28±0.33 h-1) was higher as compared with its growth on sawdust (1.62±0.17 h-1).
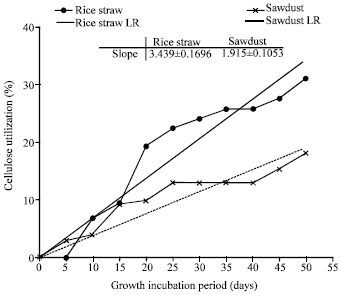 | |
| Fig. 2: | Utilization of cellulose by P. ostreatus grown on rice straw and sawdust |
The maximum values of cellulose consumption reached 31.2 and 18.2% of rice straw and saw dust respectively after 50 days incubation. Consequently, degradation of cellulose followed a trend similar to that observed from lignin and hemicellulose fractions. Aracelly-Vega et al. (2005) also reported reduction in cellulose content of coffee pulp and banana leaves after P. ostreatus cultivation. Cellulose consumption may due to the ability of P. ostreatus for different inducible cellulases production. The action of cellulases (exoglucanase, endoglucanase and cellobiase) enables fungus to degrade cellulose fraction in used substrates (Datta and Chakravarty, 2001). Higher CMCase and FPase are detected from P. ostreatus and P. sajor-caju cultivated on banana waste along 40 days incubation (Reddy et al., 2003). Cellulose degradation pattern in our results are in agreement with that obtained by Alemawor et al. (2009) who reported maximal cellulose degradation when P. ostreatus cultivated on cocoa pod husk after 35 days.
The results Fig. 3 showed that the residual hemi-cellulose was highly decreased along incubation periods in both used substrates. The rate of hemicelluloses degradation reached the maximum 66.6 and 60.0% after 50 days incubation in rice straw and saw dust, respectively.
However, the rate of initial hemi-cellulose degradation in rice straw (6.32±0.22 h-1) was higher compared with saw dust (4.95±0.56 h-1). Reduction of hemicellulose content with time may be an indication with the ability of P. ostreatus for hemicellulases (xylanase, xylosidase, arabinase and pectinase) production on used wastes. Significant hemicellulolytic activities by Pleurotus on different lignocelluosic materials have also been reported (Buswell et al., 1993). However, De-Menezes et al. (2009) showed that the production ability of hemicellulases (β-xylanase and β-xylosidase) by Pleurotus sp., BCCB068 and P. tailandia utilized 75.1 and 73.4% of xylan, respectively after 40 days incubation. In addition, Alemawor et al. (2009) reported that 20.5% of hemicellulose reduction after 35 days incubation when P. ostreatus cultivated on cocoa pod husk Protein pattern Fig. 4 showed gradually and significantly increase in total unspecified protein production by P. ostreatus and correlated positively with fermentation periods. The value of protein reached the maximum after 40 days incubation for rice straw (19.6 mg g-1) and after 45 days incubation for sawdust (15.9 mg g-1); however, the maximum values were recorded at fruiting period.
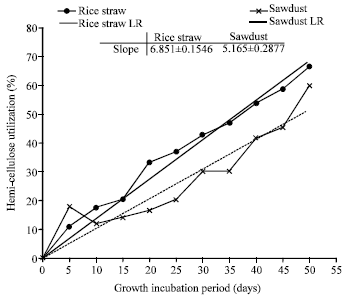 | |
| Fig. 3: | Utilization of hemi-cellulose by P. ostreatus grown on rice straw and sawdust |
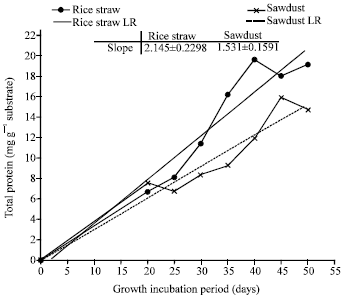 | |
| Fig. 4: | Total protein produced by P. ostreatus grown on rice straw and sawdust |
These results are very important in application field to use rice straw as fodder. Similar results were obtained by Membrillo et al. (2008) who showed increasing 48% of total protein to sugarcane bagasse when cultivated with P. ostreatus. The observed increase in protein content during mushroom growth and fruiting period indicates a positive substrates bioconversion (Alemawor et al., 2009). In this connection, Iyayi (2004) referred that protein produced from fungal bioconversion process of soluble carbohydrates was colonized into mycelial protein or single cell protein. Furthermore, edible mushrooms have been reported to be capable of transforming nutritionally low-grade agro-wastes into protein-rich byproducts which are of high biological value as they are rich in essential amino acids (Alofe et al., 1998; Manzi et al., 1999). In addition, Most of the extracellular fungal enzymes produced by the growing fungus during bioconversion process are proteinaceous in nature; thus spent enzymes could contribute some amount of protein to the substratum (Kadari, 1999).
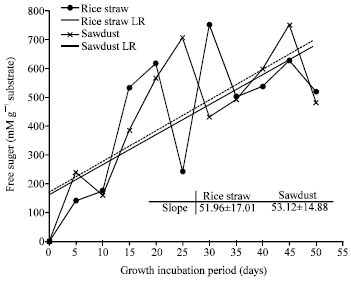 | |
| Fig. 5: | Free reducing sugar produced by P. ostreatus grown on rice straw and sawdust |
Earlier studies of mushroom growth on cassava byproducts, wheat straw, coffee husk, corn bran and rice bran have also reported similar increases in protein content (Leifa et al., 2001; Iyayi and Aderolu, 2004; Das and Mukherjee, 2007).
Results in Fig. 5 showed that the production of reducing sugars by P. ostreatus detected highly after 30 days incubation in case of rice straw (753.8 mM g-1) and after 45 days incubation for saw dust (752.2 mM g-1). The total soluble sugars on both substrates were increased and dramatically correlated directly with fermentation period. Lower values of liberated sugars observed in the 1st harvest of fermentation followed by significant increase with next harvests. Increasing the free sugars may attributed to hydrolytic enzyme efficiency of P. ostreatus for degradation of cellulose, hemicellulose and pectin fractions in used substrates. Similar results were also obtained by P. ostreatus when cultivated on cocoa pod husk composition (Alemawor et al., 2009). Furthermore, xylooligosaccharides, glucose, xylose, arabinose, cellobiose, mannose and maltose were observed due to the biodegradation of hemicellulose residues by Pleurotus sp., BCCB068 and P. tailandia strains (De-Menezes et al., 2009).
Since, the carbon sources are usually of lignocellulosic character, the mushroom during vegetative growth and fruiting period produce a wide range of enzymes to degrade the lignocellulosic substrates: peroxidases and laccases for lignin degradation and various types of glucanases, cellulases and xylanases for cellulose and hemicellulose degradation (De Groot et al., 1998). Otherwise, changes in enzyme activities occur during fruiting, indicating a connection to the regulation of fruiting body development. For example, A. bisporus and L. edodes laccase activities are highest just before fruiting initiation and decline rapidly with fruiting bodies formation; while, cellulases activities are highest when fruiting bodies develop (Ohga, 1992; De Groot et al., 1998).
Depending on the component fractions of used rice straw and sawdust; six recognized extra-cellular ligno-cellulases associated with growth and fruiting of mushroom including; exoglucanase, endoglucanase, CMCase, pectinase, Mn-peroxidase and xylanase were examined. Results showed that all enzymes were highly produced at different incubation periods of P. ostreatus grown on rice straw and sawdust. However, sawdust was the most suitable medium for different enzymes productivity compared with rice straw under the same fermentation conditions.
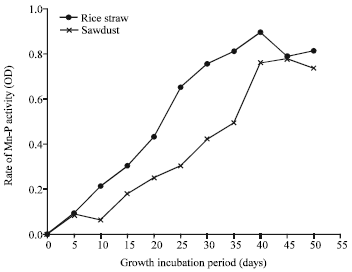 | |
| Fig. 6: | MnPase profile of P. ostreatus grown on rice straw and sawdust |
Low levels of different enzyme activities were detected at the first harvest. Furthermore, the optimum values of different enzyme activities were varied according enzyme and the type of substrate.
Present results Fig. 6 showed that the rate of P. ostreatus MnPase production was gradually increased with incubation time reaching the optimum values of activity after 40 days incubation in both substrates. White rot fungi can degrade lignin to reach other more readily available carbon sources present in lignocellulosic materials. On other hand Shashirekta and Rajarathnam (2007) reported higher lignin degradation in a period coinciding with maximum ligninase productivity during fruiting period. Due to the dissolution of lignin, the substrate turned dark brown, at the end of mushroom cropping period (Wainright, 1992). Different mushrooms are able to utilize lignin through lignin degradation by ligninase systems. However, commercial P. ostreatus was able to produce different ligninases such as; laccase, MnP and versatile peroxidase (VP) when cultivated on wheat straw (Rühl et al., 2008). Literatures have been also described the ability of different mushroom for lignin degradation and ligninase production (Eichlerova et al., 2000; Savoie et al., 2007). Similar results for MnPase pattern and other ligninases were recorded by different Pleurotus species cultivated on sugarcane bagasse, wheat straw and tree leaves (Mata et al., 2007; Elisashvili et al., 2008).
For cellulases; the results Fig. 7 indicated that the optimal exoglucanase was produced on the both substrate after 20 days, reached 4.02 and 3.87 IU g-1 on rice straw and sawdust respectively. However, endoglucanase Fig. 8 was optimally produced after 30 days of fungus growth on saw dust (6.01 IU g-1) and after 45 days incubation for rice straw (3.76 IU g-1). On the other hand, CMCase Fig. 9 reached the optimal value of productivity (13.2 IU g-1) in sawdust after 20 days. Similar pattern were obtained for the maximum cellulases productivity by P. ostreatus and P. sajor-caju cultivated on leaf biomass and pseudo-stem biomass of banana (Reddy et al., 2003).
Low levels of P. ostreatus CMCase when P. ostreatus cultivated on rice straw comparing with saw dust may explain the higher growth and mushroom yield on sawdust. Similar levels of lower CMCase productivity was reported for other Pleurotus sp., grown on rice straw (Rai and Saxena, 1990) as well as banana biomass (Reddy, et al., 2003). On the other hand, no cellulolytic enzyme activity from P. sajor-caju and Lentinula edodes was detected on sawdust culture supernatants with crystalline cellulose (Buswell et al., 1993).
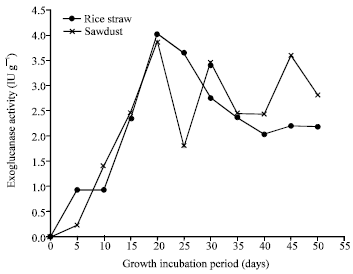 | |
| Fig. 7: | Exoglucanase profile of P. ostreatus grown on rice straw and sawdust |
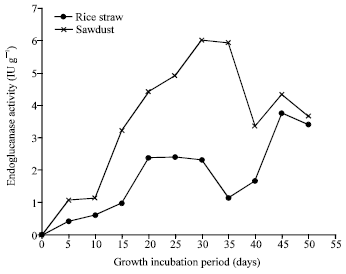 | |
| Fig. 8: | Endoglucanase profile of P. ostreatus grown on rice straw and sawdust |
This study, lowest endoglucanase activities compared to CMCase were recorded when P. ostreatus grown on rice straw or sawdust. The low endoglucanase activity limits the rate at which white rot fungi degrade native cellulose. The action is necessary for the degradation of highly ordered (crystalline) forms of cellulose where it acts synergistically with CMCase active enzymes (MacKenzie et al., 1984; Leatham, 1985).
High CMCase production was observed in case of sawdust and lower productivity was detected in case of rice straw before fruiting period. After mushroom fruiting, CMCase was considerably decreased. The patterns of endoglucanase and exoglucanase were completely different comparing with CMCase patterns. However, maximum endoglucanase (in case of sawdust, 6.01 IU g-1 after 30 days; in case of rice straw 3.76 IU g-1 after 45 days) and exoglucanase (after 20 days in both cases of saw dust 3.87 IU g-1 and rice straw 4.02 IU g-1) productivity were observed at fruiting period in both cases. Similar results for high cellulase productivity during the fruiting phase by Pleurotus sp., was also obtained by Avneesh et al. (2003).
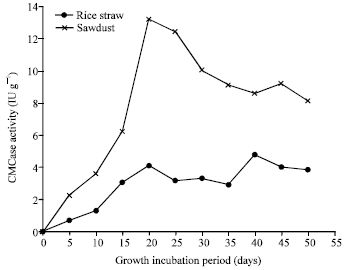 | |
| Fig. 9: | CMCase profile of P. ostreatus grown on rice straw and sawdust |
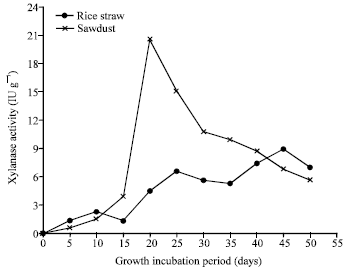 | |
| Fig. 10: | Xylanase profile of P. ostreatus grown on rice straw and sawdust |
In xylanase pattern, the results Fig. 10 showed early and highly levels of xylanase production compared with other lignocellulases in case of sawdust. P. ostreatus xylanase was recorded as optimum after 20 days in case of sawdust (20.64 IU g-1) and after 45 days in case of rice straw (8.94 IU g-1). In earlier studies, optimum xylanase production was detected after 16 days by P. ostreatus grown on wheat straw (Garzillo et al., 1994); as well as, corncob contains high level of xylan (33%) has been used as an induction substrate for different xylanases production by P. ostreatus (Qinnghe et al., 2004). However, low levels of xylanase production were detected from other P. ostreatus 3004 cultivated on corncobs (Sermanni et al., 1994). Significant higher xylanase was produced from Pleurotus sp., BCCB068 throughout the sampling period corresponding to the 20th, 30th and 40th days; while, P. tailandia produced lower values of xylanase at the same culture conditions.
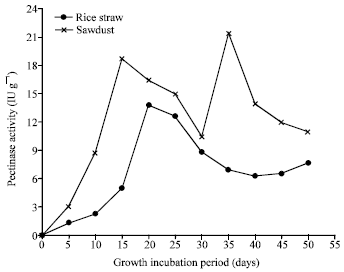 | |
| Fig. 11: | Pectinase profile of P. ostreatus profile grown on rice straw and sawdust |
De Menezes et al. (2009) attributed these results to β-xylosidase, they detected higher β-xylosidase in case of Pleurotus sp., BCCB068; while, P. tailandia did not produce detectable β-xylosidase except in 10th day.
The results also showed that xylanase productivity was highly increased in case of sawdust before P. ostreatus fruiting period. It reached the maximum value (20.64 IU g-1) at the end of growth period (20 days incubation). However, after mushroom fruiting, xylanase activity decreased 1.4-3.7 times over the values obtained before fruiting. These results are different with that obtained in case of rice straw. However, the rate of xylanase production was slightly increased in fruiting compared with growth period; it reached the maximum value (8.94 IUg-1) after 45 days incubation. Lower productivity of xylanase and other lignocellulases in case or rice straw may be attributed to the higher content of solid compounds such as silica (Van Hoest, 2006). The results of xylanase pattern on rice straw is positive correlated with that results obtained by Isikhuemhen and Mikiashvilli (2009), who observed higher xylanase productivity from P. ostreatus in fruiting than mycelial growth period when cultivated on some solid wastes. However, similar results for xylanase supported our data of xylanase activity on rice straw (Rajarathnam, 1981; Terashita et al., 1998). They observed that xylanase activity increased during vegetative mycelial growth and reached its maximum activity after cropping of P. flabellatus and P. sajor-caju.
Results of Fig. 11 showed that P. ostreatus pectinase was optimally produced (21.42 IU g-1) after 35 days in case of saw dust and after 20 days incubation (13.80 IU g-1) on rice straw. Although, pectinase production by P. ostreatus when cultivated on rice straw and sawdust has not been studied previously; however, we studied this enzyme because of the highly importance and its widely used in food processing industries (tissue maceration, juice extraction and clarification in the fruit and vegetable processing industry) as reported from literatures (Cannel and Moo-Young, 1980; Dosanjh and Hoondal, 1996). Lentinus edodes has highly advantage of possessing status, which permits the use of its metabolites including extra-cellular pectinase in food processing industry (Pariza and Foster, 1983). It was found that a high level of pectinase activity was produced by L. edodes during cultivation on strawberry pomace. Furthermore, Zheng and Shetty (2000) showed that it is feasible to use fruit processing wastes such as strawberry pomace as a raw material for production highly thermal and pH stable pectinase by food-grade fungus L. edodes. Higher pectinase productivity were obtained from four Egyptian food processing wastes (orange peel, lemon peel, apple pomace and sugar cane bagasse) during cultivation of P. ostreatus NRRL-0366. However, saccharification of some pretreated lignocellulosic wastes as well as clarification of some fruit juices using crude culture filtrates of P. ostreatus showed promising results (Rashad et al., 2009).
CONCLUSION
In conclusion, the results indicate that commercial P. ostreatus are able to grow on local rice straw and sawdust residues causing considerable degradation efficiency for different lignocelluloses fraction by producing a bulk of inducible lignocellulases. Significant variation in biodegradation and levels of enzyme productivity was done according to used substrate. The production of high enzymatic activities in both growth and fruiting periods did not necessarily result of large extent of a specific substrate component decays. On the other hand, lignin, cellulose and hemicellulose utilization confirmed to be key processes for rice straw and sawdust degradation. This indicates that the de-lignification, softness, protein richness and free sugar contents after cultivation of P. ostreatus increases the availability of rice straw to use for animal feed.
REFERENCES
- Albores, S., M.J. Pianzzola, M. Soubes and M.P. Cerdeiras, 2006. Biodegradation of agroindustrial wastes by Pleurotus spp. for its use as ruminant feed. Elect. J. Biotechnol., 9: 215-220.
Direct Link - Adsul, M.G., J.E. Ghule, H. Shaikh, R. Singh, K.B. Bastawde, D.V. Gokhale and A.J. Varma, 2005. Enzymatic hydrolysis of delignified bagasse polysaccharides. Carbohydr. Polymer, 62: 6-10.
CrossRefDirect Link - Alemawor, F., V.P. Dzogbefia, E.O.K. Oddoye and J.H. Oldham, 2009. Effect of Pleurotus ostreatus fermentation on cocoa pod husk composition: Influence of fermentation period and Mn2+ supplementation on the fermentation process. Afr. J. Biotechnol., 8: 1950-1958.
Direct Link - Saha, B.C., L.B. Iten, M.A. Cotta and Y.V. Wu, 2005. Dilute acid pretreatment, enzymatic saccharification and fermentation of wheat straw to ethanol. Process Biochem., 40: 3693-3700.
CrossRefDirect Link - Baldrian, P., V. Valaskova, V. Merhautova and J. Gabriel, 2005. Degradation of lignocellulose by Pleurotus ostreatus in the presence of copper, manganese, lead and zinc. Res. Microbiol., 156: 670-676.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Buswell, J.A., Y.J. Cai and S.T. Chang, 1993. Fungi and Substrate Associated Factors Affecting the Ability of Individual Mushroom Species to Utilize Different Lignocellulosic Growth Substrates. In: Mushroom Biology and Mushroom Products, Chang, S.T., J.A. Buswell and S.W. Chiu (Eds.). Chinese University Press, Hong Kong, pp: 141-150.
- Das, N. and M. Mukherjee, 2007. Cultivation of Pleurotus ostreatus on weed plants. Bioresour. Technol., 98: 2723-2726.
CrossRefDirect Link - De Groot, J., J. Visser, V. Griensven and J. Schaap, 1998. Biochemical and molecular aspects of growth and fruiting of the edible mushroom Agaricus bisporus. Mycol. Res., 102: 1297-1308.
Direct Link - De Menezes, C.R., I.S. Silva, E.C. Pavarina, E.F.G. Dias, F.G. Dias, M.J. Grossman and L.R. Durrant, 2009. Production of xylooligosaccharides from enzymatic hydrolysis of xylan by the white-rot fungi Pleurotus. Int. Biodeterioration Biodegradation, 63: 673-678.
CrossRef - Dosanjh, N.S. and G.S. Hoondal, 1996. Production of constitutive, thermostable, hyper active exo-pectinase from Bacillus GK-8. Biotechnol. Lett., 18: 1435-1438.
Direct Link - Eichlerova, I., L. Homolka, F. Nerud, F. Zadrazil, P. Baldrian and J. Gabriel, 2000. Screening of Pleurotus ostreatus isolates for their ligninolytic properties during cultivation on natural substrates. Biodegradation, 11: 279-287.
Direct Link - Elisashvili, V., M. Penninckx, E. Kachlishvili, N. Tsiklauri, E. Metreveli, T. Kharziani and G. Kvesitadze, 2008. Lentinus edodes and Pleurotus species lignocellulolytic enzymes activity in submerged and solid-state fermentation of lignocellulosic wastes of different composition. Bioresour. Technol., 99: 457-462.
CrossRefDirect Link - Garzillo, A.M.V., S. Di Paolo, M. Ruzzi and V. Buonocore, 1994. Hydrolytic properties of extracellular cellulases from Pleurotus ostreatus. Applied Environ. Microbiol., 42: 476-481.
Direct Link - Hatakka, A., 1994. Lignin-modifying enzymes from selected white-rot fungi: Production and role from in lignin degradation. FEMS Microbiol. Rev., 13: 125-135.
CrossRefDirect Link - Isikhuemhen, O.S. and N.A. Mikiashvilli, 2009. Lignocellulolytic enzyme activity, substrate utilization, and mushroom yield by Pleurotus ostreatus cultivated on substrate containing anaerobic digester solids. J. Ind. Microbial Biotechnol., 36: 1353-1362.
CrossRef - Kerem, Z. and Y. Hadar, 1995. Effect of manganese on preferential degradation of lignin by Pleurotus ostreatus during solid-state fermentation. Applied Environ. Microbiol., 61: 3057-3062.
Direct Link - Leatham, G.F., 1982. Cultivation of shiitake, the Japanese forest mushroom, on logs: A potential industry for the United States. Forest Prod. J., 32: 29-35.
Direct Link - Leatham, G.F., 1985. Extracellular enzymes produced by the cultivated mushroom Lentinus edodes during degradation of a lignocellulosic medium. J. Applied Environ. Biol., 50: 859-867.
Direct Link - Lechner, B.E. and V.L. Papinutti, 2006. Production of lignocellulosic enzymes during growth and fruiting of the edible fungus Lentinus tigrinus on wheat straw. Process Biochem., 41: 594-598.
CrossRefDirect Link - Leifa, F., A. Pandey and C.R. Soccol, 2001. Production of flammulina velutipes on coffee husk and coffee spent-ground. Braz. Arch. Biol. Biotechnol., 44: 205-212.
CrossRefDirect Link - Leonowicz, A., A. Matuszewska, J. Luterek, D. Ziegenhagen and M. Wojtas-Wasilewska et al., 1999. Biodegradation of lignin by white rot fungi. Fungal Genet. Biol., 27: 175-185.
CrossRef - Iyayi, E.A., 2004. Changes in the cellulose, sugar and crude protein contents of agro-industrial by-products fermented with Aspergillus niger, Aspergillus flavus and Penicillium sp. Afr. J. Biotechnol., 3: 186-188.
Direct Link - Iyayi, E.A. and Z.A. Aderolu, 2004. Enhancement of the feeding value of some agro-industrial by-products for laying hens after their solid state fermentation with Trichoderma viride. Afr. J. Biotechnol., 3: 182-185.
CrossRefDirect Link - MacKenzie, C.R., D. Bilous and K.G. Johnson, 1984. Purification and characterization of an exoglucanase from Streptomyces flavogriseus. Can. J. Microbiol., 30: 1171-1178.
Direct Link - Mandeel, Q.A., A.A. Al-Laith and S.A. Mohamed, 2005. Cultivation of oyster mushrooms (Pleurotus spp.) on various lignocellulosic wastes. World J. Microbiol. Biotechnol., 21: 601-607.
CrossRefDirect Link - Mata, G., E. Cortes and D. Salmones, 2007. Mycelial growth of three Pleurotus (Jacq.: Fr.) P. Kumm species on sugarcane bagasse: Production of hydrolytic and oxidative enzymes. Int. J. Med. Mushrooms, 9: 385-394.
Direct Link - Membrillo, I., C. Sanchez, M. Meneses, E. Favela and O. Loera, 2008. Effect of substrate particle size and additional nitrogen source on production of lignocellulolytic enzymes by Pleurotus ostreatus strains. Bioresour. Technol., 99: 7842-7847.
Direct Link - Mukherjee, R. and B. Nandi, 2004. Improvement of in vitro digestibility through biological treatment of water hyacinth biomass two Pleurotus species. Int. Biodeter. Biodegr., 53: 7-12.
Direct Link - Ortega, G.M., E.O. Martinez, D. Betancourt, A.E. Gonzalez and M.A. Otero, 1992. Bioconversion of sugarcane crop residues with white-rotfungi Pleurotus sp. World J. Microbiol. Biotechnol., 8: 402-405.
Direct Link - Cai, Q., X. Yue, T. Niu, C. Ji and Q. Ma, 2004. The screening of culture condition and properties of xylanase by white-rot fungus Pleurotus ostreatus. Process Biochem., 39: 1561-1566.
CrossRef - Rashad, M.M., H.M. Abdou, W.G. Shousha, M. Aliand and N.N. El-sayed, 2009. Utilization of some food processing wastes for production of Pleurotus ostreatus pectinases. Adv. Food Sci., 31: 151-157.
Direct Link - Sanchez, A., F. Ysunza, M.J. Beltran-Gracia and M. Esqueda, 2002. Biodegradation of viticulture wastes by Pleurotus: A source of microbial and human food and its potential use in animal feeding. J. Agric. Food Chem., 50: 2537-2542.
Direct Link - Savoie, J.M., D. Salmones and G. Mata, 2007. Hydrogen peroxide concentration measured in cultivation substrates during growth and fruiting of the mushrooms Agaricus bisporus and Pleurotus spp. J. Sci. Food Agric., 87: 1337-1344.
Direct Link - Sermanni, G., A. Annibale, D. Lena, G. Vitale, N.S. Di and E. Mattia, 1994. The production of exo-enzymes by Lentinus edodes and Pleurotus ostreatus and their use for upgrading corn straw. Bioresour. Technol., 48: 173-178.
Direct Link - Shashirekha, M.N. and S. Rajarathnam, 2007. Bioconversion and biotransformation of coir pith for economic production of Pleurotus florida: Chemical and biochemical changes in coir pith during the mushroom growth and fructification. World J. Microbiol. Biotechnol., 23: 1107-1114.
CrossRefDirect Link - Avneesh, D.S., N. Abdullah and S. Vikineswary, 2003. Optimization of extraction of bulk enzymes from spent mushroom compost. J. Chem. Technol. Biotechnol., 78: 743-752.
CrossRef - Sun, R., J.M. Lawther and W.B. Banks, 1996. Fractional and structural characterization of wheat straw hemicelluloses. Carbohydr. Polym., 29: 325-331.
CrossRefDirect Link - Tellez-Tellez, M., C. Sanchez, O. Loera and G. Diaz-Godinez, 2005. Differential patterns of constitutive intracellular laccases of the vegetative phase of Pleurotus species. Biotechnol. Lett., 27: 1391-1394.
Direct Link - Terashita, T., R. Murao, K. Yoshikawa and J. Shishiyama, 1998. Changes in carbohydrase activities during vegetative growth and development of fruit-bodies of Hypsizygus marmoreus grown in sawdust-based culture. J. Wood Sci., 44: 234-236.
Direct Link - Thomas, G.V., S.R. Prabhu, M.Z. Reeny and B.M. Bopaiah, 1998. Evaluation of lignocellulosic biomass from coconut palm as substrate for cultivation of Pleurotus sajor-caju (Fr.) Singer. World J. Microbiol. Biotechnol., 14: 879-882.
CrossRefDirect Link - Van Soest, P.J., 2006. Rice straw, the role of silica and treatments to improve quality. Anim. Feed Sci. Technol., 130: 137-171.
CrossRefDirect Link - Vishal, S., B. Petr, E. Ivana, D. Rachna, M. Datta, N. Frantisek and G. Richard, 2006. Influence of dimethyl sulfoxide on extracellular enzyme production by Pleurotus ostreatus. Biotechnol. Lett., 28: 651-655.
Direct Link - Reddy, G.V., P. Ravindra Babu, P. Komaraiah, K.R.R.M. Roy and I.L. Kothari, 2003. Utilization of banana waste for the production of lignolytic and cellulolytic enzymes by solid substrate fermentation using two Pleurotus species (P. ostreatus and P. sajor-caju). Process Biochem., 38: 1457-1462.
CrossRefDirect Link