
M.B. Achenef
Faculty of Veterinary Medicine, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
A.K. Arifah
Faculty of Veterinary Medicine, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
Y.M. Goh
Faculty of Veterinary Medicine, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
A.Q. Sazili
Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
O. Fauziah
Faculty of Medicine and Health Science, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
A. Zuraini
Faculty of Medicine and Health Science, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
M.N. Somchit
Faculty of Medicine and Health Science, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
Z.A. Zakaria
Faculty of Pharmacy, Universiti Teknologi Mara, 40450 Shah Alam, Selangor, Malaysia
Journal of Biological Sciences
Year: 2009 | Volume: 9 | Issue: 6 | Page No.: 567-572







ABSTRACT
This study was focused on the quantitative analysis of Conjugated Linoleic Acids (CLAs) in rumen liquor and assessment of the relationship between CLAs and other carbon 18 fatty acids and rumen pH of cattle slaughtered for meat consumption. The fatty acids were extracted using modified Folch method and determined by gas chromatography. The mean (n = 22) level of CLAs in the rumen liquor was 14.97 ±7.46 mg/100 mL. Cis-9, trans-11(c9, t11) was the dominant isomer of CLA followed by trans-10,cis-12 (t10, c12). The level of CLAs was positively correlated with the concentration of octadecatrienoic (18:3), cis-9, cis-12 octadecadienoic (18:2), trans-11octadecenoic (18:1) and octadecenoic (18:0) acids. Ruminal pH was positively correlated with the amount of c9, t11 and total CLA and negatively correlated with t10, c12 CLA isomer. All these correlations and relationships indicated that CLAs production in the rumen is dependent on availability of carbon 18 fatty acids and pH of the rumen liquor.
PDF Abstract XML References Citation
How to cite this article
M.B. Achenef, A.K. Arifah, Y.M. Goh, A.Q. Sazili, O. Fauziah, A. Zuraini, M.N. Somchit and Z.A. Zakaria, 2009. Conjugated Linoleic Acid in Rumen Liquor: Their Relation with Other Carbon 18 Fatty Acids and Rumen pH. Journal of Biological Sciences, 9: 567-572.
DOI: 10.3923/jbs.2009.567.572
URL: https://scialert.net/abstract/?doi=jbs.2009.567.572
DOI: 10.3923/jbs.2009.567.572
URL: https://scialert.net/abstract/?doi=jbs.2009.567.572
INTRODUCTION
Rumen is a large fermentation vat where bacteria, protozoa and fungi live and form a symbiotic relationship to facilitate digestion of plant fibers. These microbial populations play a role for the formation of Conjugated Linoleic Acids (CLAs). Conjugated linoleic acids are groups of positional and geometric isomers of octadecadienoic (18:2) acid with conjugated double bonds (Bauman et al., 1999; Tsiplakou et al., 2006). Conjugated double bonds are in either cis or trans configuration and present in positions 7 and 9, 8 and 10, 9 and 11, 10 and 12 or 11 and 13 (counting from the carboxyl end of the molecule) (Pariza et al., 2001; Bhattacharya et al., 2006).
Plant lipids undergo two important transformations when exposed to the microbial population in the rumen namely hydrolysis and biohydrogenation. The hydrolysis causes the release of free fatty acids from esterified plant lipids whereas biohydrogenation reduces the number of double bonds (Bauman et al., 1999). The primary objective of these processes is to change unsaturated fatty acids such as octadecatrienoic (18:3) and octadecadienoic (18:2) acids into saturated end-products such as octadecanoic (18:0) acid. Conjugated linoleic acid isomers are incorporated into meat and milk of ruminants before the saturation process has been completed (Tanaka, 2005; Raes et al., 2004).
In vivo and in vitro studies have shown that CLAs have positive effects on cancer, cardiovascular diseases, diabetes and obesity (MacDonald, 2000; Bhattacharya et al., 2006). These potential health benefits of CLAs have intensified the research effort to enhance the levels of these fatty acids in meat and milk. Nutritional strategies for the enrichment of these products with CLAs are directly or indirectly related with unsaturated carbon 18 fatty acids (Bessa et al., 2000; AbuGhazaleh and Jacobson, 2007).
The pH of the rumen liquor depends on the balance between acidifying and neutralizing factors in the rumen (Allen, 1997). Some dietary components rich in readily fermentable carbohydrate such as grain reduce ruminal pH whereas other dietary components such as forage increase it (Kalscheur et al., 1997; Choi et al., 2005; Cajarville et al., 2006). Thus, the pH of rumen liquor physiologically may vary from 5.50 to 7 or slightly greater than 7 (Martin and Jenkins, 2002; Duffield et al., 2004; Wilkinson, 2004). The ruminal pH is an important factor in the rumen ecosystem which affects the bacterial fermentation (Allen, 1997; AbuGhazaleh and Jacobson, 2007). For example, a known bacterium, Butyrivibrio fibrisolvens which plays a key role for the formation of c9,t11 CLA isomer has maximum activity at pH values between 7.2 and 8.2 (Bessa et al., 2000). Decrease in rumen pH often results in bacterial population shift and consequent changes in the pattern of fermentation (Bauman et al., 1999).
Studying the relationship between CLAs and other carbon 18 fatty acids as well as rumen pH will provide an insight to devise strategies to increase CLA in ruminant food products. Even though rumen is a primary site for the formation of CLAs, limited studies have been conducted on their contents in the rumen liquor and their relation with other carbon 18 fatty acids and rumen pH. Therefore, the objective of the present study was to determine the level of CLAs in rumen liquor and to assess their relation with other carbon 18 fatty acids, as well as rumen pH in local Malaysian Kedah-Kelantan cattle slaughtered for meat consumption.
MATERIALS AND METHODS
Animals and sampling: Thirteen female and 9 male healthy Kedah-Kelantan cattle ((Bos indicus), a known indigenous breed in Malaysia) which were slaughtered in 2007 for meat consumption at abattoirs, were used for sampling. The animals’ age was approximately varied from 1.25 to 2.75 years (Mean = 2.37±0.45). They were owned and managed by different farmers under routine husbandry system. They were allowed for grazing and supplied with concentrates. The most common concentrate supplements were palm kernel cake, crop residues, fruit waste and other industrial by-products.
The rumen liquor was taken immediately upon opening and squeezed through a double-layered gauze to obtain 10 mL of the liquor. Its pH was measured immediately using a pH meter (MP120, Mettler-Toledo GmbH, Switzerland) and then 25% metaphosphoric acid (Sigma Chemical Co., St. Louis, Missouri, USA) was added at a ratio of one part acid to 4 parts of rumen liquor to reduce microbial activities. Samples were kept between blocks of ice, transported to the laboratory and stored at -20°C until analysis.
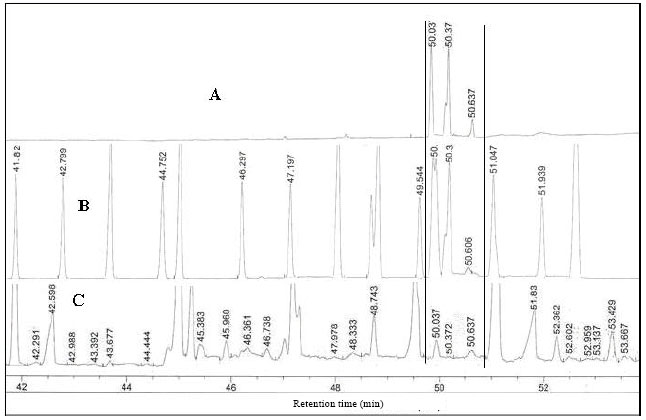 | |
| Fig. 1: | Representative chromatograph showing CLA peaks and other fatty acids in connection with their retention time. A: External standard containing CLA isomers only, B: External standard containing CLA isomers and other fatty acids, C: Peaks and retention time of a sample (rumen liquor) |
Fatty acid extraction and methylation: The total fatty acids were extracted using chloroform-methanol (2:1, v/v) (Merk kGaA, Darmstadt, Germany) based on modified Folch et al. (1957) method as it was described by Rajion et al. (1985). Methylation of the extracted fatty acids to Fatty Acid Methyl Esters (FAME) was carried out using 14% methanoic boron trifluoride (Sigma Chemical Co., St. Louis, Missouri, USA). Heneicosanoic acid (21:0) (Sigma Chemical Co., St. Louis, Missouri, USA) was used as an internal standard to facilitate the quantitative determination of fatty acids.
Chromatography: The FAME was separated on a HP-88 silica capillary column (60 m, 0.25 mm internal diameter, 0.20 μm film thickness) in a 6890 N, network gas chromatography system (Agilent Technology) which was fitted with flame ionization detector. High purity (99.99%) nitrogen (MOX, Sdn Bhd, Malaysia) was used as the carrier gas at 28.8 mL min-1 flow rate and the column head pressure was 22.52 psi. The injector and detector temperatures were programmed at 250°C. The column temperature was set at the range of 80-200°C with temperature programming at the rate of 2.5°C min-1 increment. One microliter of FAME was injected into the gas chromatography manually in splitless mode and 67 min was needed to run one sample. The individual FAME’s peak was identified using known standards of FAME containing CLA isomers (Sigma, Chemical Co., USA) and other 37 fatty acids (Sigma, Chemical Co., USA). Representative peaks of the standards and a sample are shown in Fig. 1.
Statistical analysis: Fatty acid profile was expressed in mean mg/100 mL of rumen liquor and percent total fatty acids with their respective Standard Deviation (SD). The relationships of CLAs with other fatty acids as well as rumen pH were analyzed using Pearson’s correlation. All statistical tests were conducted by statistical software (SPSS Version 15) and p<0.05 was considered significant.
RESULTS
Fatty acid profile: The fatty acid profile of rumen liquor from Kedah-Kelantan cattle slaughtered for meat consumption is shown in Table 1. Almost 50% of the total fatty acids were made by two saturated fatty acids (hexadecanoic (16:0) and octadecanoic (18:0)). The proportion of saturated fatty acids was 67.81±65.12% which showed rumen’s role as source of saturated fatty acid for ruminant’s tissue (Table 1).
| Table 1: | Fatty acid profile of rumen liquor from Kedah-Kelantan cattle slaughtered for meat consumption |
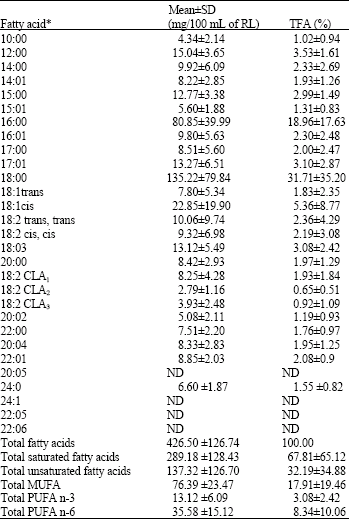 | |
| *Fatty acids are represented as the No. of carbon atoms (10-24) and No. of carbon-carbon double bond(s) (0-6), CLA1: Contains mainly c9, t11 CLA isomer, CLA2: Contains mainly t10, c12 CLA isomer, CLA3:Ccontains other isomers, Values were expressed in Mean±SD (standard deviation) and expressed in mg/100 mL of Rumen Liquor (RL) and percent total fatty acids (% TFA); ND: Not detected | |
Conjugated linoleic acids: The average amount of CLAs in the rumen liquor was 14.97±7.46 mg/100 mL and ranged from 5.52 to 30.55 mg/100 mL of the sample. Proportionally, CLAs were 3.50±0.76% of the total fatty acids. The percentages of c9, t11 and t10, c12 isomers of CLA were 55.11±16.87 and 18.64±15.61% of the total CLAs, respectively. The remaining proportion was taken by other minor isomers. Two key intermediate products, c9, t11 CLA and t11-octadecenoic acid were positively and significantly correlated (r = 0.538, p<0.05). Significant positive correlations were also observed between CLAs and octadecatrienoic (18:3) (r = 0.523, p<0.05), CLAs and cis-9 cis-12 octadecadienoic (18:2) (r = 0.551, p<0.05) and CLAs and octadecanoic (18:0) (r = 0.572, p<0.05).
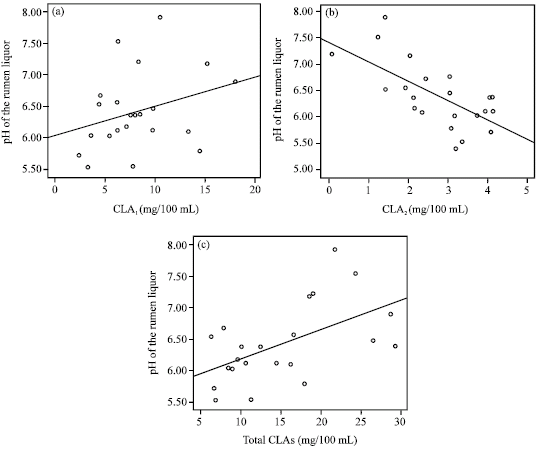 | |
| Fig. 2: | The correlations between rumen pH and c9, t11 CLA isomer (a, r = 0.305), t10,c12 CLA isomer (b, r = -0.661) and total CLAs (c, r = 0.547)(n = 22) as depicted by scattered plot (values) and the best fitting line |
Rumen pH and conjugated linoleic acids: The pH of the rumen liquor in the samples was ranged from 5.52 to 7.92 with the mean value of 6.41±0.64. It was correlated positively with c9, t11 (r = 0.305) and total CLAs (r = 0.547, p<0.05) (Fig. 2) but inversely with t10, c12 (r = -0.661, p<0.05) CLA isomer.
DISCUSSION
In the rumen, hydrolysis and biohydrogenation of glycolipids, phospholipids and triglycerides by microbes lead to the formation of free fatty acids that are more saturated (AbuGhazaleh et al., 2003). The CLAs present in the rumen liquor would be hydrolyzed to yield saturated fatty acid or passed to the next portion of the digestive system to be absorbed and incorporated in ruminant meat and milk products (Bauman et al., 1999). The ingesta reaching the small intestine contain predominantly saturated fatty acids and intermediate products including CLAs that escape the saturation process. Therefore, rumen microbial fermentation has a major impact on fatty acid profile of ruminants’ tissue (AbuGhazaleh et al., 2003; Wahle et al., 2004).
In this study, CLAs and t11-octadecenoic acid were positively correlated which was in agreement with the research work done by Bessa et al. (2000). Significantly positive correlations were also observed between total CLAs and other carbon 18 fatty acids which were consistent with the results reported by AbuGhazaleh et al. (2003). These indicate that the concentrations of CLAs and other 18 carbon fatty acids in the rumen liquor are related.
Ruminal pH is the known factor that affects biohydrogenation and hydrolysis processes in the rumen (Kalscheur et al., 1997; AbuGhazaleh et al., 2003). The amount of c9, t11 isomer of CLA in the present study was positively correlated whereas the quantity of t10, c12 isomer of CLA was negatively correlated with the rumen pH. In line with these findings, similar pattern of correlations were also reported by Choi et al. (2005). Decrease in rumen pH favours the formation of more t10 instead of t11 and thereby generates the t10, c12 isomer of CLA as an intermediate product instead of c9, t11 (Bauman et al., 1999). At higher pH, rumen bacteria produce higher proportion of c9, t11 whereas at lower pH, more proportion of t10, c12 isomer of CLA is produced (Choi et al., 2005). These results showed that t10, c12 CLA producing bacteria could be more acid tolerant than c9, t11 CLA producing bacteria. As the proportion of c9, t11 CLA isomer in milk and meat reach up to 90% of the total CLAs (Bauman et al., 1999; Khanal and Dhiman, 2004) and 55.11±16.87% in rumen liquor as it was obtained this study, the net effect of decreasing rumen pH will cause the total CLAs to decline in ruminant products.
In summary, the relationships and correlations observed between CLAs and other carbon 18 fatty acids as well as with rumen pH showed the possibility of increasing CLAs content in meat and milk of ruminants by feeding a diet rich in carbon 18 fatty acids especially octadecadienoic (18:2) and octadecatrienoic (18:3) acids and maintaining optimum pH in the rumen.
ACKNOWLEDGMENTS
The authors would like to thank the staff of Department of Veterinary Services, Putrajaya State Veterinary Department, Kuantan and Banting abattoirs, Malaysia and laboratory technicians for their contributions to this project which was funded by a research grant No. 05-01-04-SF0373, awarded by Ministry of Science, Technology and Innovation (MOSTI), Malaysia.
REFERENCES
- Bauman, D.E., L.H. Baumgard, B.A. Corl and J.M. Griinari, 2000. Biosynthesis of conjugated linoleic acid in ruminants. J. Anim. Sci., 77: 1-15.
Direct Link - Bessa, R.J., J. Santos-Silva, J.M. Ribeiroa and A.V. Portugal, 2000. Reticulo-rumen biohydrogenation and the enrichment of ruminant edible products with linoleic acid conjugated isomers. Livest. Prod. Sci., 63: 201-211.
CrossRefDirect Link - AbuGhazaleh, A.A. and B.N. Jacobson, 2007. The effect of pH and polyunsaturated 18 carbon fatty acid source on the production of vaccenic acid and conjugated linoleic acids in ruminal cultures incubated with docosahexaenoic acid. Anim. Feed Sci. Tech., 136: 11-22.
CrossRefDirect Link - Bhattacharya, A., J. Banu, M. Rahman, J. Causey and G. Fernandes, 2006. Biological effects of conjugated linoleic acids in health and disease. J. Nutr. Biochem., 17: 789-810.
CrossRefPubMedDirect Link - AbuGhazaleh, A.A., D.J. Schingoethe, A.R. Hippen and K.F. Kalscheur, 2003. Conjugated linoleic acid and vaccenic acid in rumen, plasma and milk of cows feed fish oil and fats differing in saturation of 18 carbon fatty acid. J. Dairy Sci., 86: 3648-3660.
CrossRefPubMedDirect Link - Cajarville, C., M. Aguerre and J.L. Repetto, 2006. Rumen pH, NH3-N concentration and forage degradation kinetics of cows grazing temperate pastures and supplemented with different sources of grain. Anim. Res., 55: 511-520.
CrossRefDirect Link - Choi, N., J.Y. Imm, S. Oh, B. Kim, H. Hwang and Y.J. Kim, 2005. Effect of pH and oxygen on conjugated linoleic acid (CLA) production by mixed rumen bacteria from cows fed high concentrate and high forage diets. Anim. Feed Sci. Tech., 123-124: 643-653.
CrossRefDirect Link - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - Khanal, R.C. and T.R. Dhiman, 2004. Biosynthesis of conjugated linoleic acid (CLA): A review. Pak. J. Nutr., 3: 72-81.
CrossRefDirect Link - MacDonald, H.B., 2000. Conjugated linoleic acid and disease prevention: A review of current knowledge. J. Am. Coll. Nutr., 19: 111S-118S.
PubMedDirect Link - Pariza, M.W., Y. Park and M.E. Cook, 2001. The biologically active isomers of conjugated linoleic acid. Prog. Lipid Res., 40: 283-298.
PubMedDirect Link - Raes, K., S. de Smet and D. Demeyer, 2004. Effect of dietary fatty acids on incorporation of long chain polyunsaturated fatty acids and conjugated linoleic acid in lamb, beef and pork meat: A review. Anim. Feed Sci. Technol., 113: 199-221.
CrossRefDirect Link - Tanaka, K., 2005. Occurrence of conjugated linoleic acid in ruminant products and its physiological functions. Anim. Sci. J., 76: 291-303.
CrossRefDirect Link - Tsiplakou, E., K.C. Mountzouris and G. Zervas, 2006. Concentration of conjugated linoleic acid in grazing sheep and goat milk fat. Livest. Sci., 103: 74-84.
CrossRefDirect Link - Wahle, K.W., S.D. Heys and D. Rotondo, 2004. Conjugated linoleic acids: Are they beneficial or detrimental to health?. Prog. Lipid Res., 43: 553-587.
CrossRefPubMedDirect Link - Allen, M.S., 1997. Relationship between fermentation acid production in the rumen and the requirement for physically effective fiber. J. Dairy Sci., 80: 1447-1462.
CrossRefDirect Link - Duffield, T., J.C. Plaizier, A. Fairfield, R. Bagg and G. Vessie et al., 2004. Comparison of techniques for measurement of rumen pH in lactating dairy cows. J. Dairy Sci., 87: 59-66.
Direct Link - Kalscheur, K.F., B.B. Teter, L.S. Piperova and R.A. Erdman, 1997. Effect of dietary forage concentration and buffer addition on duodenal flow of trans-C18:1 fatty acids and milk fat production in dairy cows. J. Dairy Sci., 80: 2104-2114.
Direct Link - Martin, S.A. and T.C. Jenkins, 2002. Factors affecting conjugated linoleic acid and trans-C18:1 fatty acid production by mixed ruminal bacteria. J. Anim. Sci., 80: 3347-3352.
Direct Link