
Govinda Rizal
C4 Rice Center, International Rice Research Institute, DAPO 7777, Metro Manila, Philippines
Shanta Karki
C4 Rice Center, International Rice Research Institute, DAPO 7777, Metro Manila, Philippines
Vivek Thakur
C4 Rice Center, International Rice Research Institute, DAPO 7777, Metro Manila, Philippines
Jolly Chatterjee
C4 Rice Center, International Rice Research Institute, DAPO 7777, Metro Manila, Philippines
Robert A. Coe
C4 Rice Center, International Rice Research Institute, DAPO 7777, Metro Manila, Philippines
Samart Wanchana
C4 Rice Center, International Rice Research Institute, DAPO 7777, Metro Manila, Philippines
William Paul Quick
C4 Rice Center, International Rice Research Institute, DAPO 7777, Metro Manila, Philippines
Asian Journal of Cell Biology
Year: 2012 | Volume: 7 | Issue: 2 | Page No.: 13-31







ABSTRACT
The success of the green revolution that began in the 1960s led to an increase in rice yield of up to 10 fold that was sufficient to meet the food demand of a continually growing population. This rapid increase in rice yield has not been repeated even after four decades, while the global population continues to grow rapidly. To meet the food demand of the soaring population, rice yields should increase by at least 50% within next four decades. Despite the best efforts of plant breeders, the prospect of a second drastic increase in yield using conventional approaches is unlikely but may be possible if we redesign rice photosynthesis into C4 type. A C4 type of photosynthesis is much more efficient than the C3 type found in wheat and rice and could facilitate an increase in yield potential to levels found in C4 crops like sorghum and maize. Although, C4 plants evolved from the C3, there are distinct differences in their leaf structure (evolution of Kranz anatomy) and biochemistry. The same would be needed for the development of a C4 rice plant. In 2009, a consortium of scientists was formed to develop C4 rice. The consortium is using multiple approaches which include but are not limited to; alteration of genomes of C3 and C4 plant species, exploration of wild Oryza accessions and bioinformatics to discover new genes underlying C4 photosynthesis. The already known or newly discovered genes of C4 pathway are being systematically introduced into rice. Here we review the strategies adopted by the C4 rice project coordinated by the International Rice Research Institute.
PDF Abstract XML References Citation
Received: June 15, 2012;
Accepted: August 10, 2012;
Published: October 20, 2012
How to cite this article
Govinda Rizal, Shanta Karki, Vivek Thakur, Jolly Chatterjee, Robert A. Coe, Samart Wanchana and William Paul Quick, 2012. Towards a C4 Rice. Asian Journal of Cell Biology, 7: 13-31.
DOI: 10.3923/ajcb.2012.13.31
URL: https://scialert.net/abstract/?doi=ajcb.2012.13.31
DOI: 10.3923/ajcb.2012.13.31
URL: https://scialert.net/abstract/?doi=ajcb.2012.13.31
INTRODUCTION
Rice has been cultivated for more than 9,000 years (Molina et al., 2011) and it is the primary source of energy for more than half of the world’s population. The well-known green revolution in the 1960s led to an increase in the rice yield from less than 1.5 ton ha-1 (Jennings, 1964) to the present rice yield potential of 8-10 ton ha-1 (Khush, 1995). This increase in food production sustained the simultaneous increase in global population from 3 billion in 1961 to 7 billion within five decades. The population is expected to reach 9 billion in the next four decades. The major portion of this population increase will be in the rice eating areas of Asia and Africa. To harvest enough rice for the booming population, there is a need for new varieties which can sustainably yield higher, under the perceived situation of less land, water and fertilizer inputs amid the predicted extremes of climatic change. Even with the optimum use of input and management practices, the yield potential of the current inbred indica varieties does not exceed 10 ton ha-1 (Kropff et al., 1993). One way to overcome the current yield ceiling is by genetically introducing the C4 photosynthetic pathway in rice (Sheehy et al., 2000). The concepts and practicalities of redesigning rice photosynthesis are reviewed in three classic books (Raghavendra and Sage, 2011; Sheehy, 2008; Sheehy et al., 2000). A number of reviews have underscored the importance of engineering C4 pathway in rice (Gowik and Westhoff, 2011; Hibberd and Covshoff, 2010; Kajala et al., 2011b; Langdale, 2011; Matsuoka et al., 2001; Sage and Zhu, 2011). All of these literature highlight that C4 photosynthesis is much advantageous than C3 in rice growing areas under high temperature and bright sunshine. This led to the idea to upgrade C3 rice to C4. No doubt this is a complex process as multiple genetic changes are required to alter leaf biochemical and anatomical modifications that occurred during the evolution of C4 plants from their C3 ancestors. C4 photosynthesis has evolved multiple times independently during the evolution of plants (Sage, 2004) and has involved duplication of genomes in whole or part that has created redundancy in genes, evolution of Kranz anatomy, a decrease in number of Mesophyll (M) cells between veins, a spatial shift in the metabolism of M and Bundle Sheath (BS) cells including the relocation of carboxylase enzymes (Gowik and Westhoff, 2011) and evolution of transporters to facilitate metabolic movement between the two cell types (Fig. 1b). The absence of any of these C4 traits in close relatives of rice prevents its incorporation through classical breeding strategies. The advancements in genetic engineering and plant molecular biology offer great possibilities to incorporate C4 photosynthesis in rice. While the process of photosynthesis is widely studied in both C3 and C4 plants and the introduction of C4-like metabolism into M cells of C3 plants has been attempted (Miyao et al., 2011), transforming one photosynthesis type into another is a novel attempt. This review reported the strategies being adopted and progress made towards novel C4 gene identification and transformation of known genes to integrate C4 photosynthetic system into C3 rice.
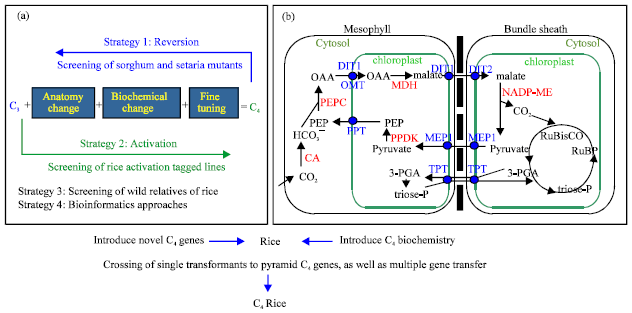 | |
| Fig. 1(a-b): | Schematic diagram of (a) novel genes related to C4 pathway, identified by utilizing various resources and (b) the known biochemical components of Nicotinamide adenine dinucleotide phosphate-malic enzyme (NADP-ME) type C4 pathway, currently being engineered into rice. The core C4 genes are in red and transporters are in blue color. PGA: Phosphoglycerate, OAA: Oxaloacetic acid, PEP: Phosphoenol pyruvate, Full names of other abbreviations used in the figure are given in Table 1 |
COMPARISON OF C3 AND C4 PHOTOSYNTHETIC PATHWAYS
The C4 photosynthetic pathway, an evolutionary development from C3, has undergone modifications in terms of biochemistry as well as leaf anatomy (Gowik and Westhoff, 2011). Most plants are either C3 or C4 depending on the type of photosynthetic pathway they use which are classified based on the first compound formed from the assimilation of atmospheric CO2. In C3 plants, CO2 is assimilated by Ribulose-1,5-bisphosphate Carboxylase Oxygenase (RuBisCO) and forms 3-phosphoglycerate (3PGA), a 3 carbon compound. The entire process is completed within M cells which are exposed to atmospheric CO2 via intercellular air spaces and have an abundance of RuBisCO enzyme. RuBisCO fixes both CO2 and O2 depending on their availability thereby facilitating both carboxylation and oxygenation reactions. The amount of RuBisCO in C3 plants is much more than the plant actually uses was evident from an experiment where tobacco plants survived normally when RuBisCO was reduced by up to 50% (Quick et al., 1991). The oxygenase activity of RuBisCO increases photorespiration and causes energy loss (Edwards et al., 2004) although this can be beneficial in times of stress as a means to dissipate excess energy (Yokota and Shigeoka, 2008). In contrast, RuBisCO in C4 plants is not expressed in M cells rather its expression is restricted to BS cells. The atmospheric CO2 is first fixed in M cells by phosphoenolpyruvate carboxylase (PEPC) forming oxaloacetate, an organic compound with 4 carbon units (Fig. 1b). Unlike RuBisCO, PEPC is an oxygen-insensitive carboxylase which has a much higher affinity for CO2 and can continue carboxylation even when the leaf internal CO2 concentration is very low (Ku et al., 1996). The CO2 fixed in the form of 4 carbon acid is transported to BS cells where it is decarboxylated by one of the three decarboxylating enzymes. The CO2 released after decarboxylation is re-fixed in BS cells by RuBisCO (Fig. 1). This decarboxylation of C4 acids creates a very high concentration of CO2 around RuBisCO. This mechanism to concentrate CO2 in BS cells prevents photo-respiratory oxygenation reactions making C4 plants generally more productive than C3 (Peterhansel and Maurino, 2011). Photorespiration is greatly enhanced at higher temperatures due to a lower CO2/O2 specificity of the RuBisCO enzyme and the enhanced solubility of oxygen in water compared to CO2. Therefore, especially in warmer climates, C4 plants have elevated photosynthesis compared to C3 plants in the same ecology (Long, 1999).
The ability of C3 (rice) and C4 (sorghum) plants to assimilate photosynthates was studied at different light intensities measured as Photosynthetic Photon Flux Density (PPFD) using infrared gas analyzer. The PPFD was gradually decreased from 2500 μmolphoton m-2 sec-1 to zero, CO2 was kept constant at 400 ppm, a flow rate of 400 μmol sec-1 was maintained and the leaf temperature was 30°C. It showed that the C4 plants can also utilize maximum level of sunlight because their rate of photosynthesis sharply increases with increasing light intensity and does not appear to saturate (Fig. 2). Radiation Use Efficiency (RUE) is up to 50% higher than that of C3 due to reduced number of photons required to fix each molecule of CO2 (Sheehy, 2008). A typical C4 plant produces 1 g of biomass for every 250-350 g of water transpired, whereas to produce the same quantity of biomass C3 plants transpire 650-800 g of water (Ehleringer and Monson, 1993). Plants with C4 photosynthesis are more efficient in using photosynthetic nitrogen than C3 plants largely due to reduced amounts of RuBisCO protein required to achieve the same rate of photosynthesis (Sage et al., 1987). The C4 CO2 concentrating mechanism therefore confers more efficient photosynthesis, combined with greater nitrogen, water and radiation use efficiency and this frequently translates into higher yields in crop plants.
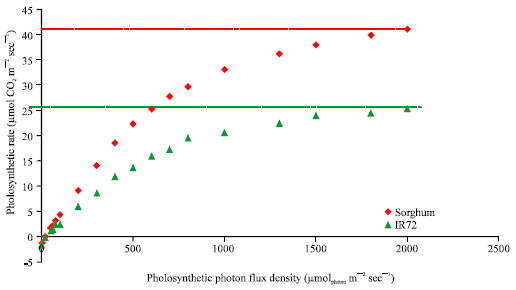 | |
| Fig. 2: | A comparison between light response curves of a C4 plant (sorghum) and C3 plant (rice, IR72) generated using readings obtained from infrared gas analyser |
SEARCH FOR GENETIC FACTORS CONTROLLING C4 LEAF ANATOMY
While the metabolic process of photosynthesis has been extensively researched and photosynthetic enzymes well studied, most of the genetic factors regulating C4 anatomy are still unknown (Langdale, 2011). We have adopted a dual approach to identify these genetic factors. The first approach is to mutate C4 plants (sorghum and Setaria viridis) to randomly hit some of the C4 characteristics and to identify the responsible genetic factors (Fig. 1a). The second approach is to use rice DNA activation tagging to over-express random rice genes and look for C4-like characteristics (Fig. 1b). Both of these approaches require screening of large populations to find desirable phenotypes. For mass screening of these populations, we searched for induced alterations in leaf Vein Density (VD). The veins of C4 plants are generally much more closely spaced than C3 plants. This is required to facilitate the precise spacing of M and BS cells (vein-BS-M-M-BS-vein) and the general requirement for a 1:1 stoichiometry of BS and M cells. Alterations to vein spacing can arise through a number of anatomical changes including alterations to M cell number between the veins, altered BS size and number or increased vein size. All of these are important changes that have occurred in C4 plants and so identification of genes that disrupt or induce these changes in C4 (sorghum) or C3 (rice) respectively could help us to understand how leaf anatomy is regulated at the genetic level. Fortunately, analysis of vein density is a relatively simple procedure that can be undertaken very quickly with a hand-held microscope in the field or laboratory and is suited to screening large populations.
INDUCED MUTATION ALTERS C4 LEAF ANATOMY
Induced mutations have greatly facilitated gene discovery, the understanding of complex traits and enhanced the speed of novel gene identification. We treated sorghum (Sorghum bicolor (L.) Moench) seeds with Ethyl Methanesulfonate (EMS) and gamma rays separately to generate two mutant populations. Based on the alteration in vein spacing from wild type VD (more than 8 veins mm-1 at the widest part of the youngest fully expanded leaves) to low VD (LVD) (less than 7 veins mm-1), we screened more than 70,000 mutant lines in the M2 generation. It was found that mutagenesis had caused a reduction in VD, largely due to increased M cell number between the veins, in 24 independent lines that we are currently analyzing in more detail. To reduce the number of non-specific mutations we are currently generating backcrossed (BC2F2) populations. To identify mutations a number of techniques are available, such as genotyping single nucleotide polymorphisms (Kwok, 2001), denaturing HPLC (DHPLC) or TILLING (McCallum et al., 2000), endonuclease cleavage method (Oleykowski et al., 1998), cel 1 based TILLING method (Till et al., 2004) and high-resolution melt analysis (Ririe et al., 1997). With the improved speed and reduced cost of large-scale DNA sequencing we have opted for next generation genome sequencing to identify the responsible mutated genes as the reference genome sequence for sorghum-BTx623 is publicly available (Paterson et al., 2009). We are currently analyzing the sequence data and hope to identify the relevant gene or genes that will then be transformed into rice and the mutant sorghum lines to analyze the effects of their expression. This direct approach of mutation discovery quickens the process of gene identification.
EMERGENCE OF C4 CHARACTERISTICS IN MUTANT RICE LINES
The most direct method in functional gene discovery is to look for a correlation between phenotype and genotype within a specific mutant. Elimination or activation of gene function through insertional mutagenesis using transfer DNA (T-DNA) or transposable elements has proven to be an extremely valuable research tool. The foreign DNA acts as a mutagen and a tag for the site of insertion allowing for a function to be assigned to a specific DNA sequence and for genes associated with a specific phenotype to be isolated. The insertion of these elements is a random event and the mutations are stable through multiple generations (Azpiroz-Leehan and Feldmann, 1997). Such approaches have their limitations, particularly when they are compensated by gene redundancy, lethality due to loss of function or where specific unknown conditions are required for the gene activity such as biotic/abiotic stresses. To overcome these limitations modified insertional elements have been developed like the gene trap systems that fuse the tagged gene with a reporter gene, such as β-glucuronidase (GUS) or Green Fluorescent Protein (GFP) (Springer, 2000). Activation tagging technique uses T-DNA or a transposable element containing multimerized Cauliflower Mosaic Virus (CaMV) 35S enhancers. They enhance gene functions in both orientations at considerable distance from their site of insertion. This results in the transcriptional activation of genes and dominant gain-of-function mutations. The resulting mutant phenotypes can reveal the normal function of the gene while the tag facilitates the identification of the location in the genome.
A considerable number of rice mutant resources have been created using chemical mutagenesis (Wu et al., 2005; Till et al., 2007), T-DNA insertion (An et al., 2003; Chen et al., 2003; Chern et al., 2007; Hsing et al., 2007; Jeong et al., 2002, 2006; Mori et al., 2007; Wan et al., 2009; Wu et al., 2003), the retrotransposons Tos17 (Hirochika, 2001), maize transposon Activator/Dissociation (Ac/Ds) elements (Kolesnik et al., 2004; Park et al., 2007; Qu et al., 2008; Upadhyaya et al., 2006), maize enhancer/suppressor mutator elements (En/Spm) (Kumar et al., 2008) and the Full-length cDNA Over-expressor gene (FOX) hunting system (Hakata et al., 2010; Kondou et al., 2009; Nakamura et al., 2007). The first rice genes identified using insertional mutagenesis were discovered simultaneously using forward (Agrawal et al., 2001) and reverse (Takano et al., 2001) genetic screens of separate Tos17 mutant populations. Genes have also subsequently been identified in forward (Jung et al., 2003), reverse (Lee et al., 2003), expression based (Kang et al., 2005) screens of T-DNA populations and populations utilizing the Ac-Ds transposon (Zhu et al., 2003, 2004) and the FOX-hunting systems (Nakamura et al., 2007) highlighting the value of these resources for functional gene discovery. Databases are available for each of these individual populations as well as databases that combined data from multiple insertional mutagenesis projects such as RiceGE/SIGnAL (http://signal.salk.edu/cgi-bin/RiceGE), OryGenes DB (http://orygenesdb.cirad.fr) and Gramene (http://www.gramene.org). Together these resources represent an extensive and diverse resource in various genetic backgrounds (Droc et al., 2006; Ware et al., 2002). The availability of mutant resources, advanced analytical techniques and the large number of databases aid to the identification of the genes of interest.
We are screening activation insertion mutants from Korea (http://postech.ac.kr/life/pfg/risd/) and Taiwan (http://trim.sinica.edu.tw). While the Korean collection consists of 47,932 T-DNA activation lines in the japonica varieties Dongjin and Hwayoung (Hsing et al., 2007), the Taiwan collection contains approximately 70,000 T-DNA insertions in the japonica variety, Tainung-67 (Jeong et al., 2006). Approximately 80,000 Flanking Sequence Tags (FSTs) are available for the Korean population and 25,000 for the Taiwan collection. The aim is to identify mutants with alteration in leaf anatomy that resemble C4 traits in rice by activation of specific genes. This could lead to the identification of novel genes controlling C4 leaf anatomy, cell biology and ultra structure. To date approximately 17,000 lines have been screened for mutants with a stable increase in VD. From the two populations more than 60 mutants have been identified with increased VD. In some of these lines, the increase is due to a reduction in the number of M cells between veins, a feature characteristic of C4 leaf anatomy. Such mutants can be used to identify the genes controlling the phenotype either through sequencing of mutagenised lines and/or transgenic testing of candidate genes. In future we will also screen Ac/Ds mutant lines from Australia (Upadhyaya et al., 2006) and FOX lines from Japan (Nakamura et al., 2007).
HARNESSING THE DIVERSITY OF RICE
Rice is one of the cultivated species of genus Oryza of Poaceae family. Genus Oryza is composed of 10 different genomes with 23 wild and 2 cultivated species. Although C4 evolved independently more than 60 times (Monson, 2003) and there are symptoms of C4 characteristics in the stem and petiole of C3 plants like tobacco (Hibberd and Quick, 2002), there are no occurrences of C4 or C4-like species within this genus Oryza. However, Oryza possesses a wide range of variation for different traits even among the accessions of a single species. Ram et al. (2007) have reported 56% genetic diversity among its diverse genomes. Among these species, Oryza rufipogon and O. nivara are currently being screened for C4-traits as these two species have higher intra-species genetic diversity; 46% for O. rufipogon (Gao et al., 2002) and 78% for O. nivara (Juneja et al., 2006) and differences in the branching pattern (Yamaki et al., 2010) and M cell conductance (Scafaro et al., 2011) were found to be correlated with yield and availability of CO2 for carboxylation, respectively. The advantage of working with wild rice species is the readily available information about genome sequences and genetic resources in Gramene database (http://www.gramene.org), Oryzabase (http://www.shigen.nig.ac.jp/rice/oryzabase/top/top.jsp), Rice-BRCdb (http://rice-brcdb.mpl.ird.fr/brcdb/) and the Oryza Map Alignment Project (OMAP, http://www.omap.org/). A high resolution Universal Core Genetic Map (UCGM) was developed by Orjuela et al. (2010) using 165 anchors and 16 accessions from O. sativa, O. barthii, O. rufipogon, O. glumaepatula and O. meridionalis. This group also developed the Paddy Map for easily generating sets of polymorphic SSR markers in any interspecific or intraspecific cross of Oryza species. Several Chromosomal Segment Substitution Lines (CSSLs) have been developed for rice by small wild rice chromosomal introgressions which were found to be widely useful for identifying QTL and genes for different traits. These resources are still underutilized for physiology and photosynthesis screening. Therefore, there exists huge potential to modulate rice physiology using these resources to improve C3 photosynthesis of rice or to make it more suitable for C4 photosynthesis. Many agronomically important traits have already been successfully introgressed into rice by conventional breeding programs (Amante-Bordeos et al., 1992; Brar and Khush, 1997, 2002; Jena, 2010). A wide variation in leaf morphology in wild Oryza species provides good reason to explore these materials in more detail that would help in rice leaf anatomy and physiology manipulation. Several studies are ongoing world-wide to look for C4-like properties in wild rice. The pioneering work of Yeo et al. (1994) showed increasing PEPcase activity in O. australiensis and much lower photorespiration rate in O. rufipogon, both of which are of direct interest in the study of C3 to C4 interconversion. They also observed large differences among Oryza species in transpiration at the single leaf level. Recent studies have shown that there is a wide variation in the photosynthetic characteristics among the wild rice (Zhao et al., 2010). They assessed photosynthesis related traits like net photosynthetic rate, leaf chlorophyll content, specific leaf area, flag leaf area and leaf N2 concentration and reported higher photosynthetic rates in O. rufipogon and O. australiensis. Both studies suggest O. rufipogon as one of the suitable candidates for improving photosynthesis in rice. At International Rice Research Institute (IRRI), the first attempt of mass screening of wild rice accessions for VD change and lower CO2 compensation point was carried out in 2008. After mass screening of CO2 compensation point, we found a reduction in CO2 compensation point for a few O. nivara accessions. These results indicate that genes responsible for leaf anatomical changes cause changes in observed CO2 compensation point and could be identified from wild rice relatives and can be used to enhance our C4 pathway engineering efforts into cultivated rice.
GENOMIC APPROACHES FOR UNDERSTANDING REGULATION OF C4 PHOTOSYNTHESIS
In a recent study, four key genes of the C4 pathway were introduced into rice M cells to generate a C4 cycle between the chloroplast and cytoplasm. Although this did not result in enhanced photosynthesis or reduced compensation point it did demonstrate the possibility to achieve C4 type photosynthesis in C3 plants (Taniguchi et al., 2008). With a two cell system, we need to observe the desired cell specific expression patterns, suitable level of expression and activity and the metabolic connectivity of the two cell types. So, it becomes important to also have knowledge of the regulatory network controlling C4 biochemistry and anatomy. This requires discovery of (1) additional genes working in coordination with the known C4 genes, (2) transcription factors regulating the activities of genes of network and their respective binding sites and (3) involvement of gene silencing mechanisms. Several functional genomics and bioinformatics approaches have been or are being applied to meet these objectives.
For the discovery of genes associated with any phenotypic trait, gene expression profiling has been one of the most commonly used approaches. Although it was known for a long time that the mature leaves relay signals of environmental cues to new leaves of the same plant (Lake et al., 2001), the differential expression within the same leaf are being studied recently (Li et al., 2010). Our experimental designs involve comparison of gene expression between (1) C3 and C4 leaves, (2) M and BS cells (3) developmentally distinct regions of a leaf and (4) different growth stages within a leaf. These experimental designs are meant to capture different and/or overlapping biological processes underlying C4 photosynthesis. A few gene expression data has been generated by microarray analysis (Sawers et al., 2007), however, majority has been generated by next generation sequencing of the transcriptome (RNA-seq), as discussed in detail below.
TRANSCRIPTOMES OF C3 AND C4 LEAVES
Comparison of gene expression in leaves of C3 and C4 species could identify genes that have turned on or off during the evolution of multiple C4 species independently, or genes whose overall expression has significantly changed. There are two such studies between C3 and C4 species of genera Cleome and Flaveria (Brautigam et al., 2011; Gowik et al., 2011). Choice of such species minimizes the differences in expression due to species-specific features, thus allowing more precise discovery of genes associated with differences in photosynthetic pathway. The findings from both studies unveiled that in addition to the core C4 pathway, there are several other functional gene classes which are affected (Brautigam et al., 2011). Among the classes which showed lower steady state mRNA level included Calvin-Benson cycle, photorespiration, protein synthesis, primary metabolism, while those which showed higher level included photosynthetic classes of photosystem1 (PSI) and cyclic electron flow, starch metabolism, nitrogen metabolism, cofactor synthesis, glucan metabolism and lipid transfer proteins. There were some deviations too: Flaveria additionally showed downregulation of PSII, attributed to different ATP and NADPH demand related to the different mode of photosynthesis in the two species. Apart from the affected gene classes, some genes were also reported as candidate C4 genes. The discovery of transporters was most important among them, as they ensure the availability of metabolites to the enzymes present in different cellular compartments. Few plastidic and mitochondrial transport proteins were largely up-regulated in C4 leaf tissue (Brautigam et al., 2011). Among the proteins with regulatory functions, 43 were significantly up-regulated in either C3 or C4 Cleome, whereas in Flaveria, several hundreds of such proteins were found to be differentially expressed and some of the important ones include auxin response factor2 (ARF2), golden2 like (GLK2), plastidic Sigma-70 like factors (SIG1 and SIG5) (Gowik et al., 2011). Other genes related to chloroplast positioning, such as giant chloroplast1 (GC1) and chloroplast unusual positioning1 (CHUP1) were also reported to be candidates for C4-associated genes.
TRANSCRIPTOMES OF MESOPHYLL AND BUNDLE SHEATH CELLS
The comparison between C3 and C4 whole leaf tissue is likely to miss genes expressed in different locations (or specific cell types), unlike their expression levels. Consequently to identify such genes, a more appropriate experimental design would be needed. From the perspective of the C4 pathway, isolation of RNA from M and BS cells separately are obvious choices. The findings in the case of maize by using either microarray or RNA-seq showed that about one-fifth of genes, expressed in leaves, are differentially expressed between M and BS cells (Sawers et al., 2007; Li et al., 2010). Several functional classes were partitioned between the two cell types including light harvesting complexes, respiration, secondary metabolism and transport. Comparison of the transcriptome data with proteome datasets of maize plastids (Friso et al., 2010) showed near-perfect correspondence between cell-specific enrichment and high correlation between differential expression (0.68<r<0.98) (Li et al., 2010). This indicates that genes involved in C4 photosynthesis are largely regulated at the transcriptional level.
TRANSCRIPTOMES ACROSS LEAF DEVELOPMENTAL GRADIENT
In addition to identifying genes involved in functioning of the photosynthetic pathway, it is also important to understand the factors involved in leaf development. This will help in identifying factors involved early in leaf development for building the cellular infrastructure where photosynthesis could operate. One such study was conducted by Li et al. (2010) in maize leaves examining the expression profile of genes in four developmentally distinct segments, namely basal, transitional, maturing and matured zones. While the fraction of functional genes was slightly higher in segment undergoing development (leaf base) compared to the fully mature segment, alternate splicing events were observed in about half of the expressed genes with introns. About eighteen distinct expression profiles were identified; six of them were considered main clusters accounting for 82% of the total genes expressed. Two clusters represented genes that are expressed at the highest levels towards the basal zone and include genes that encode enzymes for cell wall biosynthesis, DNA synthesis, cell cycle regulation and chromatin structure, protein metabolism, potential signaling proteins, auxin and brassinosteroid biosynthesis and signaling, respiratory pathways and vesicle transport. A marked change in the nature of enriched functional categories was observed for cluster showing peak expression of genes in the transition zone. Similarly, in the maturing or matured segments, the enriched gene classes included isoprenoid biosynthesis, the Calvin cycle, redox regulation and the light reactions of photosynthesis (Li et al., 2010).
REGULATORY ELEMENTS ASSOCIATED WITH C4 GENES
Known regulatory elements are often of poor resolution: The cell-specific expression in M and BS cells is generated by both transcriptional and post-transcriptional mechanisms and only for some of the C4 enzymes is the mode of regulation known, at least partially (Hibberd and Covshoff, 2010). For the regulation at the transcriptional level, cis-regulatory elements form a key component and these often reside in the range of 0.5-2 kb upstream of the transcriptional start site. Here we summarize the available information about transcriptional regulation of C4 related genes that involve cis-elements. In the case of PEPC, in C4 Flaveria, a 41 bp long region named M-enhancing module (MEM1) has been identified which drives expression in M cells (Gowik et al., 2004; Akyildiz et al., 2007). This module is located in the distal region of the core promoter and is constituted of a tetranucleotide (CACT) insertion and a nucleotide substitution (G→A). In maize, however, the M cell specificity is attributed both to a 0.6 kb upstream region (Taniguchi et al., 2000; Kausch et al., 2001) and some epigenetic modifications (Langdale et al., 1991). For PPDK, while in maize a very specific region (-301 to -296 from translational start site) is reported to be important (Matsuoka and Numazawa, 1991), in C4 Flaveria a 1 kb long region (-1212 to +279 from transcriptional start site) accounts for higher M cell expression relative to BS cells (Rosche et al., 1998). For NADP-ME, the behavior of two C4 Flaveria species were different and the one showing transcriptional regulation, namely F. trinervia, required a 2 kb region (-1758 to +305 from translational start site) for strong BS-specific expression (Lai et al., 2002). Like NADP-ME, the regulation of small subunit of RuBisCO was found to be complex and different regions for M cell repression and BS expression were reported (Viret et al., 1994). Other C4 genes are currently being studied within the C4 Rice program to derive more cell specific promoters with appropriate expression levels.
PREEXISTING ELEMENTS RECRUITED FOR CELL SPECIFIC EXPRESSION OF C4 GENES
Recent studies on the origin of cis-elements recruited by C4 genes have unveiled some interesting findings. Based on the experiments on NAD-ME and NADP-ME, it was reported that genes from C3 species contain cis-elements sufficient for BS specificity in C4 leaves (Brown et al., 2011). The cis-elements were found to be located in the coding region and required to be transcribed to be functional which strongly suggest a post-transcriptional nature of regulation (Brown et al., 2011). When the hypothesis was tested in two more C4 genes, namely PPDK and CA, one or both Untranslated Regions (UTRs) were found to be sufficient for enhanced expression in M cells (Kajala et al., 2011a).
DISCOVERY OF CIS-ELEMENTS
The locations of cis-elements of several C4 genes are available but at a poor resolution. For a successful transplantation of C4 pathway in rice we would require high resolution information about these cis-elements. The experimental approach of promoter-deletion assay provides reliable information, however, is of poor resolution in terms of location and requires significant research efforts. In silico approaches greatly help in accelerating their discovery using the prior information with improved accuracy. The recent findings on the presence of cis-elements in the transcript region (Brown et al., 2011; Kajala et al., 2011a) will further prevent the typical searches from being restricted only to the upstream regions. Availability of relevant gene expression data between photosynthetic cell types, C3/C4 leaf tissues and across the developmental gradients has helped in identification of co-expression clusters which in turn could be used for cis-element prediction. Moreover, the availability of draft genome sequences of various grass species like maize, sorghum and Setaria, also allows for sequence conservation studies of the upstream sequences housing the regulatory elements.
GENE SILENCING: EPIGENETICS AND NON-CODING RNAS
C4 differentiation occurs along a developmental gradient with undeveloped proplastids found in cells at the leaf base and fully differentiated C4 M and BS chloroplasts at the leaf tip (Li et al., 2010). C4-specific genes were recruited from existing C3 genes to encode the components necessary to the C4 mechanism. A true C4 cycle requires the co-ordinated activity of multiple enzymes in different cell types and in response to diverse environmental and metabolic stimuli. The selective expression of specific C4 pathway genes in specific cell types requires multiple levels of regulation. Although cis-acting DNA elements are important for gene regulation, chromatin configuration also plays a vital role. The expression of PEPC has been linked to epigenetic control, histone modification and chromatin remodeling in maize (Danker et al., 2008). These histone modifications significantly contribute to gene regulation by acetylation of specific histone lysine residues. Interestingly, this pattern is not dependent on gene activity, but is already established in etiolated plants. Cell-type specific chromatin modifications potentiate subsequent light activation of transcription during differentiation of photosynthetic tissues in C4 plants.
There is another gene regulatory mechanism which is likely to be involved in functioning of C4 specific features: the down-regulation of gene expression through non-coding RNAs (ncRNAs). Among various ncRNA types, the micro RNAs (miRNAs) are relatively well characterized and often show conservation across plant and animal species (Jones-Rhoades et al., 2006). Since majority of miRNA families have been reported to be involved in plant development including the vascular development (Jones-Rhoades et al., 2006; Rubio-Somoza and Weigel, 2011), their role in development of Kranz anatomy and down-regulation of photosynthesis related genes cannot be ruled out. To investigate this aspect, small RNA sequencing of appropriate leaf samples is being pursued within the C4 consortium. The sequencing data will further be analyzed to discover novel and existing miRNAs, for which a protocol is in place for high accuracy prediction of plant miRNAs from deep-sequencing data (Thakur et al., 2011).
MOLECULAR ENGINEERING APPROACHES TOWARDS DEVELOPMENT OF C4 RICE
Introducing C4 photosynthesis genes into rice: In the past there have been attempts to transfer C4 traits into C3 plants by conventional plant hybridization between the two species. This approach was useful for a limited number of plant genera such as Atriplex, Brassica, Panicum, Moricandia and Flaveria (Brown and Bouton, 1993). Unfortunately, most of those C3-C4 hybrids showed infertility due to abnormal chromosome pairing and other genetic barriers. Because many of the major traits associated with C4 photosynthesis are absent from all rice species assessed to date and wide hybridization between sorghum and rice, oat and maize failed to transfer the C4 cycle as a whole, the use of conventional breeding to achieve this goal seems unlikely. Therefore, a genetic engineering approach seems to be the most appropriate technology to transfer C4 traits into C3 plants. Although isoforms of genes encoding C4 enzymes are also present in C3 plants, they are usually expressed at very low levels and in the wrong cell types. Recent developments in plant molecular biology and genetic engineering have made it possible to introduce the desired genes encoding C4 enzymes into C3 plants using transgenic techniques (Matsuoka et al., 2001; Miyao et al., 2011). These efforts have deepened our understanding of the mechanism of C4 photosynthesis and provided valuable information about the functions and evolution of these C4 genes. This has enabled scientists to express enzymes involved in the C4 pathway at high levels comparable to C4 species and in desired locations even in the leaves of C3 plants. C4 photosynthesis depends on synchronized division of labor between M and BS cells which is achieved by differential expression of the genes encoding the enzymes and transporters of the C4 pathway. Based on primary C4 acid decarboxylating enzymes used, the C4 pathway is divided into 3 subtypes: NAD-malic enzyme, NADP-malic enzyme and PEP carboxykinase types (Huber and Edwards, 1975). In a typical NADP-ME C4 type plant, e.g maize, 21% of genes are differentially expressed between BS and M cells (Li et al., 2010). Promoters with BS or M specific activity from the C4 grasses can be used to drive tissue specific transgene expression in rice leaves. For example, the promoter of PEPCK gene from Zoysia japonica fused with β-glucuronidase expressed selectively in vascular tissues and BS cells of transgenic rice (Nomura et al., 2005). This result demonstrates that some of the C4 specific genes localized in BS cells can retain their property of cell specificity even in a C3 plant suggesting that C3 plants still possess a regulatory mechanism for gene expression of BS cell specific C4 genes at their correct sites.
Agrobacterium mediated transformation of immature embryos of indica rice varieties has proven to be highly efficient (Hiei and Komari, 2006). This has made it possible to introduce C4 genes into the indica rice varieties unlike in the past where most of the transgenic plants with C4 genes were developed using japonica varieties such as Kitaake (Ku et al., 1999; Fukayama et al., 2001; Taniguchi et al., 2008). One of the major objectives of developing C4 rice is to enable it to perform efficient photosynthesis under higher temperature and reduced water conditions. As indica rice varieties are more widely cultivated under such conditions, introduction of the C4 pathway is more beneficial. We have chosen IR64, a high yielding indica variety to insert the well characterized C4 genes such as PEPC, PPDK, NADP-ME and MDH from maize following the Agrobacterium mediated genetic transformation protocol of Hiei and Komari (2006). The C4 cycle cannot be established only with these core C4 enzymes (Miyao et al., 2011).
| Table 1: | Summary of genes being transformed into rice to build NADP-ME type of C4 photosynthesis |
 | |
Therefore, we aim to ensure a proper facilitation of exchange of metabolite fluxes into and out of the M and BS cells by inserting the appropriate transporters of the C4 pathway (Table 1). Efficient functioning of the induced C4 cycle in rice will call for a cell type specific suppression of some of the endogenous rice genes. The obvious one is RuBisCO which will be down-regulated in the M cells and over-expressed in BS cells. Other such genes include the subunits of glycine decarboxylase that are involved in the photorespiration.
FOREIGN GENE STABILITY IN THE RICE GENOME
Evidence of stability of alien C4 genes and their expression have been reported by a number of earlier attempts to place C4 genes into C3 plants such as rice (Taniguchi et al., 2008), Arabidopsis (Ishimaru et al., 1997) and tobacco (Gallardo et al., 1995). In rice, several C4 genes have been successfully transformed from maize and other closely related C4 species that have stably integrated into its genome and expressed over several generations (Taniguchi et al., 2008). Furthermore, we have T3 transgenic rice plants harbouring PPDK and PEPC genes from maize that have been constantly expressing the ZmPPDK and ZmPEPC protein over the three consecutive generations. Introducing the C4 genes from phylogenetically close species aids stable integration and expression of the transgenes in the host genome.
PYRAMIDING OF C4 GENES
Transfer of C4 photosynthetic metabolic pathway to non-C4 species would not be complete unless all associated genetic factors are fully inserted in one plant. Multiple transgenes can be stacked in a single plant by crossing of individual transgenic lines and/or by sequential transformation. This strategy of transforming one gene at a time, generating homozygous lines for each and then successive rounds of crossing or sequential transformations to pyramid the necessary C4 genes in rice might prove to be very time and labour consuming process. Moreover, multiple integration sites would further complicate production of homozygous lines. Once each gene has been transformed and tested in isolation, a multigene engineering approach could be very useful to simultaneously transfer many of the C4 genes into rice that would allow us to express multimeric proteins and study the complex genetic regulations. Emerging techniques such as artificial plant chromosome engineering (Naqvi et al., 2009), recombination-assisted multifunctional DNA assembly platform (RMDAP) (Ma et al., 2011), transcription activator like (TAL) effectors (Scholze and Boch, 2011) and zinc finger nuclease (Zeevi et al., 2012) could be applicable. Although none of the above technologies have yet been extensively tested in rice, availability of these novel tools offers new avenues for the C4 rice engineering program.
Transgenic plants with stable and appropriate level of expression of introduced C4 genes (Table 1) will be intercrossed. A prototype C4 rice plant will contain all the known C4 genes. To ensure proper functioning of the engineered C4 cycle, it will require detailed molecular, biochemical and physiological characterizations. The biochemistry will need to be assayed for correct level of enzyme function and carbon fixation in order to confirm a successful C4 cycle activity. The major physiological processes, such as gas exchange and efficiency of photochemistry will need to be analyzed. The proteome, transcriptome and metabolome should also be checked to ensure that the C4 metabolic system is well in place.
CONCLUSIONS
Given the access to advanced technologies and sustainable funding, two decades should be enough time to produce C4 rice. However, the immediate need for the next green revolution, volatile rice markets and intensive media coverage is compelling researchers to try even harder for a faster output. The C4 rice project under the aegis of C4 consortium has successfully completed the first phase during which molecular tools development, infrastructure development, recruitment of scientists and researchers were completed. More importantly, mass screening of sorghum mutants and establishment of efficient rice transformation system were accomplished. The target in coming years is to transform rice with novel C4 genes and pyramid all C4 genes into a prototype either by multigene transfer or by multiple crossing or both.
ACKNOWLEDGMENTS
The research on C4 rice is led by International Rice Research Institute and is financially supported by the Bill and Melinda Gates foundation.
REFERENCES
- Agostino, A., P. Jeffrey and M.D. Hatch, 1992. Amino acid sequence and molecular weight of native NADP malate dehydrogenase from C4 plant Zea mays. Plant Physiol., 98: 1506-1510.
CrossRefDirect Link - Agrawal, G.K., M. Yamazaki, M. Kobayashi, R. Hirochika, A. Miyao and H. Hirochika, 2001. Screening of the rice viviparous mutants generated by endogenous retrotransposon Tos17 insertion. Tagging of a zeaxanthin epoxidase gene and a novel OsTATC gene. Plant Physiol., 125: 1248-1257.
CrossRefDirect Link - Amante-Bordeos, A., L.A. Sitch, R. Nelson, R.D. Dalmacio, N.P. Oliva, H. Aswidinnoor and H. Leung, 1992. Transfer of bacterial blight and blast resistance from the tetraploid wild rice Oryza minuta to cultivated rice, Oryza sativa. Theor. Applied Genet., 84: 345-354.
CrossRefDirect Link - An, S., S. Park, D.H. Jeong, D.Y. Lee and H.G. Kang et al., 2003. Generation and analysis of end sequence database for T-DNA tagging lines in rice. Plant Physiol., 133: 2040-2047.
CrossRefDirect Link - Azpiroz-Leehan, R. and K.A. Feldmann, 1997. T-DNA insertion mutagenesis in Arabidopsis: Going back and forth. Trends Genet., 13: 152-156.
CrossRef - Brar, D.S. and G.S. Khush, 1997. Alien introgression in rice. Plant Mol. Biol., 35: 35-47.
CrossRefPubMedDirect Link - Brown, R.H. and J.H. Bouton, 1993. Physiology and genetics of interspecific hybrids between photosynthetic types. Annu. Revi. Plant Physiol. Plant Mol. Biol., 44: 435-456.
Direct Link - Chen, S., W. Jin, M. Wang, F. Zhang and J. Zhou et al., 2003. Distribution and characterization of over 1000 T-DNA tags in rice genome. Plant J., 36: 105-113.
CrossRef - Chern, C.G., M.J. Fan, S.M. Yu, A.L. Hour and P.C. Lu et al., 2007. A rice phenomics study-phenotype scoring and seed propagation of a T-DNA insertion-induced rice mutant population. Plant Mol. Biol., 65: 427-438.
CrossRef - Danker, T., B. Dreesen, S. Offermann, I. Horst and C. Peterhaensel, 2008. Developmental information but not promoter activity controls the methylation state of histone H3 lysine 4 on two photosynthetic genes in maize. Plant J., 53: 465-474.
CrossRefDirect Link - Droc, G., M. Ruiz, P. Marmande, A. Pereira and P. Piffanelli et al., 2006. OryGenesDB: A database for rice reverse genetics. Nucleic Acids Res., 34: D736-D740.
CrossRefDirect Link - Ehleringer, J.R. and R.K. Monson, 1993. Evolutionary and ecological aspects of photosynthetic pathway variation. Annu. Rev. Ecol. Syst., 24: 411-439.
CrossRefDirect Link - Friso, G., W. Majeran, M. Huang, Q. Sun and K.J. van Wijk, 2010. Reconstruction of metabolic pathways, protein expression and homeostasis machineries across maize bundle sheath and mesophyll chloroplasts: Large-scale quantitative proteomics using the first maize genome assembly. Plant Physiol., 152: 1219-1250.
CrossRefPubMed - Fukayama, H., H. Tsuchida, S. Agarie, M. Nomura and H. Onodera et al., 2001. Significant accumulation of C4-specific pyruvate, orthophosphate dikinase in a C3 plant, rice. Plant Physiol., 127: 1136-1146.
Direct Link - Gallardo, F., M. Miginiac-Maslow, R.S. Sangwan, P. Decottignies and E. Keryer et al., 1995. Monocotyledonous C4 NADP+-malate dehydrogenase is efficiently synthesized, targeted to chloroplasts and processed to an active form in transgenic plants of the C3 dicotyledon tobacco. Planta, 197: 324-332.
CrossRef - Gao, L.Z., B.A. Schaal, C.H. Zhang, J.Z. Jia and Y.S. Dong, 2002. Assessment of population genetic structure in common wild rice Oryza rufipogon Griff. Using microsatellite and allozyme markers. Theor. Applied Genet., 106: 173-180.
CrossRef - Gowik, U. and P. Westhoff, 2011. The path from C3 to C4 photosynthesis. Plant Physiol., 155: 56-63.
CrossRefDirect Link - Hakata, M., H. Nakamura, K. Iida-Okada, A. Miyao and M. Kajikawa et al., 2010. Production and characterization of a large population of cDNA-overexpressing transgenic rice plants using Gateway-based full-length cDNA expression libraries. Breed. Sci., 60: 575-585.
CrossRefDirect Link - Hatch, M.D., 1987. C4 photosynthesis: A unique elend of modified biochemistry, anatomy and ultrastructure. Biochim. Biophys. Acta (BBA)-Rev. Bioenerget., 895: 81-106.
CrossRefDirect Link - Hibberd, J.M. and W.P. Quick, 2002. Characteristics of C4 photosynthesis in stems and petioles of C3 flowering plants. Nature, 415: 451-454.
Direct Link - Hiei, Y. and T. Komari, 2006. Improved protocols for transformation of indica rice mediated by Agrobacterium tumefaciens. Plant Cell Tissue Org. Cult., 85: 271-283.
CrossRefDirect Link - Hirochika, H., 2001. Contribution to the Tos17 retrotransposon to rice functional genomics. Curr. Opin. Plant Biol., 4: 118-122.
CrossRef - Hsing, Y.I., C.G. Chern, M.J. Fan, P.C. Lu and K.T. Chen et al., 2007. A rice gene activation/knockout mutant resource for high throughput functional genomics. Plant Mol. Biol., 63: 351-364.
CrossRef - Huber, S.C. and G.E. Edwards, 1975. Regulation of oxaloacetate, aspartate and malate formation in mesophyll protoplast extracts of three types of C4 plants. Plant Physiol., 56: 324-331.
CrossRefDirect Link - Ishimaru, K., H. Ichikawa, M. Matsuoka and R. Ohsugi, 1997. Analysis of a C4 maize pyruvate, orthophosphate dikinase expressed in C3 transgenic Arabidopsis plants. Plant Sci., 129: 57-64.
CrossRef - Jena, K.K., 2010. The species of the genus Oryza and transfer of useful genes from wild species into cultivated rice, O. sativa. Breed. Sci., 60: 518-523.
CrossRef - Jeong, D.H., S. An, H.G. Kang, S. Moon and J.J. Han et al., 2002. T-DNA insertional mutagenesis for activation tagging in rice. Plant Physiol., 130: 1636-1644.
CrossRefDirect Link - Jeong, D.H., S. An, S. Park, H.G. Kang and G.G. Park et al., 2006. Generation of a flanking sequence-tag database for activation-tagging lines in japonica rice. Plant J., 45: 123-132.
CrossRef - Juneja, S., A. Das, S.V. Joshi, S. Subhash and Y. Vikal et al., 2006. Oryza nivara (Sharma et Shastry) the progenitor of O. sativa (L.) subspecies indica harbours rich genetic diversity as measured by SSR markers. Curr. Sci., 91: 1079-1085.
Direct Link - Jung, K.H., J. Hur, C.H. Ryu, Y. Choi, Y.Y. Chung, A. Miyao, H. Hirochika and G. An, 2003. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol., 44: 463-472.
CrossRefDirect Link - Kajala, K., S. Covshoff, S. Karki, H. Woodfield and B.J. Tolley et al., 2011. Strategies for engineering a two-celled C4 photosynthetic pathway into rice. J. Exp. Bot., 62: 3001-3010.
CrossRef - Kang, H.G., S. Park, M. Matsuoka and G. An, 2005. White-core endosperm Floury endosperm-4 in rice is generated by knockout mutations in the C-type pyruvate orthophosphate dikinase gene (OsPPDKB). Plant J., 42: 901-911.
PubMed - Kolesnik, T., I. Szeverenyi, D. Bachmann, C.S. Kumar and S. Jiang et al., 2004. Establishing an efficient Ac/Ds tagging system in rice: Large-scale analysis of Ds flanking sequences. Plant J., 37: 301-314.
PubMedDirect Link - Kondou, Y., M. Higuchi, S. Takahashi, T. Sakurai and T. Ichikawa et al., 2009. Systematic approaches to using the FOX hunting system to identify useful rice genes. Plant J., 57: 883-894.
PubMed - Kropff, M.J., K.G. Cassman, H.H. van Laar and S. Peng, 1993. Nitrogen and yield potential of irrigated rice. Plant Soil, 155-156: 391-394.
CrossRefDirect Link - Ku, M.S.B., S. Agarie, M. Nomura, H. Fukayama and H. Tsuchida et al., 1999. High-level expression of maize phosphoenolpyruvate carboxylase in transgenic rice plants. Nat. Biotechnol., 17: 76-80.
CrossRefPubMedDirect Link - Ku, M.S., Y. Kano and M. Matsuoka, 1996. Evolution and expression of C4 photosynthesis genes. Plant Physiol., 111: 949-957.
Direct Link - Kwok, P.Y., 2001. Methods for genotyping single nucleotide polymorphisms. Annu. Rev. Genomics Hum. Genet., 2: 235-258.
PubMed - Lake, J.A., W.P. Quick, D.J. Beerling and F.I. Woodward, 2001. Plant development: Signals from mature to new leaves. Nature, Vol. 411.
CrossRef - Langdale, J.A., 2011. C4 cycles: Past, present and future research on C4 photosynthesis. The Plant Cells, 23: 3879-3892.
Direct Link - Lee, S., J. Kim, J.S. Son, J. Nam and D.H. Jeong et al., 2003. Systematic reverse genetic screening of T-DNA tagged genes in rice for functional genomic analyses: MADS-box genes as a test case. Plant Cell Physiol., 44: 1403-1411.
PubMed - Ma, L., J. Dong, Y. Jin, M. Chen. X. Shen and T. Wang, 2011. RMDAP: A versatile, ready-to-use toolbox for multigene genetic transformation. PLoS One, Vol. 6.
CrossRefDirect Link - Matsuoka, M., R.T. Furbank, H. Fukayama and M. Miyao, 2001. Molecular engineering of C4 photosynthesis. Ann. Rev. Plant Physiol. Plant Mol. Bio., 52: 297-314.
CrossRefDirect Link - McCallum, C.M., L. Comai, E.A. Grene and S. Henikoff, 2000. Targeting induced local lesions IN genomes (TILLING) for plant functional genomics. Plant Physiol., 123: 439-442.
PubMedDirect Link - Miyao, M., C. Masumoto, S. I. Miyazawa and H. Fukayama, 2011. Lessons from engineering a single-cell C4 photosynthetic pathway into rice. J. Exp. Botany, 62: 3021-3029.
CrossRef - Molina, J., M. Sikora, N. Garud, J.M. Flowers and S. Rubinstein et al., 2011. Molecular evidence for a single evolutionary origin of domesticated rice. PNAS, 108: 8351-8356.
CrossRef - Monson, R.K., 2003. Gene duplication, neofunctionalization and the evolution of C4 photosynthesis. Int. J. Plant Sci., 164: 43-54.
CrossRef - Mori, M., C. Tomita, K. Sugimoto, M. Hasegawa and N. Hayashi et al., 2007. Isolation and molecular characterization of a Spotted leaf 18 mutant by modified activation-tagging in rice. Plant Mol. Biol., 63: 847-860.
PubMed - Nakamura, H., M. Hakata, K. Amano, A. Miyao and N. Toki et al., 2007. A genome-wide gain-of function analysis of rice genes using the FOX-hunting system. Plant Mol. Biol., 65: 357-371.
PubMed - Naqvi, S., G Farre, G. Sanahuja, T. Capell, C. Zhu and P. Christou, 2009. When more is better: Multigene engineering in plants. Trends Plant Sci., 15: 48-56.
PubMed - Nomura, M., T. Higuchi, Y. Ishida, S. Ohta and T. Komari et al., 2005. Differential expression pattern of C4 bundle sheath expression genes in rice, a C3 plant. Plant Cell Physiol., 46: 754-761.
PubMed - Oleykowski, C.A., C.R.B. Mullins, A.K. Godwin and A.T. Yeung, 1998. Mutation detection using a novel plant endonuclease. Nucleic Acids Res., 26: 4597-4602.
PubMed - Peterhansel, C. and V.G. Maurino, 2011. Photorespiration redesigned. Plant Physiol., 155: 49-55.
CrossRefDirect Link - Quick, W.P., U. Schurr, K. Fichtner, E.D. Shulze, S.R. Rodermel, L. Bogorad and M. Stitt, 1991. The impact of decreased Rubisco on photosynthesis, growth, allocation and storage in tobacco plants which have been transformed with antisense rbcS. Plant J., 1: 51-58.
Direct Link - Qu, S., A. Desai, R. Wing and V. Sundaresan, 2008. A versatile transposon-based activation tag vector system for functional genomics in cereals and other monocot plants. Plant Physiol., 146: 189-199.
PubMed - Ririe, K.M., R.P. Rasmussen and C.T. Wittwer, 1997. Product differentiation by analysis of DNA melting curves during the polymerase chain reaction. Anal. Biochem., 245: 154-160.
PubMedDirect Link - Sage R.F, R.W. Pearcy and J.R. Seemann, 1987. The nitrogen use efficiency of C3 and C4 plants. Plant Physiol., 85: 355-359.
PubMed - Scafaro, A.P., S. Von Caemmerer, J.R. Evans, B.J. Atwell, 2011. Temperature response of mesophyll conductance in cultivated and wild Oryza species with contrasting mesophyll cell wall thickness. Plant Cell Physiol., 34: 1999-2008.
CrossRef - Scholze, H. and J. Boch, 2011. TAL effectors are remote controls for gene activation. Curr. Opin. Microbiol., 14: 47-53.
CrossRef - Springer, P.S., 2000. Gene traps: Tools for plant development and genomics Plant Cell, 12: 1007-1020.
CrossRefDirect Link - Takano, M., H. Kanegae, T. Shinomura, A. Miyao, H. Hirochika and M. Furuya, 2001. Isolation and characterization of rice phytochrome a mutants. Plant Cell, 13: 521-534.
PubMed - Thakur, V., S. Wanchana, M. Xu, R. Bruskiewich, W.P. Quick, A. Mosig and X.G. Zhu, 2011. Characterization of statistical features for plant microRNA prediction. BMC Genomics, Vol. 12.
CrossRef - Till, B.J., J. Cooper, T.H. Tai, P. Colowit, E.A. Greene, S. Henikoff and L. Comai, 2007. Discovery of chemically induced mutations in rice by TILLING. BMC Plant Biol., Vol. 7.
CrossRef - Till, B.J., S.H. Reynolds, C. Weil, N. Springer and C. Burtner et al., 2004. Discovery of induced point mutations in maize by TILLING. BMC Plant Biol., Vol. 4.
CrossRef - Upadhyaya, N.M., Q.H. Zhu, X.R. Zhou, A.L. Eamens and M.S. Hoque et al., 2006. Dissociation (Ds) constructs, mapped Ds launch pads and a transiently-expressed transposase system suitable for localized insertional mutagenesis in rice. Theo. Appl. Genet., 112: 1326-1341.
CrossRef - Viret, J.F., Y. Mabrouk and L. Bogorad, 1994. Transcriptional photoregulation of cell-type-preferred expression of maize rbcS-m3: 3' and 5' sequences are involved. Proc. Natl. Acad. Sci., 91: 8577-8581.
PubMed - Wan, S., J. Wu, Z. Zhang, X. Sun and Y. Lv et al., 2009. Activation tagging, an efficient tool for functional analysis of the rice genome. Plant Mol. Biol., 69: 69-80.
CrossRef - Ware, D.H., P. Jaiswal, J. Ni, I.V. Yap and X. Pan et al., 2002. Gramene, a tool for grass genomics. Plant Physiol., 130: 1606-1613.
Direct Link - Wu, C., X. Li, W. Yuan, G. Chen and A. Kilian et al., 2003. Development of enhancer trap lines for functional analysis of the rice genome. Plant J., 35: 418-427.
PubMed - Wu, J.L., C. Wu, C. Lei, M. Baraoidan and A. Bordeos et al., 2005. Chemical and irradiation-induced mutants of indica rice IR64 for forward and reverse genetics. Plant Mol. Biol., 59: 85-97.
PubMed - Yeo, M.E., A.R. Yeo and T.J. Flowers, 1994. Photosynthesis and photorespiration in the genus oryza. J. Exp. Bot., 45: 553-560.
CrossRef - Zeevi, V., Z. Liang, U. Arieli and T. Tzifira, 2012. Zinc finger nuclease and homing endonuclease-mediated assembly of multigene plant transformation vectors. Plant physiol., 158: 132-144.
PubMed - Zhao, M., Z. Ding, R. Lafitte, E. Sacks, G. Dimayuga and D. Holt, 2010. Photosynthetic characteristics in Oryza species. Photosynthetica, 48: 234-240.
CrossRef - Zhu, Q.H., K. Ramm, R. Shivakkumar, E.S. Dennis and N.M. Upadhyaya, 2004. The ANTHER INDEHISCENCE1 gene encoding a single MYB domain protein is involved in anther development in rice. Plant Physiol., 135: 1514-1525.
Direct Link - Zhu, Q.H., M.S. Hoque, E.S. Dennis and N.M. Upadhyaya, 2003. Ds tagging of BRANCHED FLORETLESS 1 (BFL1) that mediates the transition from spikelet to floret meristem in rice (Oryza sativa L). BMC Plant Biol., 3: 6-18.
CrossRef - Raghavendra, A.S. and R.F. Sage, 2011. C4 Photosynthesis and Related CO2 Concentrating Mechanisms. 1st Edn., Springer, New York, Pages: 410.
Direct Link - Ram, S.G., V. Thiruvengadam and K.K. Vinod, 2007. Genetic diversity among cultivars, landraces and wild relatives of rice as revealed by microsatellite markers. J. Applied Genet., 48: 337-345.
PubMed - Paterson, A.H., J.E. Bowers, R. Bruggmann, I. Dubchak and J. Grimwood et al., 2009. The sorghum bicolor genome and the diversification of grasses. Nature, 457: 551-556.
CrossRefDirect Link - Edwards, G.R., V.R. Franceschi and E.V. Vozsenenskaya, 2004. Single-cell C4 photosynthesis versus the dual-cell paradigm. Annu. Rev. Biol., 55: 173-196.
CrossRefDirect Link