
F. Azam
Nuclear Institute for Agriculture and Biology, P.O. Box 128, Jhang Road, Faisalabad, Pakistan
S. Farooq
Nuclear Institute for Agriculture and Biology, P.O. Box 128, Jhang Road, Faisalabad, Pakistan
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 6 | Page No.: 528-535







ABSTRACT
Form (NH4 or NO3) and availability of N has significant implications to the functioning and sustainability of agroecosystems. Most of the fertilizer nitrogen (N) applied to agricultural soils is in the form of NH4 or NH4-forming fertilizers. This form of N is rapidly oxidized to NO3 by nitrifying microorganisms leading to significant losses of N through NO3-leaching and denitrification. Both denitrification and NO3-leaching have environmental implications and economic concerns. Strategies have therefore been sought to regulate the process of nitrification leading to its complete or partial inhibition. Indeed, climax ecosystems are developed in such a way that the process of nitrification is already fairly inhibited. This paper presents an overview on: I) the process of nitrification, ii) microorganisms involved, iii) the implications of nitrification and nitrification inhibition to ecosystem functioning and finally iv) the methods to inhibit nitrification.
PDF Abstract XML References Citation
How to cite this article
F. Azam and S. Farooq, 2003. Nitrification Inhibition in Soil and Ecosystem Functioning - An Overview. Pakistan Journal of Biological Sciences, 6: 528-535.
DOI: 10.3923/pjbs.2003.528.535
URL: https://scialert.net/abstract/?doi=pjbs.2003.528.535
DOI: 10.3923/pjbs.2003.528.535
URL: https://scialert.net/abstract/?doi=pjbs.2003.528.535
INTRODUCTION
Nitrogen (N) is one of the most widely distributed elements in nature, with atmosphere as the main reservoir. Of the total N found in nature, 99.96% is present in the atmosphere. Of the remaining 0.04%, biosphere contains only 0.005%. In spite of the small proportion found in living beings, N is most often the limiting nutrient for crop production since only a fraction of atmospheric N2 is made available to the plants through biological nitrogen fixation. However, introduction and use of chemical nitrogenous fertilizers has resulted in substantial increases in crop yields. According to statistics by FAO (2001), about 42 million tons of fertilizer N is being used annually on a global scale for the production of 3 major cereal crops i.e., wheat, rice and maize (17, 9 and 16 million tons, respectively). However, fertilizer N is not an unmixed blessing since the crop plants are able to use only about 50% of the applied fertilizer N, while 25% is lost from the soil-plant system through leaching, volatilization and denitrification etc. causing an annual financial loss of ca 3 billion US$. Incidentally, the losses are similar for the 3 crops mentioned above but twice as much from rice compared to wheat and maize when computed on the basis of per unit of fertilizer N applied.
The soil, which is the major source of plant-available N, accounts for a minute fraction of the lithospheric N, while only a very small proportion is directly available to plants as NH4 and/or NO3. Although most plants are able to use both NH4-N and NO3-N, the later is the predominant source of N for plants grown under arable/terrestrial situations. This is primarily because of compulsion rather than preference as most of the NH4-N is rapidly nitrified under optimum conditions of temperature, moisture and aeration. As a result, NH4-N is available to the plants only for a limited time period and most of the time, plant roots are confronted with NO3-N as a source of N.
Nitrification is a key process in managed agricultural ecosystems because the conversion of NH4 to NO3 can lead to substantial loss of agricultural N (both applied as chemical fertilizers or present as native soil N) by leaching and/or denitrification. The loss of fertilizer N is of concern not only because of economic reasons but also due to the pollution potential of different N forms. Some of the adverse environmental effects of excessive use of nitrogenous fertilizers include I) methemoglobonemia in infants due to NO3 and NO2 in waters and food, ii) cancer due to secondary amines, iii) respiratory illness due to NO3, aerosols, NO2 and HNO3, iv) eutrophication due to N in surface waters, v) material and ecosystem damage due to HNO3 in rain water, vi) plant toxicity due to high levels of NO2 and NH4 in soils, vii) excessive plant growth due to more available N and viii) depletion of stratospheric ozone due to NO and N2O. It is because of these concerns that concerted efforts have been and are being made to reduce the use or increase the efficiency of fertilizer N uptake by crop plants.
Multi-disciplinary approaches have been identified and put to practice over the past few decades to increase use efficiency and decrease the losses of fertilizer N. These include a) breeding crop varieties with higher fertilizer use efficiency, b) management of fertilizer N, c) use of chemicals inhibitory to specific N transformation processes like urea hydrolysis and nitrification i.e., urease and nitrification inhibitors, respectively, to slow the process of NH4 formation from urea and to regulate the process of NO3 formation, d) supplementation/integration of fertilizer N with organic manures e.g., green manures. These approaches applied singly or in combination, have certainly helped alleviate the problems arising as a result of fertilizer N use. Fertilizer management through improved formulations, mode and time of application and placement etc. has also been found helpful to overcome some of the problems. Likewise, a large variety of chemicals have been tested as potent inhibitors of specific N transformation processes including urea hydrolysis and nitrification. With these inhibitors, not only use efficiency of fertilizer N is enhanced, but a significant reduction occurs in losses due to NH3 volatilization, denitrification and NO3 leaching. Currently, increasing attention is being paid to the development of controlled-release and stabilized fertilizers that seem more efficient and environment friendly (Trenkel, 1997).
Amongst different N cycle processes vis-à-vis ecosystem functioning and environmental concerns, the process of nitrification stands out to be the most important process. Nitrification inhibition could lead to: i) increased rhizospheric microbial activities including biological N2 fixation and production of phytohormones, ii) enhanced mineralization of native soil N thus decreasing dependence on chemical fertilizers, iii) increased efficiency of fertilizer N through decreased losses via denitrification and NO3 leaching and iv) greater photosynthate partitioning to the rhizosphere thus enriching the soil with organic matter. It is important, therefore, to develop an understanding of microbes responsible for nitrification, factors affecting nitrification, methods to regulate the process and its implications to ecosystem functioning.
Microbiology of nitrification
The biological oxidation of ammonia to nitrate, termed as nitrification, is a two-step process mediated by autotrophic bacteria. Ammonia is first oxidized to NO2– that in turn is oxidized to NO3– as follows:
2NH4+ + 3O2 ⤑ 2HNO2 + 2H+ + 2H2O
2HNO2 + O2 ⤑ 2NO3– + 2H+
The source of NH4+-N could be soil organic matter (mineralization by soil microorganisms) and chemical fertilizers. In the case of soil, organic N is used by the ammonifiers, while chemical fertilizers either contain NH4+ as such or its precursors.
Several genera and species of ammonium and NO2– oxidizing heterotrophs including fungi (Aspergillus flavus, Neurospora crassa, Penicillium sp.), actinomycetes (Streptomycetes sp., Nocardia sp.) and bacteria (Arthrobacter sp., Azotobacter sp., Pseudomonas fluorescens, Aerobacter aerogenes, Bacillus megaterium, Proteus sp.) have been reported. However, autotrophic nitrifiers are generally considered as the main organisms responsible for most of nitrification. Ammonium oxidizing autotrophs include Nitrosomonas, Nitrosolobus and Nitrosospira. These organisms have been isolated from a variety of soil environments with ubiquitous distribution. Nitrite produced by the ammonium oxidizing autotrophs is rapidly oxidized to NO3– by Nitrobacter species. All nitrifiers are obligate aerobes and hence a restricted nitrification under waterlogged or aquatic environments can be observed. In addition, these microorganisms, especially Nitrobacter, are fairly sensitive to acidic pH. As a result nitrification is inhibited in climax ecosystems like forest soils with thick layer of leaf litter and zones of acidic pH (Roberge and Knowles, 1966). The process of nitrification itself may lead to lowering of pH of the medium due to release of H+ as shown in the equation above.
Why to inhibit or regulate nitrification?
As mentioned above, nitrification is one of the key N cycle processes under most arable situations on land. The following discussion will highlight the need to inhibit nitrification in order to maintain the economy of agro-ecosystems.
Nitrification as a source of atmospheric N2O: Concentrations of N2O in the atmosphere have increased markedly in recent years and are continuing to increase at 0.25% per annum (Denmead 1991). This increase is attributed mainly to biospheric processes. Flood irrigation leads to rapid nitrification/denitrification (Bacon et al., 1986) resulting in considerable amounts of N2O being emitted to the atmosphere (Lindau et al., 1990) which may amount to 35-45% of the applied N. On the global level, >65% of the atmospheric N2O comes from the soil, which is twice the amount produced by burning fossil fuels and four times the amount evolved from the oceans. Rates of denitrification in the soil may be as high as 30 kg N lost ha–1 day–1. However, more realistic values could be 3 kg N ha–1 day–1. Being a greenhouse gas (Bouwman, 1990), N2O contributes substantially to the destruction of stratospheric ozone (Crutzen, 1981). Efforts have therefore been devoted to identifying the processes that contribute to N2O emissions and possible mitigation strategies.
Among different processes, nitrification and denitrification are reported to be the main contributors to atmospheric N2O (Bouwman, 1990; Azam et al., 2002). However, since the two processes occur simultaneously (aerobic and anaerobic microsites can develop within the same aggregate, supporting nitrification and denitrification, respectively), it is not easy to ascertain the real contribution of either process to the observed N2O fluxes (Arah, 1997). Nevertheless, nitrification is reported to make a substantial contribution to the N2O emissions under aerobic conditions (Williams et al., 1998). Higher N2O emissions are often reported from fertilized (a routine practice in the present-day agriculture) than unfertilized soils, rates of emission being greatest following application of NH4 or NH4-forming fertilizers (Breitenbeck et al., 1980; Flessa et al., 1996; Azam et al., 2002). In several studies, using isotope methodology and nitrification inhibitors, this increase is attributed to losses of N2O occurring during the process of nitrification (Bremner and Blackmer, 1978; Arah, 1997; Abbasi and Adams, 2000). Estimates of the amount of N2O resulting from nitrification are variable but generally account for <1% of the fertilizer N applied (Breitenbeck et al., 1980). In the case of anhydrous NH3, however, the losses may increase to 6B7% (Smith and Chalk, 1980). In most studies, the onset of N2O emission is observed very early during the incubation, while nitrification continues for extended periods of time (Simarmata et al., 1993). Williams et al. (1998) reported active nitrification 7B12 days after application of NH4NO3, while a flush of N2O emission from soil was observed around day 1, followed by a decline. These authors showed very low molar ratios of NO to N2O and suggested that denitrification was the dominant process involved in N2O emission. Azam et al. (2002) have reached to a similar conclusion from model experiments conducted under laboratory conditions.
Under the agro-climatic conditions of Pakistan, contribution of nitrification to N2O emissions may be higher than generally reported. Urea, which is the major nitrogenous fertilizer in Pakistani agriculture, is rapidly hydrolyzed followed by a quick nitrification of the resultant NH4 especially when applied under relatively warmer conditions. Thus not only nitrification contributes to N2O emissions but the process of denitrification is fairly well supported by sustained availability of NO3. Because of the low organic matter content of the soils that would limit the process of denitrification, a major source of N2O evolved may still be nitrification. However, hardly any work has been reported from Pakistan on these aspects, whereas some good studies have been reported recently on denitrification from wheat-maize cropping systems (Mahmood et al., 1999 and 2000). In most soils, formation and emissions of N2O to the atmosphere are enhanced by an increase in available mineral N through increased rates of nitrification and denitrification. Therefore addition of N in organic or inorganic compounds eventually leads to enhanced N2O emissions; N derived from BNF and returned to the soil in residues is also susceptible to such losses.
Pollution of ground water: Uncontrolled and excessive nitrification may lead to ground water contamination with NO3 and NO2 as well as increased concentration of the later in eatables, especially vegetables leading to human health hazards. Nitrate itself is not a threat while NO2 is definitely a potential health hazard and that too when found at places at a wrong time. In water bodies, however, NO3 and other forms of N may encourage the growth of algae and subsequently the bacteria leading to exhaustion of molecular oxygen thereby affecting animal life. Indeed whole ecological balance of water bodies may change due to the so-called “eutrophication”.
Consumption of water and vegetables containing excessive amounts of NO3 may lead to the production of NO2 in the stomach and the later becomes particularly dangerous for the babies. Methaemoglobonaemia (blue-baby syndrome) may occur in 1-year old babies taking diet with too much NO3. Stomach cancer has also been associated with the concentration of NO3 in potable water. Again, it is NO2 that reacts with amines to form N-nitroso compounds which are reported to cause stomach cancer. Such an illness may result from consumption of vegetables containing high concentrations of NO3 originating from soil or irrigation water.
Nitrification and ecosystem functioning: Nitrification inhibition and consequent accumulation of NH4 would lead to:
| 1 | Increased microbial activities including biological nitrogen fixation, a process that is known to be inhibited more by NO3-N than NH4-N. |
| 2 | Greater photosynthate partitioning to the rhizosphere thus enriching the soil with organic matter |
| 3 | Enhanced mineralization of native soil N through the so-called priming effect or added nitrogen interaction that is more with NH4-N than NO3-N |
| 4 | Increased efficiency of fertilizer N use by plants through decreased losses via denitrification and NO3 leaching |
Microorganisms are known to prefer NH4 over NO3 as a source of N (Jansson, 1958). However, this preference is consequential rather than the reason. This is because of the fact that not all organisms possess nitrate reductase to enable them assimilate NO3, while almost all of them will be able to assimilate NH4. In addition, assimilation of NO3 is more energy intensive as compared to that of NH4. Hence, for efficient assimilation of NO3, ample supply of easily oxidizable C will be required. Studies involving the use of glucose as a C source indeed reveal similar assimilation of both NH4 and NO3 by the soil microorganisms (Azam et al., 1988, 1993). Nevertheless, presence of NH4 leads to an enhancement in microbial activities in terms of respiratory response. In experiments aimed at studying the mineralization of native soil N, NH4-N is reported to have a significantly higher effect as compared to NO3-N. This so-called “priming” effect or added nitrogen interaction (ANI) has frequently been reported using both isotopic as well as non-isotopic methods and is found to increase with the amount of applied N (Jenkinson et al., 1985; Azam, 1990). Azam (1990) suggested that an indirect effect of chemical fertilizers as well as green manures is their positive influence on the mineralization and plant availability of N from the soil organic reserves.
One of the processes reported to be substantially affected by mineral N is the biological N2 fixation. This has been unequivocally proved in legumes at each level of the process starting from nodule initiation to actually fixation at the level of nitrogenase (Ledgard et al., 1987; Blumenthal et al., 1997). The effect is more intense with NO3 than NH4 leading to studies aimed at successful production of NO3 tolerant legume types (Herridge and Bergerson, 1988). This strategy is particularly important under conditions where legumes are grown together with non-legumes and the later are fertilized with chemical fertilizers. The nature has also endowed the rhizobia with the ability to avoid accumulation of inhibitory levels through dissimilatory NO3 reduction i.e., denitrification. Several studies suggest substantial denitrifying ability of these bacteria (Rosen et al., 1996).
Plant species vary in their ability to utilize NH4 or NO3 as the sole source of N although a majority of them grow best with a mixture of the two. However, when used as an exclusive source of N, NH4 may cause growth inhibition in many species, particularly in those grown under arable conditions (Cramer and Lewis, 1993; Marschner, 1999). Under these conditions, nitrification is generally quite rapid and hence deleterious effects of NH4 are avoided. Under saline conditions also, NH4 is reported to increase the sensitivity of plants whereas NO3 has been reported to moderate the negative effects of salinity (Khan et al., 1994). However, the plants are bound to face higher concentrations of NH4 under saline conditions because of the inhibitory effects of salts on the process of nitrification. It would appear therefore that nitrification inhibition would be a blessing for arable plants grown on normal agricultural soils whereas it may be an added problem for those grown on salt-affected lands. Several studies indeed show a positive effect of nitrification inhibitors on plant growth and N use efficiency (Crawford and Chalk, 1993; Lodhi and Azam, 1998; Lodhi et al., 1996 a,b). The mechanisms involved include i) decrease in losses through denitrification and NO3 leaching, ii) increased microbial activities in the presence of prolonged availability of NH4 and iii) increased rhizpospheric activities etc. Nitrification inhibition may not only decrease in the loss of N through denitrification and NO3 leaching, but conserve the applied N through enhanced immobilization of the persistently available NH4-N.
Form of N plays a significant role in affecting root growth, rhizodeposition and the concomitant changes in different rhizospheric microbial functions including root-induced N mineralization. In wheat and maize, root growth may be restricted in NH4 compared to NO3-fed plants and may be attributed to an increased root respiration, greater allocation of photosynthates to nitrogenous than structural component and increased export of carbon (probably as amino acids) from root to shoot than that occurring under NO3 nutrition. In addition, NH4 nutrition leads to a higher rhizodeposition (Giordano et al., 1994) thereby enlarging the below-ground sink for photosynthates, most probably at the expense of plant tops thereby reducing the biomass yield. However, increase in rhizodeposition due to increased/sustained availability of NH4 may also prove beneficial to plants in terms of increased microbial activities and especially the mineralization of native soil N. In laboratory experiments, a significant increase in the mineralization of soil N has been observed following addition of easily oxidizable C (Azam et al., 1989).
It has also been suggested above, that mineralization of N from soil organic matter is more intense in the presence of NH4 than NO3. Jenkinson et al. (1985) attributed this to “pool substitution” whereby the native N stands proxy for the applied N giving the impression of enhanced mineralization of the later. The fact remains, however, that applied N (especially NH4) leads to an increase in the availability of soil N. Inhibition of nitrification may therefore lead to a higher mineralization of native soil N thereby augmenting N supplies to plants. In addition, microorganisms responsible for the synthesis of aggregation-adhesion macromolecules may be encouraged by higher availability of carbonaceous materials in the rhizosphere. This will result in better soil structure as well as improved moisture holding capacity of the soil at the root surface. The later may help the plants withstand drought stress at least temporarily. Thus, in spite of the negative effects of NH4, inhibition of nitrification may still exert beneficial effects on plant growth as mentioned above. The negative effects can be overcome to a significant extent by developing plant types more efficient in using NH4; the so-called “ammoniphilic plants” (Prasad et al., 1983). Plants like rice, sugarcane and kallar grass (Leptochloa fusca) etc. growing under high soil moisture conditions can be considered as ammoniphilic plants. Efforts are needed, however, to engineer arable crops (like wheat) for improved tolerance to NH4 while employing nitrification inhibitors.
Another aspect worth consideration is the susceptibility of NO3 to leaching beyond the effective root zone after being converted to Ca(NO3)2 in the presence of ionic Ca. In Pakistan, the soils are mostly calcareous and hence the conditions are quite conducive to this mode of NO3 escape especially following organic amendment that helps in the release of Ca. The leaching is more pronounced in clayey soils at near neutral pH as negative charge on the clays repels NO3 thereby facilitating the process of leaching. Hence, not only the use efficiency of NO3 will remain low under these conditions but N economy of the system will be negatively affected.
How to inhibit nitrification?
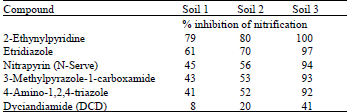
The underlying concept in using nitrification inhibitors (NIs) is to decrease the availability of NO3 and hence its vulnerability to escape mechanisms as the later is directly proportional to the former. A great deal of work has been done and reported on the ways to retard/inhibit the rate of nitrification not only to reduce fertilizer N losses (Aulakh et al., 1984) but also to prolong the persistence of fertilizer N in ammoniacal form (Crawford and Chalk, 1993). In recent years a large number of chemicals, including pesticides, have been used to inhibit nitrification (Bremner and Bundy, 1974; Feng and Barker, 1990; McCarty and Bremner, 1989; Lodhi et al., 1996b). Some effective nitrification inhibitors along with their inhibition potential are listed in Table 1.
Among the different inhibitors, N-Serve and DCD are indeed the most frequently used (Hauck, 1980; Chalk et al., 1990; Bronson et al., 1992; Guiraud et al., 1992; Abbes et al., 1995; Lodhi et al., 1996b; McTaggart et al., 1997). The recommended rate of N-Serve application is 1.1 kg ha–1, while DCD can be applied together with the fertilizer at 1-5% on N content basis. Dyciandiamide is an efficient inhibitor (Rodgers and Ainsworth, 1982). Being nonvolatile, nonhygroscopic and relatively soluble in water (23 g L–1 at 13 °C), it is one of the most convenient inhibitors to be used in formulations with a a variety of fertilizers (Gioacchini et al., 2002).
In addition to the synthetic inhibitors, allelochemics released by plants are also reported to have an inhibitory effect. Rice (1984) postulated that because inhibition of nitrification results in conservation of both energy and nitrogen, vegetation in late succession or climax ecosystems contains plants that release allelochemicals that inhibit nitrification in soil.
| Table 1: | Some commonly used nitrification inhibitors with variable inhibitory effect in different soils (adopted from McCarty and Bremner, 1989) |
 | |
Some natural products like neem (Azadirachta indica) cake are reported to inhibit the activity of nitrifiers (Sahrawat and Parmar, 1974).
An ideal nitrification inhibitor should be mobile, persistent and economical in use. A major consideration during the selection of NIs is their high effectiveness at the lowest possible application rates with a minimum of side effects. Thus the availability of an inhibitor at effective concentrations is essential. This can be achieved by coating fertilizer granules with the inhibitor or by incorporating the latter into granules (Trenkel, 1997; Zerulla et al., 2001). The aim of both the approaches is to ensure an intimate and uniform interaction of the substrate with the inhibitor. The application of an effective concentration of an NI to soil, together with N fertilizer, is a difficult task since it involves different crops, different forms and rate of N application etc. As a result, different concentrations of NI will reach the nitrifiers, particularly if N is applied as granules (Azam et al., 2001). Among the different inhibitors, N-Serve and DCD are indeed the most frequently used (Hauck, 1980; Chalk et al., 1990; Walters and Malzer, 1990; Bronson et al., 1992; Guiraud et al., 1992; Abbes et al., 1995; Lodhi et al., 1996b; McTaggart et al., 1997). Dyciandiamide is an efficient inhibitor (Rodgers and Ainsworth, 1982). Being nonvolatile, nonhygroscopic and relatively soluble in water (23 g L–1 at 13 °C), it is one of the most convenient inhibitors to be used in formulations with a variety of fertilizers (Gioacchini et al., 2002). The inhibitory effect is fairly dependent on the persistence of the chemical in soil. For example, DCD is highly susceptible to degradation at temperatures above 25 °C. Likewise, 50% of the N-Serve may be lost 5 weeks after application to the soil.
Conclusion: While application of chemical fertilizers to agricultural crops has resulted in tremendous increase in yield, problems arising due to the escape to the environment of different nitrogen species, especially N2O, NO2 and NO3, have raised serious economic and environmental concerns. Of the different processes responsible for these concerns, nitrification and denitrification are of prime importance. Hence, efforts have been made to regulate the process of nitrification (major source of different N species) as a means to enhancing the use efficiency of N, decreasing environmental/economic concerns and optimizing the functioning of agro-ecosystems. Use of nitrification inhibitors has been found helpful in mitigating the negative effects of fertilizer application. However, continued efforts need to be made for finding more efficient and environment friendly inhibitors to suit the ever-changing agro-climatic conditions. Fertilizer formulations containing efficient nitrification inhibitors could prove an appropriate way of application.
REFERENCES
- Azam, F., 1990. Comparative effects of organic and inorganic nitrogen sources applied to a flooded soil on rice yield and availability of N. Plant Soil, 125: 255-262.
CrossRefDirect Link - Azam, F., G. Benckiser, C. Mueller and J.C.G. Ottow, 2001. Release, movement and recovery of 3,4-Dimethylpyrazole Phosphate (DMPP), ammonium and nitrate from stabilized nitrogen fertilizer granules in a silty clay soil under laboratory conditions. Biol. Fert. Soils, 34: 118-125.
CrossRefDirect Link - Bronson, K.F., A.R. Mosier and S.R. Bishnoi, 1992. Nitrous oxide emissions in irrigated corn as affected by nitrification inhibitors. Soil Sci. Soc. Am. J., 56: 161-165.
Direct Link - Crawford, D.M. and P.M. Chalk, 1993. Sources of N uptake by wheat (Triticum aestivum L.) and N transformations in soil treated with a nitrification inhibitor (nitrapyrin). Plant Soil, 149: 59-72.
Direct Link - Gioacchini, P., A. Nastri, C. Marzadori, C. Giovannini, L.V. Antisari and C. Gessa, 2002. Influence of urease and nitrification inhibitors on N losses from soils fertilized with urea. Biol. Fert. Soils, 36: 129-135.
CrossRefDirect Link - Lodhi, A. and F. Azam, 1998. Yield and nitrogen uptake of wheat (Triticum aestivum L.) as affected by nitrapyrin and a nitrification inhibiting insecticide. Cereal Res. Commun., 26: 305-312.
Direct Link - Lodhi, A., N.N. Mailk and F. Azam, 1996. Growth and nitrogen nutrition of maize (Zea mays L.) in soil treated with the nitrification-inhibiting insecticidde baythroid. Biol. Fert. Soils, 23: 161-165.
Direct Link - Lodhi, A., N.N. Malik and F. Azam, 1996. Growth and nitrogen nutrition of rice (Oryza sativa L.) in soil treated with N-serve and a nitrification inhibiting insecticide. Pak. J. Bot., 28: 75-83.
Direct Link - Mahmood, T., R. Ali, F. Azam and K.A. Malik, 1999. Comparison of two versions of the acetylene inhibition/soil core method for measuring denitrification loss from an irrigated wheat field. Biol. Fert. Soils, 28: 328-331.
CrossRefDirect Link - McTaggart, I.P., H. Clayton, J. Parker, L. Swan and K.A. Smith, 1997. Nitrous oxide emissions from grassland and spring barley, following N fertiliser application with and without nitrification inhibitors. Biol. Fertil. Soils, 25: 261-268.
CrossRefDirect Link