
Hala I. El-Adawi
Mubarak City for Scientific
Research and Technology
Applications (MuCSAT),
New Borg El-Arab, Alexandria, Egypt
Journal of Medical Sciences
Year: 2006 | Volume: 6 | Issue: 5 | Page No.: 812-817







ABSTRACT
The present study seeked to test the hypothesis that diabetes, characterized by overactivity of the local Rennin-angiotensin System (RAS), leads to signals enhancement and diversification at branching points and thus permits combinatorial interactions between multiple pathways. The streptozotocin (STZ) rat model of type I-diabetes were investigated and confirmed earlier studies that hyperglycemia activates RAS. The pathways discussed in the present study were analyzed using Western blot analysis to investigate the protein density for each pathway. Mitogen-activated Protein Kinase (MAPK) Cascades, showed altered activation in the heart of STZ-diabetic animals. The present study shows, for the first time, that the phosphorylation of Akt (protein kinase B, PKB) is significantly increased as early as 14 days following induction of type-I diabetes. Since Akt plays a critical role in controlling the balance between survival and apoptosis, further research is urgently required for understanding the molecular basis of Akt activity dysregulation which may leads to new therapeutic discovery.
PDF Abstract XML References Citation
How to cite this article
Hala I. El-Adawi, 2006. Early Up-Regulation of Multiple Signaling Pathways in Type I Diabetic Cardiomyopathy. Journal of Medical Sciences, 6: 812-817.
DOI: 10.3923/jms.2006.812.817
URL: https://scialert.net/abstract/?doi=jms.2006.812.817
DOI: 10.3923/jms.2006.812.817
URL: https://scialert.net/abstract/?doi=jms.2006.812.817
INTRODUCTION
Approximately 150 million people worldwide suffer from diabetes (Grundy et al., 1999). Heart failure is the major cause (65%) of death among type I diabetic patients (Geiss et al., 1995). Diabetic Cardiomyopathy has been shown to be a critical factor in heart failure, independent of atherosclerosis, hypertension and vascular malfunction (Rubler et al., 1972; Galderisi et al., 1991) and it has caused shifts in metabolism, cellular biochemistry and structure (Chathanm and Forder 1997; Fang et al., 2004). Cardiomyopathy in diabetic patients is characterized by early diastolic dysfunction, followed by late systolic dysfunction (Rubler et al., 1978; Litwin et al., 1990; Fein and Sonnenblick, 1994). Diabetic Cardiomyopathy has been observed even in insulin-treated type-I diabetic patients (Kannel et al., 1974; Regan et al., 1977). However, the cellular and molecular mechanisms underlying cardiomyopathy and heart failure in type I diabetes are unknown. The serine-threonine kinase, Akt, is a critical intracellular mediator of the actions of insulin and IGF-I (Dudek et al., 1997; Kauffmann-Zah et al., 1997; Kulik et al., 1997). In other settings, Akt activation in the heart counteracts the defects common to diabetic cardiomyopathy by promoting cardiomyocyte survival, improving cellular calcium handling and contractile function (Cittadini et al., 2006; Rota et al., 2005) and enhancing glucose uptake (Hernandez et al., 2001; Gavete et al., 2002). The goal of the present study is to record the Akt activity status at the early development of type-1 diabetic cardiomyopathy. This study is based on the hypothesis that Akt activation in the heart will inhibit diabetes-induced cardiomyocyte apoptosis while mediating significant functional and metabolic benefits in diabetic cardiomyopathy.
Understanding the role of specific signaling pathways in diabetic cardiomyopathy and developing approaches to local modulation of these pathways, may provide novel therapeutic approaches for this important condition.
MATERIALS AND METHODS
Development and characterization of diabetic rats: We investigated the STZ rat model of type I diabetes mellitus. Males Sprague-Dawley rats weighing 150-200 g were made diabetic by a single intraperitoneal injection of STZ (65 mg kg-1). Normal rats of comparable age and weight to be used as controls were injected with vehicle only (100 mM citrate buffer, pH 4.5). Rats were studied 14 days following STZ injection at which time blood glucose concentration was 19.8± 1.5 mM compared to control (4.4±0.3 mM). Similar to previous experience (Xu et al., 1996), the diabetic rats exhibited at a 21% reduction in body weight and 25% reduction in heart weight compared with control but the heart weight-to-body weight ratio was not different from control. Animals were sacrificed for both control and diabetic experimental groups. The Left Ventricle (LV) free wall was dissected and the tissues were rinsed in saline to remove excess blood, snap-frozen in liquid nitrogen and stored at -70°C.
Tissue extracts: To prepare tissue extracts, tissue were washed in PBS and then extracted in lysis buffer containing (in m mol L-1) Tris-HCl (pH7.4) 20, NaCl 100, EDTA 5, NaF 50, Na3 P2 O7 10, Na3VO4I, phenylmethyl/sulfonyl floride 1 and 1.0% Triton X-100, 10% glycerol, 0.1% SDS, 1.0% deoxycholic acid, 10 μg mL-1 aprotinin and 10 μg mL-1 leupeptin. The extract were centrifuged at 10 000xg for 15 min. Protein concentration was determined by the Bio Rad Protein assay.
Protein analysis by immunoblotting: Tissue extract were incubated with 1 μg mL-1 of the antiphosphospecific antibodies (cell Signaling, Beverly, MA) overnight at 4°C. Immunocomplexes were collected by incubating with 50 μL of protein A-or G-Sepharose for 2 h. Immunoprecipitation were washed, resuspended in 2x sample buffer containing 50 m mol L-1. Tris (pH 6.8), 2% SDS, 2% B-mercaptoehanol, 2% glycerol and bromophenol blue. The samples were subjected to SDS-PAGE and were transferred to reinforced nitrocellulose membranes (Schleicher and Schuell). The membranes were blocked with 5% BSA in Tris-buffered saline-Tween solution (20 m mol L-1 Tris-HCL (pH 7.4), 150 m mol L-1 NaCl and 0.05% Tween 20) for 2 h at room temperature. Blots were immunolabelled overnight at 4°C with primary antibody. Bound primary antibody was detected with horseradish peroxidase (HRP)-conjugated secondary antibody (1:2000) and HRP-conjugated antibiotin antibody (1:1000) for 1 h at room temperature. Proteins were visualized by enhanced chemiluminescence (Amersham). Quantitative immunoreactivity was determined by densitometric analysis (Jandel Scientific).
Statisical analysis: Data were expressed as mean±SEM. Comparisons between control and diabetic myocardium were performed by student nonpaired t-test. A value of p<0.05 was accepted as statistically significant.
RESULTS
In order to study the effect of early diabetic condition on multiple signaling pathways we performed IP western blotting to examine the phosphorylation of downstream signals of RAS.
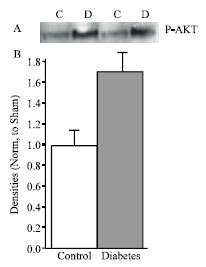 | |
| Fig. 1: | A, Representative Western blot analysis of P-Akt in control (n = 5) and diabetes (n = 5) rats. B, densitometric analysis of Western blot. Data are mean±SEM protein levels normalized to control values |
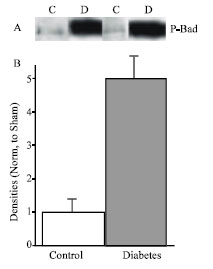 | |
| Fig. 2: | A, representative Western blot analysis of P-Bad in control (n = 5) and diabetic (n = 5) rats. B, Densitometric analysis of western blot. Data are mean±SEM protein levels normalized to control values |
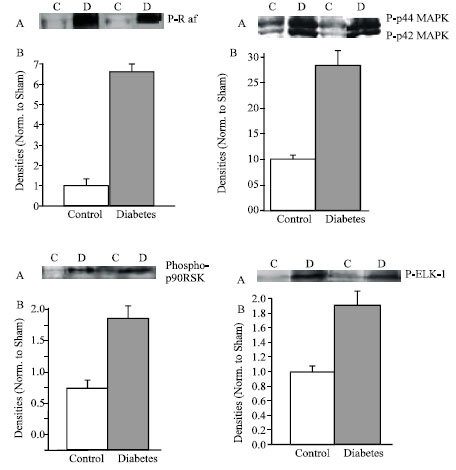 | |
| Fig. 3: | Diabetes upregulated MAPK signaling cascade. A, representative western analysis of p-Raf, p-44/42, phosphor-p-90 RSK and Elk-1. B, Densitometric analysis of western blot. Data are mean±SEM protein levels normalized to control values |
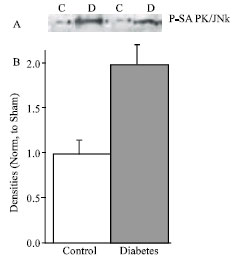 | |
| Fig. 4: | A, Representative Western analysis of p-SAPK/JNK in control (n = 5) and diabetic (n = 5) rats. B, Densitometric analysis of western blot. Data are mean±SEM protein levels normalized to control values |
PI3K pathway: The activation of PI3K leads to the activation of Akt an important player in survival signaling in which the activation of Akt inhibits the proapoptotic BCL2 family member Bad. Phosphorylated Akt was increased by 1.7-folds in diabetic hearts compared to control (Fig. 1). Downstream of Akt we measured the serine phosphorylation of Bad which records increasing to 5-folds in diabetic heart compared with control (Fig. 2).
MAPK signaling cascade: The present study detected as early as two weeks post STZ treatment the activation status of MAPK pathways.
Serine phosphorylation of Raf was highly increased in diabetic myocytes compared to control (6.5-fols). Figure 3 showed that significant increase followed by threonine-ryrosine phosphorylation of p44/42 MAPK recording 2.7 folds. Two downstream targets of p44/42 MAPK have been identified, serine phosphorylation of p90RSK and serine phosphrylation of Elk-1 which recorded an increase to 2.2 folds and 1.9-folds respectively. These findings show that Raf/MEK/ERK pathway was also involved in early regulated signaling diabetic cardiomyopathy.
Next, the study investigated whether Ras transduces signaling through the JNK/SAPK. We detected the threonine-tyrosine phosphorylation of JNK/SAPK by western blot analysis using antiphosphospecific ther183/tyr185 antibodies. The activation reached to 2-folds in diabetic myocytes in comparison with control (Fig. 4).
DISCUSSION
Two weeks after induction of diabetes, the heart is in a compensate stage with remodeled some signal pathways. However, abnormalities of the message expression and protein density of cardiac K+ channel genes, Kv 2.1 Kv 4.2 and Kv 4.3 were detected (Qin et al., 2001). Hyperglycemia activates the local Ras (Fiordaliso et al., 2000; Deng et al., 2004; Machachova et al., 2004), but the understanding of the intracellular signal transduction pathways involved is incomplete. The data presented in this study indicates that overactivity of RAS induced activation of p44/42 through a central role of Ras-Raf-MEK-ERK pathway. That activation transmitted signals to the nucleus where phosphorylation of p90RSK has been detected in addition to the phosphorylation of the transcription factor Elk-1.
These results confirmed the activation of local myocardial RAS which is related to the genesis of diabetic cardiomyopathy (Christlieb and Underwood, 1979; Khatter et al., 1996). As well as emphasized the beneficial effects of RAS blockade in diabetes to reduce the rate of development of hypertension in diabetic patients (Sowers et al., 2001).
Another important signal pathway, Akt, was also studied. It was earlier reported that the activation of Akt was reduced in diabetic heart (Chen et al., 2005; Donthi et al., 2004; Ren et al., 2003). This study shows, for the first time, that the phosphorylation of Akt is significantly increased at the early stage during the development of diabetic cardiomyopthy. Although the potential mechanism of action responsible for reduced Akt activation is still unclear, diminished drive for Akt activation from certain cardioprotective agents such as IGF-1, insulin and gp130-dependent cytokines may be directly responsible for the development of diabetic cardiac mechanical dysfunctions.
Lastly, activation of Bad and its upstream signal Akt and or RSK may dramatically improve cardiac function and protect against apoptosis, establishing an important role for the Akt in the maintenance of heart morphology and function.
Further insight into the mechanism responsible for the dysregulation of Akt phosphorylation may provide new opportunities to design novel therapeutic strategies for diabetic cardiomyopthy.
REFERENCES
- Chen, H.S., Y.X. Shan, T.L. Yang, H.D. Lin, J.W. Chen, S.J. Lin and P.H. Wang, 2005. Insulin deficiency downregulated heat shock protein 60 and IGF-I receptor signaling in diabetic myocardium. Diabetes, 54: 175-181.
Direct Link - Cittadini, A., M.G. Monti, G. Iaccarino, F. Dri Rella and P.N. Tsichlis et al., 2006. Adenoviral gene transfer of Akt enhances myocardial contractility and intracellular calcium handling. Gene Ther., 13: 8-19.
Direct Link - Donthi, R.V., G. Ye, C. Wu, D.A. McClain, A.J. Lang and P.N. Epstein, 2004. Cardiac expression of kinase-deficient 6-phosphofructo-2-kinase/ fructose-2, 6-bisphosphatase inhibits glycolysis, promotes hypertrophy, impairs myocyte function and reduces insulin sensitivity. Biol. Chem., 279: 48085-48090.
Direct Link - Fang, Z.Y., J.B. Prins and T.H. Marwick, 2004. Diabeteic cardiomyopathy: Evidence, mechanism and therapeutic implications. Endocr. Rev., 25: 543-567.
Direct Link - Fiordaliso, F., B. Li, R. Latini, E.H. Sonnenblick, P. Anversa, A. Leri and J. Kajstura, 2000. Myocyte death in streptozotosin-induced diabetes in rats is angiotensin II-dependent. Lab. Invest., 80: 513-527.
PubMedDirect Link - Gavete, M.L., M. Agote, M.A. Martin, C. Alvarez and F. Escriva, 2002. Effect of chronic under nutrition on glucose uptake and glucose transporter proteins in rat heart. Endocrinology, 143: 4295-4303.
Direct Link - Qin, D., B. Huang, L. Deng, H. El-Adawi, K. Ganguly, J.R. Sowers and N. El-Sherif, 2001. Downregulation of K(+) channel genes expression in type I diabetic cardiomyopathy. Biochem. Biophys. Res. Commun., 283: 549-553.
Direct Link - Ren, J., J. Duan, K.K. Hintz and B.H. Ren, 2003. High glucose induces cardiac insulin-like growth factor I resistance in ventricular myocytes: Role of Akt and ERK activation. Cardiovasc. Res., 57: 738-748.
Direct Link - Rota, M., A. Boni, K. Urbanek, E. Padin-Iruegas and T.J. Kajstura et al., 2005. Nuclear targeting of Akt enhances ventricular function and myocyte contractility. Circ. Res., 97: 1332-1341.
Direct Link - Sowers, J.R., M. Epstein and E.D. Frohlich, 2001. Diabets, hypertension and cardiovascular disease: An update. Hypertension, 37: 1053-1059.
Direct Link