
J.O. Olomukoro
Department of Animal and Environmental Biology, University of Benin, P.MB. 1154, Benin City, Nigeria
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 2 | Page No.: 309-314







ABSTRACT
The Macrobenthic community assemblage associated with Eichhornia crassipes (water hyacinths) was investigated at two sampling stations of Warri River. One hundred and twelve species of benthic organisms in forty-one families were identified. All except mollusca group are well known freshwater forms which are limited in longitudinal distribution to areas with salinity below 1.02%. Ephemeroptera, Coleoptera, Trichoptera and Arachnida were restricted to the freshwater areas (upstream station 1) with salinity values of 0.02-0.46%. Dominant groups are Oligochaeta (Naididae), Ephemeroptera (Baetidae) and Diptera (Chironomidae) with 13, 10 and 9 taxa, respectively. In this respect, they are considered to be the most sensitive groups in water hyacinths to salinity gradation in Warri River. Therefore they serve as very useful bioindicators of environmental change including human-induced stress factors in aquatic ecosystem.
PDF Abstract XML References Citation
How to cite this article
J.O. Olomukoro, 2007. Salinity and the Macrobenthic Community Structure in Eichhornia crassipes of Warri River, Nigeria. Journal of Biological Sciences, 7: 309-314.
DOI: 10.3923/jbs.2007.309.314
URL: https://scialert.net/abstract/?doi=jbs.2007.309.314
DOI: 10.3923/jbs.2007.309.314
URL: https://scialert.net/abstract/?doi=jbs.2007.309.314
INTRODUCTION
Eichhornia crassipes (Water hyacinths) grow over a wide variety of wetland types from lakes, ponds, streams, waterways, ditches and backwater areas (Grodowitz, 1998; Home Back, 2004) and have constituted a severe environmental and economic problem to the Gulf of Guinea (West African coast) over the past four decades. Water hyacinths, although an environmental nuisance, can destroy native wetlands and waterways, degrades water quality and dramatically alters native plants and animal communities killing native fish and other wildlife (Land Protection, 2004), have been found to obtain their nutrients directly from the water and currently used in waste water treatment facilities (Dempster and Manning, 1991) and also termed as a biological museum (Egborge, 1980).
Benthic organisms are plants and animals, which live on/or in bottom sediments or are attached to solids objects (living or non-living), floating on, or partly/completely submerged in water. In freshwater environment over 50% of benthic organisms live on the hanging roots of floating aquatic plants such as the water hyacinth, E. crassipes (Egborge, 1994). Egborge (1998) work on water hyacinth was the earliest and the only existing hitherto describing an array of animals (of over 150 different species) on the roots of water hyacinth in the Badagry Creek. Egborge (1994) observed that fish would more easily obtain food items concentrated on the roots of a mat of flooding weeds.
Benthic macrofauna species are important as food for economically important fish and shellfish species in most aquatic environment where they are the major secondary producers (Ajao and Fagade, 2002). They have been employed as indicators of organic pollution and other human-induced stress factors in aquatic ecosystem (Ajao and Fagade, 1990).
Since reports on Floating aquatic macrophytes associated animals of Nigeria are generally lacking although such studies may be very essential for aquatic weed biological control strategies particularly in the Niger Delta where they constitute problems in the waterways. Therefore the purpose of the present study was to establish a baseline reference source and highlight the benthic community structure of water hyacinth vital for teaching and research.
Study area: Warri River (Long. 6°00’E, Lat. 5° 20’N) is one of the most important coastal rivers of the Niger Delta of Nigeria. A detailed description of this river has been given elsewhere (NEDECO, 1961; Oronsaye, 1996; Olomukoro and Egborge, 2003, 2004).
Station 1 (at Agbarho) is a permanent freshwater part of the Warri River. Station 2, (at Aladja), is not entirely fresh throughout the year, but become brackish due to incursion of marine waters from Forcados during the dry season months (December-March).
MATERIALS AND METHODS
E. crassipes were sampled for benthos from January 2002 to May 2003. The floating macrophytes were collected from the mid-stream and dusted in a white polypropylene bucket containing ten percent formalin. Prior to dusting, each macrophyte is allowed to remain in the prepared medium for 3-5 min. There after, the macrophytes were again transferred to another bucket of water and dusted vigorously. The contents of each bucket were filtered through a set of Tyler sieves of 2 and 1 mm, 150 and 100 μm. Macrobenthic organisms retained in the filters were washed into polypropylene bottles and preserved in five percent formalin. The partially decayed leaves, roots and stolons of the plants were thoroughly searched to secure organisms clinging to them.
Chloride determination was by the argentometric method. From chlorinity, salinity values were obtained by titrimetric method and the titrations were done on a magnetic stirrer.
Sorting and identification of organisms to the lowest possible taxonomic level were done using an American optical dissecting microscope, model 570 and HM-Lux 3 microscope. Identification of organisms was possible by using appropriate keys and works of Pennak (1978), Macan (1959), Word and Whipple (1959), Mellanby (1963), Hynes (170) and Needham and Needham (1978) and Brinkhurst (1966).
Data analysis: Estimation of monthly species richness in each station was carried out using Margalef’s index of species diversity (Clifford and Stephenson, 1975). The system of Slack et al. (1977) was used in the determination of abundant, dominant and rare group of genera. Statistical analysis, which is the measurement of correlation coefficients between salinity and major groups of benthic macroinvertebrates was computed (Brase and Brase, 1987).
RESULTS
Benthic fauna in E. crassipes: The macro-invertebrate communities associated with E. crassipes of Warri River consists of 119 species in forty-one families. These are reflected in Table 1. Three groups of benthic organisms were dominant in E. crassipes at the study stations. These are Oligochaeta (Naididae), Ephemeroptera (Bactidae) Diptera (Chironomidae) and Mollusca (Hydrohiidae) with 13, 10, 9 and 5 taxa, respectively. Of all the total assemblage of benthic macro-invertebrates recorded, Mollusca made up the dominant group in the studied area, representing 48.62% of the benthic population. This was followed by the Decapoda, Oligochaeta, Diptera and Ephemeroptera accounting for 17.25, 14.44 and 6.86% respectively. Other groups- Odonata, Coleoptera, Trichoptera and Arachnida were relatively unimportant.
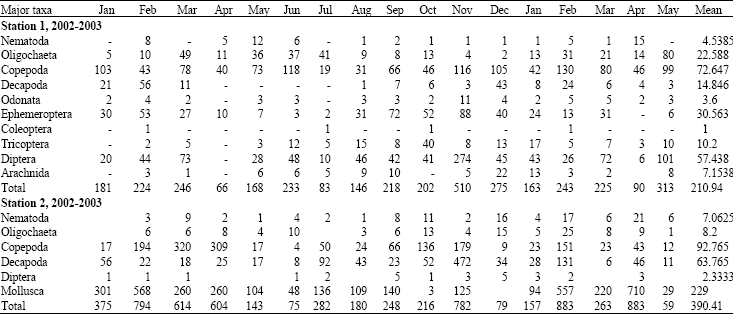
Seasonality of benthos: The seasonal variations in the population density of the dominant groups of benthic invertebrates are shown in Table 2.
At station 1, Oligochacta occurrence was significant in most months in E. crassipes. Although its appearance was pronounced in March and July 2002 and May 2003, it was relatively less important between August and December 2002. Ephemeroptera showed high fluctuations in density in E. crassipes. The population density was high in January to March, 2002 and August to January 2003. Low density was recorded between February and July 2002. It was dominant in the habitat at the end of the rainy season. Diptera, like Ephemeroptera, showed high monthly fluctuations in E. crassipes. The relative importance of Diptera among the benthos is shown in Table 1. Dipteran larvae were predominant throughout the study period with high density in March, November 2002 and March, May 2003. In this station, Odonata, Trichoptera, Coleoptera and Arachnida were more significant than abundant between August 2002 and January 2003.
At the brackish water of station 2, Oligochaeta occurrence was significant in most months in 2002 and 2003 (Table 2). While no appearance of Ephemeroptera nymph was recorded in E. crassipe, the occurrence of Diptera was quite insignificant throughout the months of sampling. Extremely low population density of diptera was recorded in all the biotope during the dry and rainy season months. Their appearance was more minima than maxima during the study period.
Decapoda was as important as Gastropoda in the E. crassipes where they were more often present and abundant during the seasons. The latter made astronomic appearances in January to August 2002 and December 2002 to May 2003.
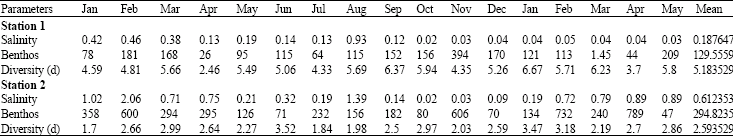
Salinity variation and benthos: The seasonal variation in salinity of the stations during 2002/2003 is shown in Table 3. Salinity varied from 0.03% at station 1 (Agbarho) to 2.00% at station 2 (Aladja). The peak period of salinity was February and August 2002 when salinities were 0.93 and 2.06% at Agbarho and Aladja, respectively. There was a gradual increase of salinity from fresh water salinities at the upstream station 1 to fresh/brackish water at the downstream station 2. At the former station the peak of benthic fauna abundance corresponds to the period salinity values were at their minimal levels (Table 3). The high density of benthos in February and November 2002 and February and April 2003 has no direct relationship with salinity variations in the downstream station 2. Also, the overall benthic fauna abundance is observed to be influenced by the corresponding high increase in the density of Mollusca known to be restricted to the latter station (Table 1).
Diversity: Margalef’s diversity index (D) which is the measurement of the number of individuals and gives information on species richness was used to assess monthly diversity variation in each station (Table 3).
| Table 1: | Composition, abundance and distribution of Macrofauna in E. crassipe of Warri River, 2002-2003 |
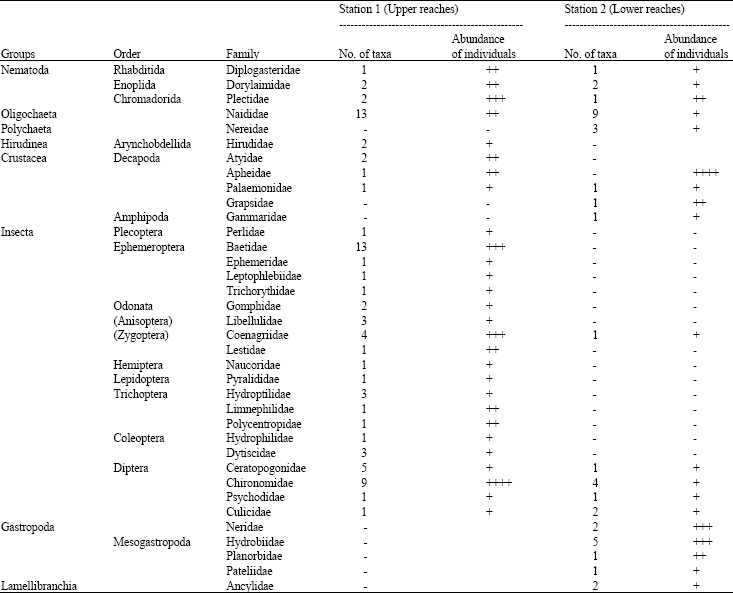 | |
+ = Rare (<10 indiv), ++ = Common (>10 indiv.), +++ = Dominant (>100 indiv.), ++++ = Abundant (>500 indiv.) | |
| Table 2: | The abundance and seasonal variation of major groups of benthic macroinvertebrates in E. crassipe of Warri River 2002-2003 |
 | |
| Table 3: | Salinity variation, benthos abundance and diversity (D) in the study stations of Warri River, 2002-2003 |
 | |
| Table 4: | Correlation coefficient values between salinity and major groups of benthic fauna in the stations |
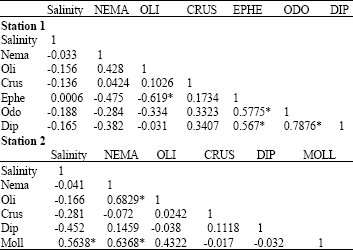 | |
Significant relationships are asterisked | |
At station 1, index values range from 2.46 to 6.67. The highest diversity was recorded in January, 2003; while the month of April, 2002 had lowest diversity of 2.46. Station 2 however showed relatively low diversity in few months with values ranging from 1.70 in January, 2002 to 3.52 in June of the same year. However, monthly fluctuations in diversity in the study stations are quite obvious. Highest diversity was recorded in both stations in the dry season.
Correlation coefficient analyses: Correlation coefficient analyses of salinity variables and major groups of benthic macroinvertebrates in each station were computed (Table 4). At station 1, salinity was negatively insignificantly correlated with all the major groups-Nematoda, Oligochaeta, Crustacea, Ephemeroptera, Odonata and Diptera. Among the organisms, positive significant correlations were established between Ephemeroptera and Odonata/Diptera and between Odonata and Diptera.
At station 2, salinity had negative correlation with all the benthic groups except Mollusca, which was significantly correlated (0.56377) with salinity. However, positive relationships were established between Nematoda and Oligochaeta/Mollusca.
DISCUSSION
Salinity ranges indicate freshness at certain times of the year and brackishness at other times. Reid (1961) has observed the tendency for salinities to remain fresh even at the mouth of estuaries for large rivers like Amazon and Mississippi. The seasonality in the fluctuation in salinity is apparent, with higher salinity values in the dry season months, particularly at station 2. The low salinity values during the rainy season per months could be attributed to the dilution of the river waters by masses of freshwater from the enormous tributaries in the area (Egborge, 1987; Oronsaye, 1996). The high chloride content may indicate also that water is being made saline by encroachment of seawater (Jackson et al., 1989).
Apparently salinity showed significant difference in the two stations. With this, the macro invertebrates are subjected to salinity fluctuations of 0.95 to 2.06% at Agbarho (1) and Aladja (2) stations. Salinity variation may be a limiting factor in the distribution of benthos, particularly the insecta groups. One other important controlling factor is the amount of dissolved salts in the water especially calcium carbonate which is the essential materials for shell construction in shell fishes. This distribution pattern is not surprising as molluscan are prevalent in brackish waters, streams or lakes whose surface waters are less acid than pH 6.2 (Pennak, 1978).
The Zoobenthic communities associated with the water hyacinth, E. crassipes at station 2 are admixtures of fresh and brackish water species. The brackish nature of the environment is further confirmed by the presence of well-known brackish water forms such as the errant polychaete, Nereis sp., the mollusc, Potamopyrgus fuscatus, P. radula, Neritina glabrata and the crab, Sesarma alberti. Diversity increase in most months was observed in the freshwater compared to the brackish environment.
High diversity results when many species have equal or near equal opportunity of co-existence. A decrease in diversity and corresponding increase in abundance of available species is a prevalent community response to environmental disturbance. Therefore, low diversity is an indication of stress in the environment while high diversity is a reflection of stress-free environment. According to Mackie, 1998, the longer a locality has been in the same condition the richer is its biotic community and the more stable it is.
Hydrobiological investigations revealed the inverse correlation of salinity with all the major groups of benthic organisms except mollusca which was particularly restricted to the brackish downstream station 2. The complete absence of Ephemeroptera, Odonata, Coleoptera, Trichoptera and Arachnida in the latter station shows that they cannot tolerate any slightest change in salinity value beyond the freshwater concentration. However, Olomukoro (1996) has reported a single collection of Baetis sp. (Ephemeroptera) in the brackish environment during the wet season months. Baetis sp. is known to tolerate any environmental change to an extent compared to other may fly larvae. It appears that other factors other than salinity determines the densities of benthic organisms in E. crassipes of Warri River. Grazing therefore may be an important factor in seasonal variations and interrelationships among the species. A reciprocal relationships between the population densities of Odonata and Ephemeroptera/Diptera indicates that Odonata grazing of Ephemeroptera and Diptera controlled the population size of the latter groups. The same pattern of relationship exist between Mollusca and Nematoda.
Also, the inverse correlation between Ephemeroptera and Oligochaetes in E. crassipes could be that Oligochaetes are generally utilized as a food source, although the feeding relationships were not experimentally attempted in this study.
The whole array of common and rare zoobenthos on the host of E. crassipes provides an opportunity for research, especially with respect to the economic and practical utility in fish culture practices of the communities of the zoobenthic organisms which comprise the food spectra of detritophagous and omnivorous fish and which are also of relatively big size for favourable discriminatory selection by fish.
ACKNOWLEDGMENT
I am grateful to Dr. Graham De Heaume of the Preparatory Department of the International Institute of Entomology, London, who provided useful texts for the identification of the specimens.
REFERENCES
- Egborge, A.B.M., 1987. Salinity and the distribution of cladocera in Warri River, Nigeria. Hydrobiologia, 145: 159-167.
CrossRefDirect Link - Egborge, A.B.M., 1994. Water Pollution in Nigeria: Biodiversity and chemistry of Warri River. Ben Miller Books, Nigeria,.
Direct Link