
T. Eslamzadeh
Department of Biology, College of Sciences Shiraz University,
Shiraz, 71454, Iran
International Journal of Soil Science
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 75-80







ABSTRACT
Osmotic potential, percent of ash, mineral salt content and heavy metals of the halophyte Salicornia europea, collected from Maharloo region, Shiraz, were determined. Sodium content in ash was found to be up to 30%, equivalent to 31500 μg g-1 dry matter. The high sodium content of the plant especially in the shoot as compared with normal content of 10 μg g-1 in non-halophytes suggests that the plant can be considered as a sodium hyperaccumulator and it can be used as a means of phytoremediation for saline media, including soil and water. The content of mineral nutrients K, Mg, Ca, Zn and Cu were also determined and were compared with the normal amounts needed for plant growth. The results indicate that mobile elements such as Na, K and Mg are mostly transported to the shoot, where, they may be compartmentalized in the leaf cell vacuoles; whereas the immobile elements such as Ca, Cu, Zn and Fe are mainly retained in the root. The amount of Mg in the shoot and also the amounts of copper and iron in the root are much higher than the normal amount found in other plants; indicating that the root tissues of the plant can be further studied to be used as biosorbent for heavy metal contaminants in wastewater and soil. The amounts of heavy metals such as Ni, Cr, Cd, Pb and Hg were also determined. The results indicate that the amounts of these heavy metals are relatively high in this plant.
PDF Abstract XML References
How to cite this article
T. Eslamzadeh, 2006. Salicornia europea, a Bioaccumulator in Maharloo Salt Lake Region. International Journal of Soil Science, 1: 75-80.
DOI: 10.3923/ijss.2006.75.80
URL: https://scialert.net/abstract/?doi=ijss.2006.75.80
DOI: 10.3923/ijss.2006.75.80
URL: https://scialert.net/abstract/?doi=ijss.2006.75.80
INTRODUCTION
With respect to salt tolerance, there is a wide spectrum of plants ranging from very sensitive glycophytes showing the effects of salt injury at a concentration of less than 50 mM; to halophytes that complete their life cycles at a salt concentration of as high as 500 mM (Clipson et al., 1985; Ungar, 1999; Volkmar et al., 1997). Unfortunately, the major economical plants are almost universally non-halophytic. For example, bean yield is inhibited almost completely at 50 mM NaCl (Mass and Grieve, 1987). Usually there is a threshold concentration of salt above which glycophytes begin to show signs of growth inhibition, leaf discoloration and loss of dry weight. Among crops, maize, onion, citrus, pecan, lettuce and bean are highly sensitive to salt; cotton and barley are moderately tolerant; and sugar beet and date palms are highly tolerant (Greenway and Munns, 1980).
Plant mineral nutrition, a much older field of study than phytoremediation, has recently focused on enhancing the ability of plants to accumulate mineral nutrients from the environment more efficiently. This effort has been directed at both reducing the amount of mineral fertilizer input into agricultural system and at alleviating human diseases caused by deficiencies in plant-derived from deficiency of essential and beneficent micro nutrients, such as Fe, Zn and Se. The interesting convergence of these two fields has led to realization that phytoremediation and enhanced mineral nutrition are two sides of the same coin (Salt et al., 1988). Plants possess different potential mechanisms that may be involved in the detoxification of heavy metals and thus tolerance to metal stress. Some of these mechanisms are controlled at the cellular level (Hall, 2002; Volkmar et al., 1997; Hasegawa et al., 2000 ) and some, at whole plant (Cobbet et al., 2000 ). Salicornia europea is a halophyte living around salt marshes. In the present study, the salt content of this plant ash has been determined both in the root and shoot. The aim of the research is to study the potential of this plant to accumulate salts and its capacity to be used as a means of phytoremediation in saline water and soil. Being consumed as a source of food by both human and animals, it is also a valuable source of essential micronutrients such as, Fe and Zn, protein, determining the levels of heavy metals in this plant is of interest to animal nutritionists.
MATERIALS AND METHODS
This study was conducted in 2002 in Biology Department, College of Sciences, Shiraz University, Iran.
Salicornia plants were collected from the vicinity of Maharloo salt lake near Shiraz, in Fars Province. After washing the wet soil particles from the Plant surface, leaf cell sap was extracted and its osmotic potential was determined by cryoscopic method. Both shoot and root fresh and dry weights were determined. For drying, plant materials were kept in an oven set at 75°C for 24 h. To determine the mineral content,
The dry tissues were ashed in a furnace set at 600 °C for 8 h. The percentage of dry matter and ash were determined for both shoot and root. Mineral content in shoot and root ash was determined by both XRF and atomic absorption spectrophotometer.
RESULTS AND DISCUSSION
The osmotic potential of leaves cell sap as determined by cryoscopic method, was about -8.5 MPa, which is much lower than the amount reported for glycophytes (-1 to -3 Mpa) indicating the presence of a very high soluble organic and inorganic materials. The shoot and root dry weight and the percentage of ash are shown in Table 1 and some of the elements present in both shoot and root ashes are shown in Table 2. There was a considerable amount of sodium both in shoot and root ashes, amounting to 30 and 24% of ashes in these tissues respectively, corresponding to 31500 and 11040 μg g-1 dry weight. For C4 and CAM plants, Na+ is a required macronutrient (10 μg g-1 dry weight) needed for regeneration of phosphoenolpyruvate in C4 and CAM Pathway (Johnstone et al., 1988). In some halophytes sodium can replace potassium but not completely in its physiological functions. Since NaCl is the most abundant salt encountered by plants under salinity stress, transport systems that facilitate compartmentation of Na+ I vacuole are of critical importance (Binzel, 1988). In this study the sodium content in dry matter of the shoot is much higher than the root (Table 2).Na+ is mainly transported to the shoots to be accumulated in the leaf vacuoles. The succulent feature of this plant leaves is apparently due to this high accumulation of Na+. Vacuolar compartmentation of Na+ probably results in part from the activity of a family of Na+-H+ antiporters (Aspe, 2002). According to Table 2 and 3, only a small portion of absorbed K+ is transported to the shoot. The results also show much higher Na+ content than K+ both in root and shoot. Na+ has been shown to compete with K+ in inward rectifying channels (Anderson et al., 1999).
| Table 1: | Content of dry matter and ash in root and shoot of Salicornia europea % |
| Table 2: | Mineral content in dry matter of Salicornia europea (μg g-1) compared with normal content in plant dry matter |
 | |
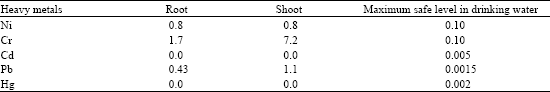
| Table 3: | Heavy metal content in cell sap of root and shoot of Salicornia europea (μg g-1) |
 | |
Other reports have suggested K+ outward rectifying channels might facilitate Na+ influx as well. The high affinity K+ transporter (HKT) from wheat (Schatman et al., 1991) and low affinity cation transporter (LCT1) from wheat (Schatman et al., 1997) facilitate Na+ transport with isolated membranes, providing evidence to support their possible function in Na+ uptake since high affinity K+ transporter1(HKT1) transcripts are expressed predominately in the roots, it most probably functions in the control of Na+ flux into the xylem for export to the shoot. Both Ca2+ and K+ affect intracellular Na+ concentration (Zhong and Lauchli, 1994).
The elements Ni, Cr, Cd, Pb and Hg were not detectable in ash, so they were checked in cell sap by means of atomic absorption spectrophotometer (Table 3).
As shown in Table 3, Cd and Hg were not detected in cell sap. The amount of Ni was shown to be 0.8 μg g-1 both in the root and shoot cell sap, which is 8 times the permissible amount in drinking water. The amount of Cr was shown to be up to 7.2 μg g-1 (72 times) and Pb 1.1 μg g-1 (1000 times) as compared with the permissible level in drinking water. So, the amounts of these heavy metals in the cell sap may be toxic for animal consumption. Many soil trace minerals such as Cu, Fe, Mn, Ni and Zn are absorbed by plant roots. Although these elements are essential (Epstein, 1972, 1999) they can also be potentially toxic. Plants possess complex mechanisms to control them. For reasons not yet clear, plants also have the ability to acquire and detoxify non-essential elements such as Cd, Cr and Pb (Van Hoof et al., 2001).
The majority of plants grown in metal enriched soils have mechanisms that exclude these potentially toxic metals from their shoots using their roots to either pump metal ions back into the soil (Van Hoof et al., 2001), or store them in their root vacuolar compartment. There are also much evidence showing that salinity increases intracellular Ca2+ concentration. Increased intracellular Ca2+ concentration in corn root protoplast (Lynch et al., 1989 ) in response to salinity, has been attributed to the role of calcium in signal transduction (Reid et al., 1993). In unicellular alga Duniella salina, Ca2+ absorption from external medium increases and the magnitude of this increase depends on the degree of salt stress (Ko and Lee, 1995). Other reports indicate increased cytosolic Ca2+ concentration in barley root protoplast and also in cells of wheat aleurone layer in response to salinity (Bittisnich et al., 1989; Bush, 1996). H+ pumps in the plasma membrane and tonoplast provide the driving force for secondary transport of ions. A vacuolar H+-ATPase generates aΔ pH and membrane potential across the tonoplast. Activity of these pumps is required for secondary transport of ions associated with plant responses to salinity stress; since the activity of these H+- pumps is increased by salinity (Hasegawa et al., 2000). Vacuolar compartmentation of Na+ results in part from the activity of a family of Na+/ H+ antiporters (Aspe, 2002). Little is known about the mechanism of metal translocation to the shoot, though it has been suggested that coordination of the metal ion with free histidine may play a role (Kramer et al., 1996). Within these cells metal ions are detoxified by compartmentalization in the vacuoles (Kupper et al., 1999; Kramer et al., 2000 ), where they are most likely coordinated by organic acids (Salt et al., 1998; Taiz and Zeiger, 2002) the succulent feature of the plant might be an evidence for this. Recently researchers have focused their attention on the various metal hyperaccumulating species, which are close relatives of Arabidopsis thaliana (Macnair et al., 1999B). Hyperaccumulators are species capable of accumulating metals at levels 100-fold greater than those typically measured in shoots of the common non-accumulator plants (Lasat, 2002; Baker et al., 2000). Emerging knowledge will hopefully lead to identification of new plants useful for remediation of metal polluted soils as well as plants with enhanced mineral composition for improved human nutrition and health (Taiz and Zeiger, 2002). This plant is used as a salad vegetable and sometimes as main dish together with mushroom, fish and spices. Having the potential to accumulate heavy metals from the surrounding, one should make sure of its suitability as a food source when grown in polluted habitats like Maharloo Salt Lake region. Our unpublished data has shown that it can also be considered as a source of protein.
ACKNOWLEDGMENT
Great appreciation is offered to Professor B. Kholdebarin for his kindly valuable guidance.
REFERENCES
- Apse, M.P. and E. Blumwald, 2002. Engineering salt tolerance in plants. Curr. Opin. Biotechnol., 13: 146-150.
CrossRef - Baker, A.J.M., S.P. McGrath, R.D. Reeves and J.A.C. Smith, 2000. Metal Hyperaccumulator Plants: A Review of the Ecology and Physiology of a Biological Resource for Phytoremediation of Metal-Polluted Soils. In: Phytoremediation of Contaminated Soil and Water, Terry, N. and G. Banuelos (Eds.). Lewis Publishers, Boca Raton, FL., pp: 85-108.
Direct Link - Binzel, M.L., F.D. Hess, R.A. Bressan and P.M. Hasegawa, 1988. Intracellular compartmentation of ions in salt adapted tobacco cells. Plant Physiol., 86: 607-614.
CrossRefDirect Link - Bush, D.S., 1996. Effects of gibberellic acid and environmental factors on cytosolic calcium in wheat aleurone cells. Planta, 199: 89-99.
Direct Link - Cobbett, C.S., 2000. Phytochelatins and their roles in heavy metal detoxification. Plant Physiol., 123: 825-832.
CrossRefDirect Link - Epstein, E., 1999. Silicon. Annu. Rev. Plant Physiol. Plant Mol. Biol., 50: 641-664.
CrossRefPubMedDirect Link - Greenway, H. and R. Munns, 1980. Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol., 31: 149-190.
CrossRefDirect Link - Hall, J.L., 2002. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot., 53: 1-11.
CrossRefDirect Link - Hasegawa, P.M., R.A. Bressan, J.K. Zhu and H.J. Bohnert, 2000. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol., 51: 463-499.
CrossRefDirect Link - Johnstone, M., C.P.L. Grof and P.F. Brownell, 1988. The effect of sodium nutrition on the pool sizes of intermediates of the C4 photosynthetic pathway. Plant Physiol., 15: 749-760.
Direct Link - Kramer, U., J.D. Cotter-Howells, J.M. Charnock, A.J.M. Baker and J.A.C. Smith, 1996. Free histidine as a metal chelator in plants that accumulate nickel. Nature, 379: 635-638.
Direct Link - Lynch, J., V.S. Polito and A. Lauchli, 1989. Salinity stress increases cytoplasmic calcium activity in maize root protoplasts. Plant Physiol., 90: 1271-1274.
Direct Link - Macnair, M.R., S.B. Huiston, L. Saumitou and D. Petit, 1999. Zinc tolerance and hyperaccumulation are genetically independent characters. Proc. R. Soc. lond., 266: 2175-2179.
Direct Link - Reid, R.J., M. Tester and F.A. Smith, 1993. Effects of salinity and turgor on calcium influx in chara. Plant Cell Environ., 16: 547-554.
Direct Link - Salt, D.E., R.D. Smith and I. Raskin, 1998. Phytoremediation. Annu. Rev. Plant Physiol. Plant Mol. Biol., 49: 643-668.
CrossRefDirect Link - Schatman, D.P., S.D. Tyreman and B.R. Terry, 1991. The K+/ Na selectivity of a cation channel in the plasma membrane of root cells does not differ in salt tolerant and salt sensitive wheat species. Plant Physiol., 97: 598-605.
Direct Link - Van Hoof, N.A.L.M., P.L.M. Koevoets, H.W.J. Hakwoort, W.M. Bookum, H. Schate, J.A.C. Verkleij and W.H.O. Ernst, 2001. Enhanced ATP-dependent copper efflux across the root cell plasma membrane in copper tolerant Silene vulgaris. Physiol. Plant., 113: 225-232.
Direct Link - Volkmar, K.M., Y. Hu and H. Stephan, 1997. Physiological responses of plants to salinity: A review. Can. J. Plant Sci., 78: 19-27.
Direct Link - Zhong, H. and A. Lauchli, 1994. Spacial distribution of solutes, K, Na, Ca and deposition rates in the growth zone of primary cotton roots: Effects of NaCl and CaCl2. Planta, 194: 34-41.
CrossRef