
Yahya S. Masrahi
Department of Biology, Faculty of Science, Jazan University, Jazan, Saudi Arabia
LiveDNA: 966.33187
Asian Journal of Plant Sciences
Year: 2020 | Volume: 19 | Issue: 4 | Page No.: 443-454







ABSTRACT
Background and Objective: Urochondra setulosa (Trin.) C.E. Hubb. is a perennial halophytic grass of Poaceae, widely distributed in coastal areas from Eastern Africa, through Arabia to NW India. This species shows high adaptation with drought and salinity stresses. Therefore, the objective of this study was to determine the micromorphological and anatomical features that responsible for it’s high adaptability to harsh conditions. Materials and Methods: Leaf micromorphology and anatomy were studied by LM, SEM whereas elemental analysis was determined by energy-dispersive X-ray spectroscopy (EDS). Dew collecting ability was observed in the field as well as in the laboratory by exposing leaves to the mist stream generated by a cold mist humidifier. Results: The adaxial leaf surface is partitioned longitudinally by ridges and grooves. Both surfaces have two types of trichomes, prickle hairs, and papillae, in which prickle hairs were more density. Silicon was detected in the cuticle with more percentage in the prickle hairs. Leaf surface structures have "wetness" traits that improve dew collecting ability. Salt glands were observed in the adaxial surface, with diurnal activity combined with wetness properties. The leaf has kranz anatomy with extensions of chlorenchymatous bundle sheath, indicating to C4 photosynthetic pathway with the NAD-ME subtype. Conclusion: Micromorphological and anatomical characteristics in the leaf seem to greatly enhance the adaptability of Urochondra setulosa and enable it to cope with drought and salinity conditions.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Yahya S. Masrahi, 2020. Ecological Adaptations of Urochondra setulosa (Poaceae) against Drought and Salinity. Asian Journal of Plant Sciences, 19: 443-454.
DOI: 10.3923/ajps.2020.443.454
URL: https://scialert.net/abstract/?doi=ajps.2020.443.454
DOI: 10.3923/ajps.2020.443.454
URL: https://scialert.net/abstract/?doi=ajps.2020.443.454
INTRODUCTION
Drought and Salinity stress are of the most limiting environmental factors affecting vegetation in arid and semiarid habitats. The existence of prominent diversity of plant species in these habitats suggests that such plants have an array of morpho-anatomical and physiological adaptations to endure drought and salinity stress1,2. Among vegetation units in arid and semiarid habitats, halophytes represent species that have a high resistance to both drought and salinity in their environment.
Supratidal zones along the coastal belt of Sabkhas and islands of Southern Red Sea are highly stressed habitats with high temperatures (average annual 28-30°C) and low precipitation rates (63-150 mm year–1). These conditions lead to potential evapotranspiration rates of 1800-3000 mm year–1, which are among the highest globally3. As evaporation exceeds precipitation, there is an enrichment of salts in the surface layers as these zones just above capillary fringe in which seawater rises by capillarity from the seawater table. Owing to high evaporation, the Red sea among the highest values in salinity4, so the concentration of the salts in the surface layers of soil in Sabkhas is very high. A combination of drought and high salinity in these habitats play a significant role in determining the distribution of halophytes. Furthermore, the functional limits of salt tolerance of each halophytic species are strongly correlated with their distribution pattern5.
Although there have been some studies dealt with integrated morpho-anatomical characters of halophytes and its relation to physiological mechanisms for survival in both drought and saline habitats2,6-8, there have been few studies in this context about grass leaf micromorphology and anatomy9-11.
Urochondra setulosa (Trin.) C.E. Hubb. is a perennial halophytic grass of Poaceae (subfamily Chloridoideae). This species grows in coastal habitats from Eastern Africa, through Arabia to NW India12,13 In Saudi Arabia, U. setulosa inhabit sabkhas of Red Sea coastal area and some islands of Western and Southwestern part of country13.
Previous studies showed that U. setulosa could survive 1000 mM NaCl, indicating that it is a very high salt tolerant14-16. These studies have focused on some physiological aspects of salt tolerance in terms of ionic content and seed germination. However, there have been no studies done on the micromorphological and anatomical aspects of U. setulosa in relation to salinity and drought tolerance. Morpho-anatomical features are important mechanisms for plant survival in harsh conditions, and with its linkage of physiological characters, these features represent crucial adaptation against drought and salinity. The knowledge of such adaptive mechanisms of halophytes, particularly those distributed in a high stressful habitat like southern Red Sea-type ecosystems, is therefore of increasing significance from both an agricultural and an ecological point of view.
The objective of this study was to assess some micromorphological and anatomical features of leaf in U. setulosa, as adaptive traits to salinity and drought that prevailing in its natural habitats.
MATERIALS AND METHODS
Study site: Al Ga'afari island lies 20 km SW of Jazan coast, 16°41'N, 42°33'E (Fig. 1). This island has a small area of ~0.7 km2 , with a sandy beach and sandy-loam sabkha inland. The climate, as a part of Jazan city (nearest climatic station), characterized by a high temperature of 30°C as an annual mean and low precipitation rate of ~150 mm year–1, with high humidity of >60% most of the year months. The rainfall season is mainly in the summer months. Vegetation is sparse with patches of halophytes, dominated by Salvadora persica, Suaeda vermiculata, Zygophyllum album, Limonium axillare and Urochondra setulosa.
Field observations and sample collection: Three field trips were conducted in the period of 1-5 April 2018. Field observations were performed in the course of all day to monitor diurnal patterns of salt crystals deposited on the adaxial leaf surfaces, as well as dew precipitated that observed in the early morning. Leaf samples were collected and dividing into two sets, one stored in 70% ethanol to subsequent examination by light microscopy (LM), scanning electron microscopy (SEM) and energy-dispersive X-ray spectroscopy (EDS), whereas other transfer freshly to the lab for dew collecting experiment.
LM, SEM microscopy and EDS analysis: Transverse sections of leaves were cut with a sharp razor. The sections were then stained with neutral red and examined under a light microscope. Pieces of the leaf (adaxial surfaces) were examined with SEM (JSM-6380 LA-JEOL, Japan). Dried pieces of leaves (3-5 mm) were mounted on the stub on double side carbon tap, sputter-coated with gold and examined under high vacuum with an accelerating voltage of 10 kV.
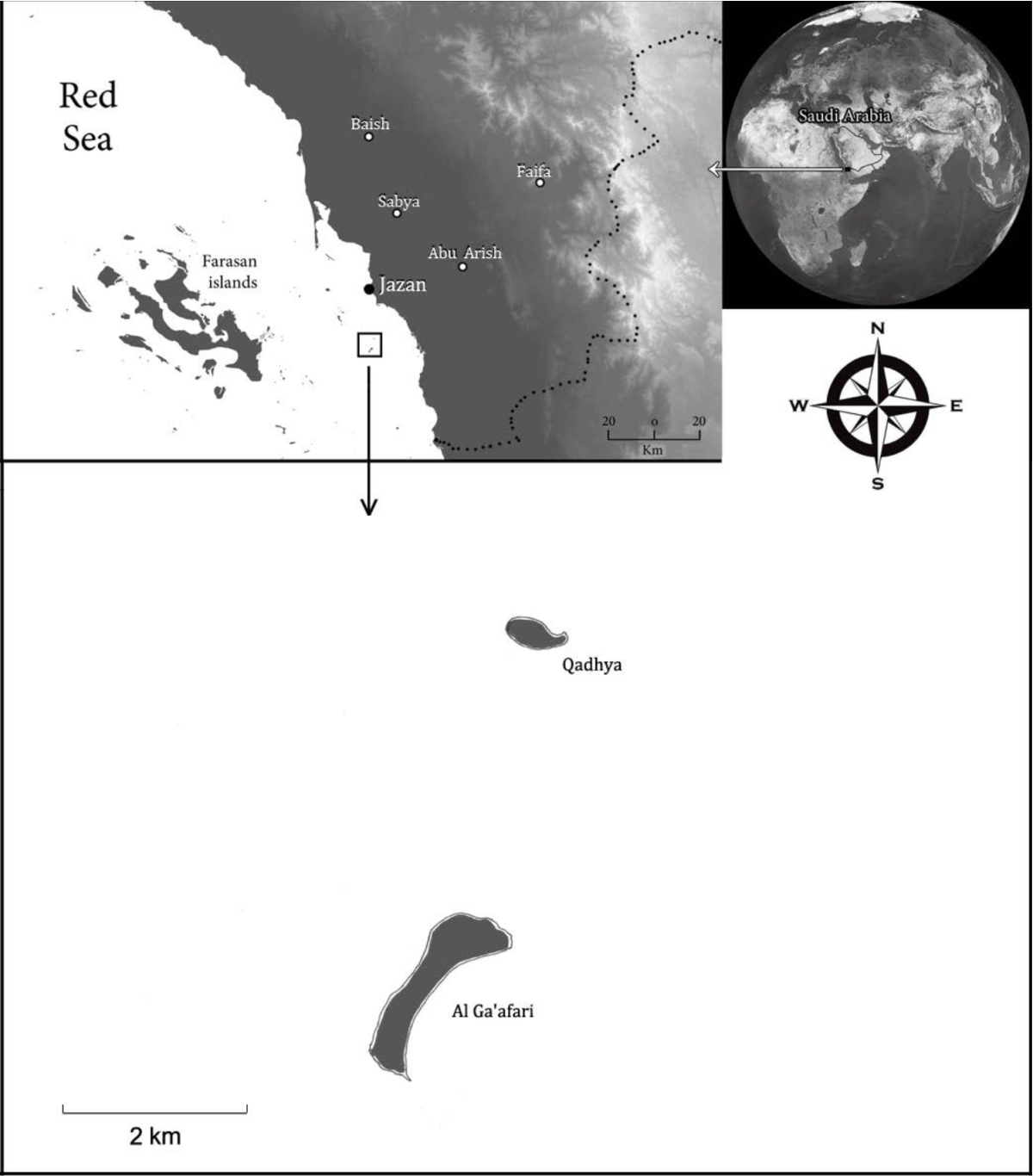 |
| Fig. 1: | Study site, Al Ga'afari island, in which Urochondra setulosa represents one of the dominant species |
Elemental analysis of the leaf epidermal surface was determined by energy-dispersive X-ray spectroscopy (EDS) (JSM-6380 LA-JEOL, Japan).
Dew collecting experiment: Dew collecting ability of leaf surfaces was observed in the field. To study this ability in the lab, fresh and clean leaves were chosen to put in front of the purifying mist stream generated by a cold mist humidifier (BLACK+DECKER HM3000), at a distance of 15 cm from the mist outlet. The flow of the mist was adjusted by control dial to a low flow rate of ~1 mL h–1. The speed flow of the mist stream was ~ 0.7 m sec–1. This low flow rate was chosen to mimic normal conditions in dewy nights, and then reveal the efficiency of dew collecting with more precisely. Dew collection ability was performed for 1 min. and immediately imaging by digital camera adjusted to a stereomicroscope (SONY FD Mavica 2.0 MP).
RESULTS AND DISCUSSION
Micromorphology of leaf surface: Figure 2a reveals the general habit of plants and leaves. Enlarged adaxial leaf surface shows a prominent microstructure comparing with the abaxial surface (Fig. 2b-e). The surface of U. setulosa leaf, as many grasses, divided into longitudinal zones of ridges and grooves between them, causing the leaf surface to appear undulating. Two types of trichomes are distributed on both surfaces, prickle hairs and papillae. While papillae did not show notable differences, prickle hairs were more density on the adaxial leaf surface.
 |
| Fig. 2(a-e): | (a) Urochondra setulosa habit, (b) Adaxial leaf surface, This image was taken at 6 pm, Note salt crystals on the surface, (c) SEM of leaf surface showing ridge (R) and grooves (G), (d) Transverse section in leaf. Extensions of chlorenchymatous bundle sheath are indicated by arrowheads, Note prickle hair (pr),vascular bundle (vb), bulliform cell (bc), ridge (R) and grooves (G), (e) Enlargement of square area in c, Note papilla (pa) and prickle hair (pr) |
It was observed from SEM analysis that, Epicuticular wax not thick, covering nearly all surfaces of the epidermis (Fig. 3), except prickle hair tips. According to the classification of the epicuticular waxes proposed by Barthlott et al.17, U. setulosa showed the crystalloid type of platelets. This pattern of wax protruding from the pervasive wax film.
Stomata are not observed clearly in the images of LM or SEM. It is known that stomata in subfamily Chloridoideae are generally small with narrow, oblong slit that difficult to seen in the presence of other epidermal features (epicuticular waxes, trichomes),as well as its location in grooves or close to it18,19.
Elemental composition analysis of the surface material of epidermis (cuticle and wax) indicated that the silicon (Si) was the third main element (after C,O), deposited with more percentage on the tips of prickle hairs than on the other areas of the epidermis (Fig. 4, 5).
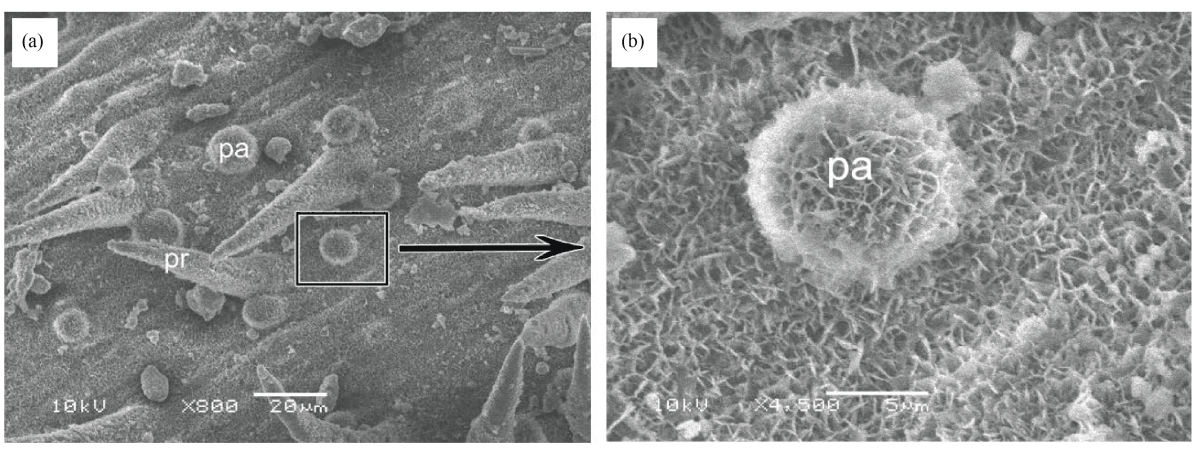 |
| Fig. 3(a-b): | (a) SEM of a part of adaxial leaf surface, Note papilla (pa) and prickle hair (pr) and (b) Enlargement of square area in a, showing epicuticular wax depositions, with platelets manner |
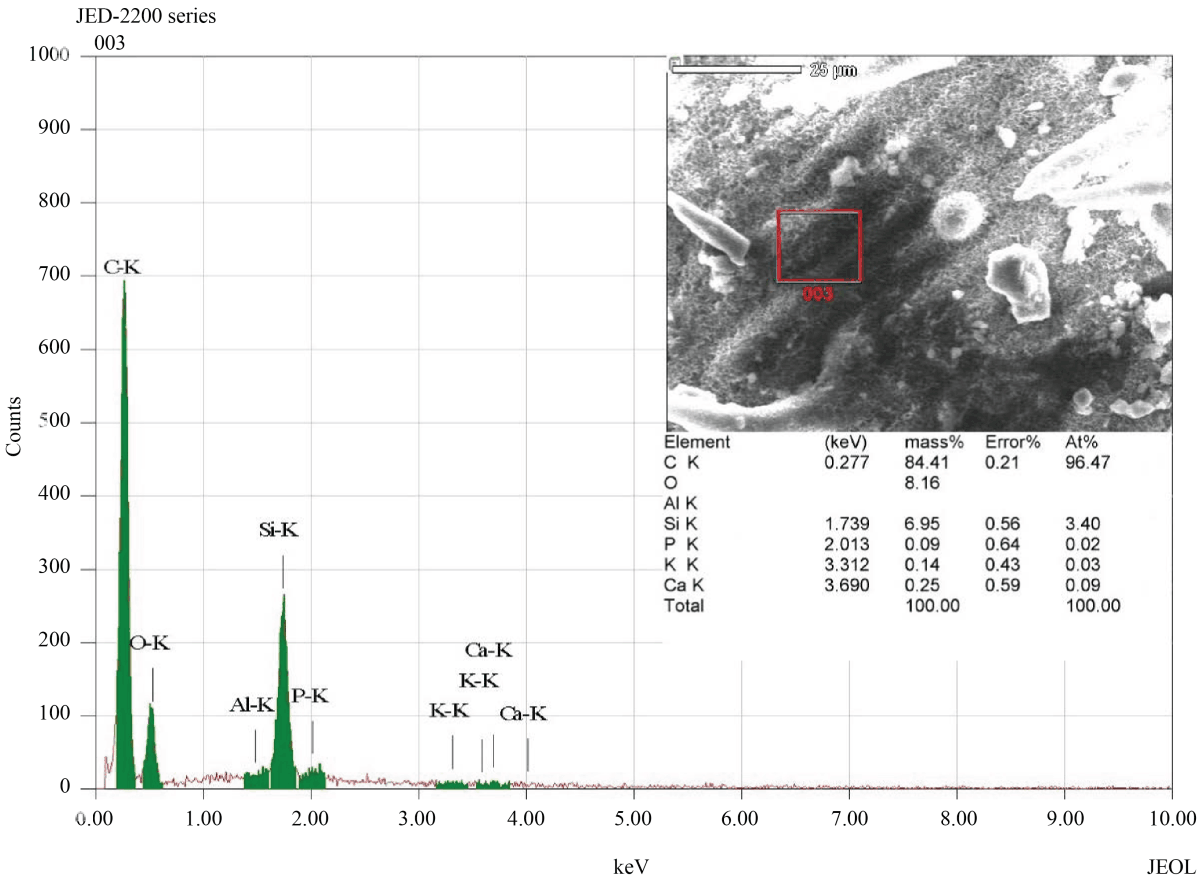 |
| Fig. 4: | The elemental composition of leaf surface between trichomes, The red square in the inset image shows the selected EDS examination field. The resulting spectrum reveals that the silicon (Si) is the third main element, on the basis of mass percentage, present in the selected field (after C,O) |
Dew condensation and salt deposition: In the early morning (5-7 am), drops of dew were observed on the leaf surface of U. setulosa in its natural habitat (Fig. 6a, b). Although dew condensation was observed also on the other leaf and plant surfaces of associated species in the habitat, dew condensation was prominent and dense on the leaves of U. setulosa. After few hours from the early morning, temperature increases resulting in evaporation of dewdrops, leaving salt crystals that appear in a full dry state at the end of the day (4-6 pm) (Fig. 2b).
To assess the role of leaf surface structures in dew collection ability, a dew collecting experiment was done in the laboratory, as described above. After 1 min of exposed to mist stream of cold mist humidifier, leaf surface located in front of the mist stream shows deposited droplets on the surface.
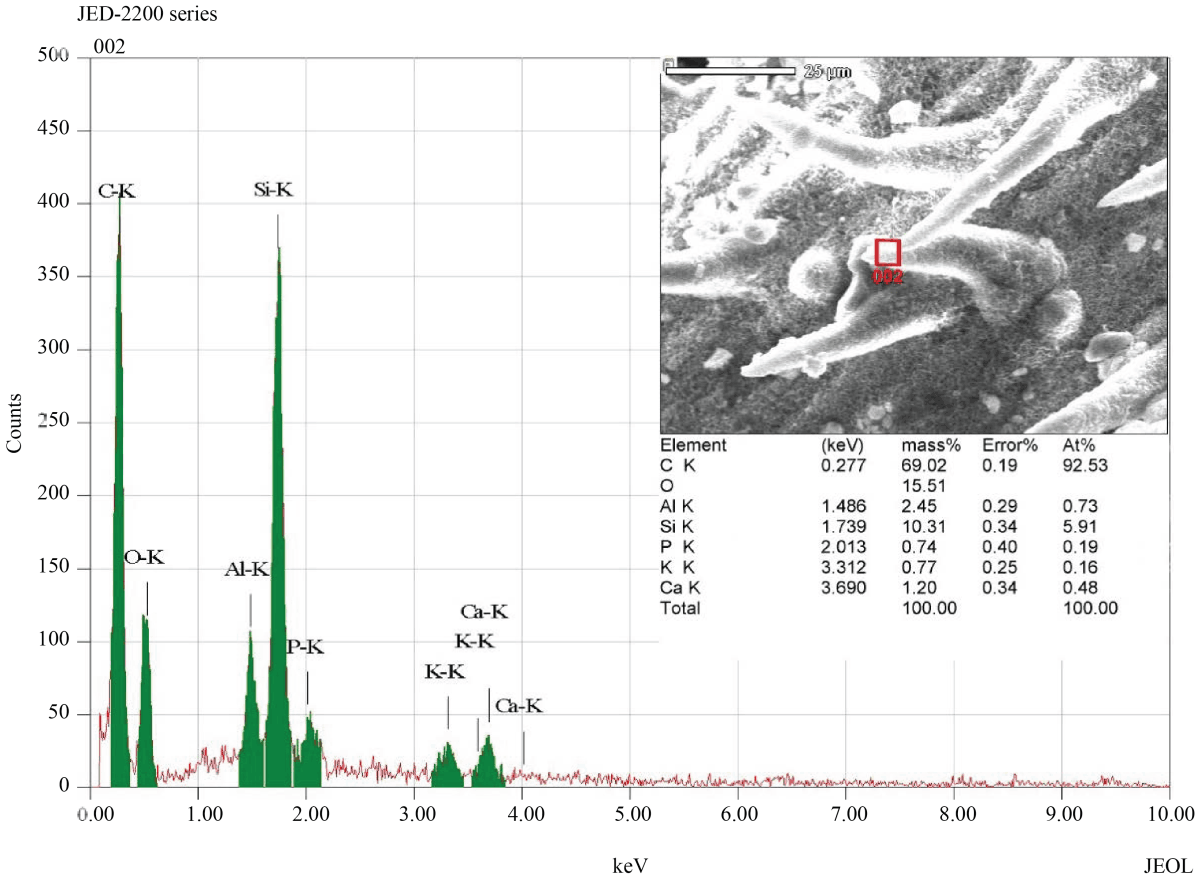 |
| Fig. 5: | The elemental composition of prickle hairs, The red square in the inset image shows the selected EDS examination field. The resulting spectrum reveals that the silicon (Si) is the third main element, on the basis of mass percentage, present in the selected field (after C,O) |
Droplets appeared to start at the tips of prickle hairs, growing gradually to bigger droplets, coalesce together to make observed drops (Fig. 6c, d). Dew collection ability was observed on both surfaces of the leaf.
Leaf anatomy: Figures 2d, 7, 8 reveal transverse sections in the leaf. Ridges have a high density of prickle hairs comparing with other areas. Adaxial epidermal cells of ridges are thick-walled with a thick layer of fibers beneath them, making a sclerenchymatous cap of the ridges (Fig. 7a, b, d). Abaxial ridges also showed sclerenchymatous cap but less in thickening. Beneath the sclerenchymatous cap, large colorless of parenchyma cells can be seen (Fig. 7b, d), that differ from surrounding chlorenchymatous cells in that they are larger and lack chloroplasts. Bulliform cells are found on the adaxial surface in the base of grooves (Fig. 2d, 7a). In lack of turgor pressure due to water stress, these cells are responsible for leaf rolling (Fig. 7c).
Salt glands are located in the flanks of grooves area in the adaxial surface, whereas not observed in the abaxial surface (Fig. 8a). Salt glands are bicellular, consisting of flask-shaped basal cells and globular cap cells (Fig. 8b).
U. setulosa leaves have kranz type anatomy, indicating to C4 photosynthetic pathway. Kranz anatomy here has extensions of chlorenchymatous bundle sheath from the vascular bundle to the patches of mesophyll cells (Fig. 2d, 7a). Plant distribution depends on climatic and edaphic conditions, as well as structural and functional features enabling plants to cope with these conditions19,20. Sabkhas of southern Red Sea coasts and islands have highly stressful conditions of drought and salinity. It is well known that drought and salinity have tightly related effects to which plants exhibit many morphoanatomical and physiological adaptations7,21-23. Although morphoanatomical features are common among many halophytic species, each species has its distinctive array of features20.
Leaf surface of U. setulosa has invaginations with distinct ridges and deep grooves which are known to prevent water loss by lowering leaf conductance in many grasses11. These ridges fit together upon leaf rolling decreasing adaxial surface area and lowering the transpiration rate. Bulliform cells were reported on the adaxial surface at the base of grooves. Bulliform cells are large water containing thin-walled cells characteristic of many grasses2,8,24. These highly vacuolated cells lose water under dry conditions causing inward leaf rolling where ridges fit together25.
 |
| Fig. 6(a-d): | (a and b) Images of Urochondra setulosa leaves in its natural habitat, This image was taken at 6 am Note the dew depositions on the leaf surface, (c) Leaf surface before and (d) After exposing to mist stream generated by a mist humidifier Note starting deposition of a droplet (No. 1) and the gradually growing (coalesce with each other) to become a large droplet (No. 2, 3) |
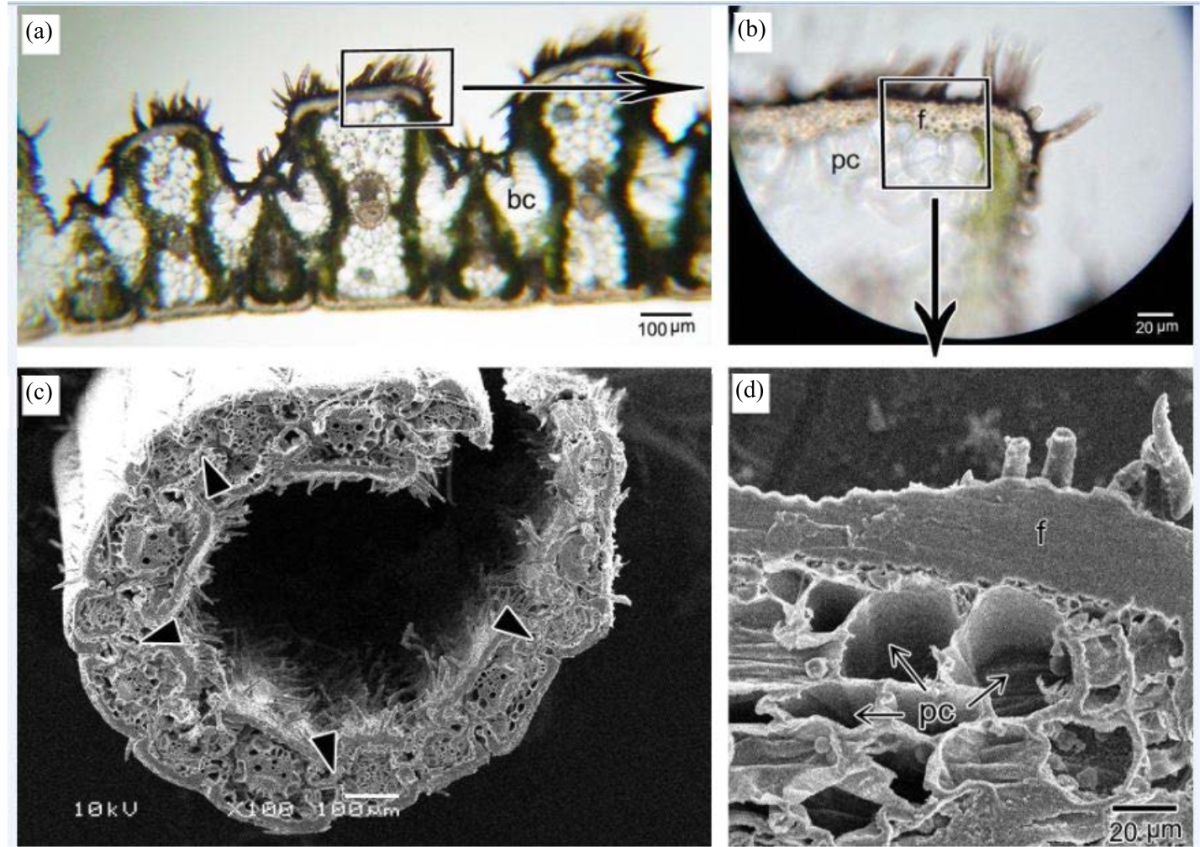 |
| Fig. 7(a-d): | (a) Transverse section in leaf. Note bulliform cell (bc), (b) Enlargement of rectangle area in A, Note fibers (f) and parenchyma cells (pc), (c) SEM of transverse section in leaf showing rolling as a result to the lack in turgor pressure of bulliform cells (arrowheads), (d) Enlargement of square area in B. Note the thickness of fibers layer (f) and parenchyma cells (pc) beneath them |
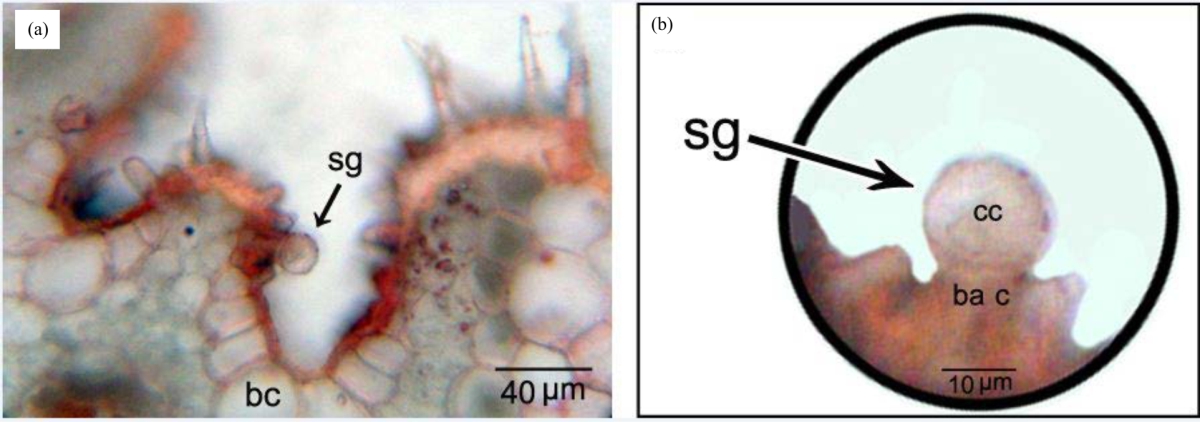 |
| Fig. 8: | (a) Salt gland (sg) in the flank of groove area in the adaxial surface. Note bulliform cell (bc), (b) Salt gland (sg) comprising basal cell (ba c) and globular cap cell (cc) |
The ridge itself has a thick sclerenchymatous cap of fibers which enhances mechanical strength, reduces water loss, and provides protection against high solar radiation.
Moreover, U. setulosa leaf trichomes found on both surfaces are prickle hairs and papillae. Prickle hairs represent the most prominent microstructures on both leaf surfaces. The presence and density of hairs on the leaf surface is a characteristic feature of plants in hot arid habitats26,27. The dense hairs on the leaf surface substantially increase the reflectance of solar radiation resulting in reduced heat load, lower leaf temperature and reduced transpiration rate26,27. These hairs also enhance water use efficiency and lower photoinhibition28,29. This type of trichomes effectively increases leaf boundary layer thickness and decrease the transpiration rate27,29,30. In addition, the numerous papillae on both leaf surfaces restrict airflow in the grooves which further helps water conservation11. Furthermore, epicuticular wax of U. setulosa showed the crystalloid type of platelets. Despite the low thickness of epicuticular wax, it plays a significant role in decreasing water loss due to its hydrophobic nature and increases the reflection of incident solar radiation due to its 3-dimensional shape31.
Prickle hairs tips are not covered by wax, making the tips hydrophilic, enhancing wettability of these surfaces32. Under favorable conditions, water molecules collect as very small droplets on the tip of cone-like prickle hairs33. These features collectively contribute to the dew collection ability of U. setulosa observed in dewy nights in the field and in laboratory experiments. Condensation of dew on leaf surface creates moist air next to leaf surface leading to higher vapor pressure which stimulates greater stomatal opening and CO2 uptake with low transpiration rate34-36. These characteristics greatly enhance water use efficiency. After starting at prickle hairs tips, water droplets gradually grow to bigger droplets and coalesce to make observed drops and water patches where gravity then drives water in grooves along leaf axis (Fig. 6a, b). Small droplets can also evaporate quickly due to higher surface to volume ratio, whereas larger droplets may continue for several hours37. Further, dew condensation on leaf surfaces dissolves precipitated salt crystals observed in the early morning in the field, resulting in "salty drops" on leaf surfaces. When dissolved in the water, salt will cause a reduction in the vapor pressure at the water surface, decreasing the evaporation rate38, resulting also to more persistence of drops on the leaf surfaces.
The salt glands in the leaf of U. setulosa were observed in the flanks of grooves area in the adaxial surface. This location gives protection to the salt glands, which has been observed also in other Chloridoideae39,40. Salt glands were bicellular, consisting of flask-shaped basal cell and globular cap cell, as in most halophytic grasses39,41. The basal cell is directly in contact with colorless parenchyma cells underneath. In halophytes, this parenchyma represents water-storing cells that accumulate salt ions in their vacuoles (mainly as NaCl) against a diffusion gradient42,43. The basal cell act as a "collecting cell" to ions from the parenchyma cells, and transfer them to cap cell which acts as "secreting cell" 39.
Diurnal pattern of salt crystals deposited on the adaxial leaf surface that observed in the end of the day and dew condensation that observed in the early morning, indicate to the activity of salt glands that permanently excrete salt ions. The activity of salt glands was found to be greatest in high saline habitats41,44. The development of hydrostatic pressure due to the accumulation of salt ions in salt glands might serve to remove tiny salt crystals44, as well as removing by dissolving in dew water in the dewy nights. Salt crystals are hygroscopic and become deliquescent at higher humidity, increasing leaf surface wetness45 even in non-dewy nights at prevailing high humidity. These conditions in the natural habitat of U. setulosa enhance "wetness" of leaf surface and activity of salt secretion by salt glands. Although halophytes require some salt ions for optimal growth (typically Na and Cl) in levels higher than those found in non-saline soils, high concentrations of these ions have toxic effects on a metabolic process like chloroplast function and ion homeostasis46,47. Salt glands, with its diurnal rhythm of activity, combined with surface wetness features of leaf, represent an efficient mechanism to salt excretion and maintain ion homeostasis.
Silicon (Si) represents the second most abundant element, after oxygen, in the earth soils48. Plant roots absorb silicon from the soil in the form of silicic acid49,50 and deposited as hydrated amorphous silica51,52. Silicon is known to enhance plant protection from multiple abiotic and biotic stresses, including salinity, physical damage, radiation, drought, high temperature, herbivores and diseases53. Although most plants do not accumulate silicon perceptibly, grasses (Poaceae) one of the few groups that accumulate substantially more silicon in their tissues53,54. Leaf epidermis and its appendages are one of the most intensely silicified parts in grasses50, in which silicon accumulates into, and on, the cell wall matrix55, forming silica-cuticle double layer56. This accumulation of silicon onto the cell wall matrix is comparable to the role of lignin in lignified walls52. Comparing to lignification, the energy costs of silicon deposition were estimated to be 20-times lesser57, which is, metabolically, low-cost solving for getting mechanical rigidity. Silicon in the Surface materials of the epidermis (leaf Cuticle) of U. setulosa was the third main element (after C,O) on the basis of mass percentage (~7%), whereas in prickle hairs were more (~10%). These percentages of silicon in U. setulosa leaf lay in the maximum range known in Plants, which vary between 0.1 to 10% or more48,52. The deposition of silicon in cuticle enhances mechanical strength and rigidity greatly decrease water loss through transpiration and thus reduce drought stress49,53,56,58. Furthermore, silicon was densely found in prickle hairs especially in the point-tips. This manner of silicon deposition reinforces rigidity and sharpness of the prickle hairs, enhancing its function as dew collectors, in which water molecules tend to captured as a very small droplet on the tip of the tapered prickle hair. Also, the rigidity of prickle hairs will enhance other known functions of this type of trichomes, like increasing in the thickness of the leaf boundary layer and reflectance of solar radiation.
U. setulosa leaves have kranz anatomy, indicating to C4 photosynthetic pathway. Kranz anatomy here have extensions of chlorenchymatous bundle sheath from the vascular bundle to the mesophyll cells. This form of Kranz anatomy belongs to the subtype NAD-ME of C4 photosynthetic pathway59,60, in which the decarboxylation enzyme is NAD-malic. Koteyeva et al.61 confirmed also this subtype in U. setulosa biochemically by enzyme assay. Plants with C4-photosynthetic pathway predominantly inhabit hot and arid environments, because this pathway includes mechanisms tolerate, and adapted to, high temperature and low soil water conditions62, resulting in high water use efficiency. Furthermore, along with all subtypes of C4 photosynthesis, NAD-ME characterizes species that inhabit drier and saline environments63-65. The high water use efficiency of C4 plants (especially with NAD-ME subtype) represents an advantage under salinity stress because the higher water use efficiency decreasing the amount of salt ions that plant must excrete63. This feature lowers the metabolic cost as the salt secretion is an energy consumptive process46.
CONCLUSION
On the basis of present study results, it can be concluded that U. setulosa is a remarkable halophytic grass with many micromorphological and anatomical features of leaves that collectively enhance cope with extreme conditions of both drought and salinity in its natural habitat. Micromorphological characteristics including ridges and deep grooves with dense trichomes of prickle hairs and papillae. These traits lead to the reflectance of solar radiation, reduce heat load, increase the boundary layer and decrease transpiration. Prickle hairs also enhance "wetness" of the leaf surface and, with the activity of salt glands, improve salt excretion, dew collection and retain it ability. Silicon, which impregnated with cuticle, enhances mechanical strength and rigidity of the epidermal structures. Adaxial leaf surfaces have bulliform cells that play a substantial role in leaf rolling and reopen in response to water stress conditions. Anatomically, U. setulosa have kranz type with extensions of chlorenchymatous bundle sheath, indicated to C4 photosynthetic pathway, with NAD-ME subtype. These features of U. setulosa leaf greatly enhance adaptation with drought and high salinity, leading to higher water use efficiency in high stressful habitat.
SIGNIFICANCE STATEMENTS
This study revealed that U. setulosa leaves have prominent morpho-anatomical characteristics. Previously researchers concentrated on some physiological features of this species, especially salt resistance, whereas micromorphology and anatomy of leaves still unknown properly. The results of this study suggested that micromorphological and anatomical traits of U. setulosa leaf enhance, collectively, adaptation against drought and high salinity, explaining many physiological features reported formerly.
REFERENCES
- Bohnert, H.J., D.E. Nelson and R.G. Jensen, 1995. Adaptations to environmental stresses. Plant Cell., 7: 1099-1111.
CrossRefPubMedDirect Link - Grigore, M.N., L. Ivanescu and C. Toma, 2014. Halophytes: An Integrative Anatomical Study. Springer International Publishing. Cham, Switzerland.
CrossRefDirect Link - Le Houerou, H.N., 2003. Bioclimatology and phytogeography of the red sea and aden gulf basins: a monograph (with a particular reference to the highland evergreen sclerophylls and lowland halophytes). Arid Land Res. Manag., 17: 177-256.
CrossRefDirect Link - McLachlan, A., 2018. Africa, coastal ecology. In: Encyclopedia of Coastal Science, Schwartz, M.L., Springer International Publishing, Cham, Switzerland, pp: 4-9.
CrossRefDirect Link - Ungar, I.A., 1998. Are biotic factors significant in influencing the distribution of halophytes in saline habitats? Bot. Rev., 64: 176-199.
CrossRefDirect Link - Tattini, M., D. Remorini, P. Pinelli, G. Agati, E. Saracini, M.L. Traversi and R. Massai, 2006. Morpho-anatomical, physiological and biochemical adjustments in response to root zone salinity stress and high solar radiation in two Mediterranean evergreen shrubs, Myrtus communis and Pistacia lentiscus. New Phytol., 170: 779-794.
CrossRefDirect Link - Grigore, M.N. and C. Toma, 2017. Anatomical Adaptations of Halophytes. Springer International Publishing, Cham, Switzerland.
CrossRefDirect Link - Abernethy, G.A., D.W. Fountain and M.T. McManus, 1998. Observations on the leaf anatomy of Festuca novae‐zelandiaeand biochemical responses to a water deficit. New Zealand J. Bot., 36: 113-123.
CrossRefDirect Link - Gielwanowska, I., E. Szczuka, J. Bednara and R. Górecki, 2005. Anatomical features and ultrastructure of Deschampsia antarctica (poaceae) leaves from different growing habitats. Ann. Bot., 96: 1109-1119.
CrossRefDirect Link - Maricle, B.R., N.K. Koteyeva, E.V. Voznesenskaya, J.R. Thomasson and G.E. Edwards, 2009. Diversity in leaf anatomy, and stomatal distribution and conductance, between salt marsh and freshwater species in the C4 genus Spartina (Poaceae). New Phytol., 184: 216-233.
CrossRefDirect Link - Gandhi, D. and S. Albert, 2015. Morphometric variations in caryopses and seedlings of two grass species growing under contrasting habitats. Not. Sci. Biol., 7: 355-360.
CrossRefDirect Link - Ghazanfar, S.A., 1998. Vegetation of the plains. In: Vegetation of the Arabian Peninsula, Ghazanfar,S.A. and M. Fisher, Springer, Netherlands pp:175-190.
CrossRefDirect Link - Gulzar, S., M.A. Khan and L.A. Ungar, 2003. Salt tolerance of a coastal salt marsh grass. Commun. Soil Sci. Plant Anal., 34: 2595-2605.
CrossRefDirect Link - Shaikh, F., B. Gul, W.Q. Li, X.J. Liu and M.A. Khan, 2007. Effect of calcium and light on the germination of Urochondra setulosa under different salts. J. Zhejiang Univ. Sci. B, 8: 20-26.
CrossRefPubMedDirect Link - Gulzar, S. and M.A. Khan, 2006. Comparative salt tolerance of perennial grasses. In: Ecophysiology of High Salinity Tolerant Plants, Khan, M.A. and D.J. Weber, Springer, Netherlands, pp: 239-253.
CrossRefDirect Link - Barthlott, W., C. Neinhuis, D. Cutler, F. Ditsch, I. Meusel, I. Theisen and H. Wilhelmi, 1998. Classification and terminology of plant epicuticular waxes. Bot. J. Linn. Soc., 126: 237-260.
CrossRefDirect Link - Liu, H. and C.P. Osborne, 2014. Water relations traits of C4 grasses depend on phylogenetic lineage, photosynthetic pathway, and habitat water availability. J. Exp. Bot., 66: 761-773.
CrossRefDirect Link - Gauhl, E., 1976. Photosynthetic response to varying light intensity in ecotypes of Solanum dulcamara L. from shaded and exposed habitats. Oecologia, 22: 275-286.
CrossRefDirect Link - Batanouny, K.H., 2013. Plants in the Deserts of the Middle East. Springer, Berlin, Heidelberg.
CrossRefDirect Link - Smith, S.D., R.K. Monson and J.E. Anderson, 1997. Physiological Ecology of North American Desert Plants. Springer, Berlin, Heidelberg.
CrossRefDirect Link - Dajic, Z., 2006. Salt stress. In: Physiology and Molecular Biology of Stress Tolerance in Plants, Rao, K.V.M., A.S. Raghavendra and K.J. Reddy, Springer, Netherlands, pp: 41-99.
CrossRefDirect Link - Singh, M., J. Kumar, S. Singh, V.P. Singh and S.M. Prasad, 2015. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Bio/technol., 14: 407-426.
CrossRefDirect Link - Alvarez, J.M., J.F. Rocha and S.R. Machado, 2008. Bulliform cells in Loudetiopsis chrysothrix (Nees) Conert and Tristachya leiostachya Nees (Poaceae): structure in relation to function. Braz. Arch. Biol. Technol., 51: 113-119.
CrossRefDirect Link - Xu, P., A. Ali, B. Han and X. Wu, 2018. Current advances in molecular basis and mechanisms regulating leaf morphology in rice. Front. Plant Sci.
CrossRefDirect Link - Ehleringer, J.R. and H.A. Mooney, 1978. Leaf hairs, effects on physiological activity and adaptive value to a desert shrub. Oecologia, 37: 183-200.
CrossRefDirect Link - Lin, H., Y. Chen, H. Zhang, P. Fu and Z. Fan, 2017. Stronger cooling effects of transpiration and leaf physical traits of plants from a hot dry habitat than from a hot wet habitat. Funct. Ecol., 31: 2202-2211.
CrossRefDirect Link - Ripley, B.S., N.W. Pammenter and V.R. Smith, 1999. Function of leaf hairs revisited: the hair layer on leaves Arctotheca populifolia reduces photoinhibition, but leads to higher leaf temperatures caused by lower transpiration rates. J. Plant Physiol., 155: 78-85.
CrossRefDirect Link - Bickford, C.P., 2016. Ecophysiology of leaf trichomes. Funct. Plant Biol., 43: 807-814.
CrossRefDirect Link - Johnson, H.B., 1975. Plant pubescence: An ecological perspective. Bot. Rev., 41: 233-258.
CrossRefDirect Link - Koch, K., B. Bhushan and W. Barthlott, 2010. Multifunctional plant surfaces and smart materials. In: Springer Handbook of Nanotechnology, Bhushan, B., Springer, Berlin, Heidelberg, pp:1399-1436.
CrossRefDirect Link - Cao, M., J. Ju, K. Li, S. Dou, K. Liu and L. Jiang, 2014. Facile and large-scale fabrication of a cactus-inspired continuous fog collector. Adv. Funct. Mater., 24: 3235-3240.
CrossRefDirect Link - Smith, W.K. and T.M. McClean, 1989. Adaptive relationship between leaf water repellency,stomatal distribution,and gas exchange. Am. J. Bot., 76: 465-469.
CrossRefDirect Link - Brewer, C.A., W.K. Smith and T.C. Vogelmann, 1991. Functional interaction between leaf trichomes, leaf wettability and the optical properties of water droplets. Plant Cell Environ., 14: 955-962.
CrossRefDirect Link - Monteith, J.I., 1995. A reinterpretation of stomatal responses to humidity. Plant Cell Environ., 18: 357-364.
CrossRefDirect Link - Brain, P. and D.R. Butler, 1985. A model of drop size distribution for a system with evaporation. Plant Cell Environ., 8: 247-252.
CrossRefDirect Link - Al-Shammiri, M., 2002. Evaporation rate as a function of water salinity. Desalination, 150: 189-203.
CrossRefDirect Link - Liphschitz, N. and Y. Waisel, 1974. Existence of salt glands in various genera of the gramineae. New Phytol., 73: 507-513.
CrossRef - Somaru, R., Y. Naidoo and G. Naidoo, 2002. Morphology and ultrastructure of the leaf salt glands of Odyssea paucinervis (Stapf) (Poaceae). Flora - Morphol., Distribution, Funct. Ecol. Plants, 197: 67-75.
CrossRefDirect Link - Céccoli, G., J. Ramos, V. Pilatti, I. Dellaferrera, J.C. Tivano, E. Taleisnik and A.C. Vegetti, 2015. Salt glands in the poaceae family and their relationship to salinity tolerance. Bot. Rev., 81: 162-178.
CrossRefDirect Link - Dassanayake, M. and J.C. Larkin, 2017. Making plants break a sweat: the structure, function, and evolution of plant salt glands. Front. Plant Sci.
CrossRefDirect Link - Semenova, G.A., I.R. Fomina and K.Y. Biel, 2010. Structural features of the salt glands of the leaf of Distichlis spicata Yensen 4a (Poaceae). Protoplasma, 240: 75-82.
CrossRefDirect Link - Hansen, D.J., P. Dayanandan, P.B. Kaufman and J.D. Brotherson, 1976. Ecological adaptations of salt marsh grass Distichlis spicata (Gramineae) and environmental factors affecting its growth and distribution. Am. J. Bot., 63: 635-650.
CrossRefDirect Link - Burkhardt, J., M. Hunsche, 2013. “Breath figures” on leaf surfaces—formation and effects of microscopic leaf wetness. Front. Plant Sci.
CrossRefDirect Link - Hasegawa, P.M., R.A. Bressan, J.K. Zhu and H.J. Bohnert, 2000. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol., 51: 463-499.
CrossRefDirect Link - Parida, A.K. and A.B. Das, 2005. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf., 60: 324-349.
CrossRefPubMedDirect Link - Epstein, E., 1994. The anomaly of silicon in plant biology. Proc. Nat. Acad. Sci. USA, 91: 11-17.
CrossRefDirect Link - Cooke, J. and M.R. Leishman, 2011. Is plant ecology more siliceous than we realise? Trends Plant Sci., 16: 61-68.
CrossRefDirect Link - Kumar, S., M. Soukup and R. Elbaum, 2017. Silicification in grasses: variation between different cell types. Front. Plant Sci.
CrossRefDirect Link - O'Reagain, P.J. and M.T. Mentis, 1989. Leaf silicification in grasses — A review. J. Grassland Soc. Southern Africa, 6: 37-43.
CrossRefDirect Link - Albersheim,P., A. Darvill, K. Roberts, R. Sederoff and A. Staehelin 2011. Plant Cell Walls. Garland Science, New York,.
CrossRefDirect Link - Ma, J.F. and N. Yamaji, 2006. Silicon uptake and accumulation in higher plants. Trends Plant Sci., 11: 392-397.
CrossRefDirect Link - Motomura, H., T. Fujii and M. Suzuki, 2006. Silica deposition in abaxial epidermis before the opening of leaf blades of Pleioblastus chino (poaceae, bambusoideae). Ann. Bot., 97: 513-519.
CrossRefDirect Link - Yan, G.C., M. Nikolic, M.J. Ye, Z.X. Xiao and Y.C. Liang, 2018. Silicon acquisition and accumulation in plant and its significance for agriculture. J. Integr. Agric., 17: 2138-2150.
CrossRefDirect Link - Ma, J.F., 2004. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr., 50: 11-18.
CrossRefDirect Link - Raven, J.A., 1983. The transport and function of silicon in plants. Biol. Rev., 58: 179-207.
CrossRefDirect Link - Kido, N., R. Yokoyama, T. Yamamoto, J. Furukawa, H. Iwai, S. Satoh and K. Nishitani, 2015. The matrix polysaccharide (1;3,1;4)-β-d-glucan is involved in silicon-dependent strengthening of rice cell wall. Plant Cell Physiol., 56: 268-276.
CrossRefDirect Link - Dengler, N., 1994. Quantitative leaf anatomy of C3 and C4 grasses (poaceae): bundle sheath and mesophyll surface area relationships. Ann. Bot., 73: 241-255.
CrossRefDirect Link - Koteyeva, N.K., E.V. Voznesenskaya and G.E. Edwards, 2015. An assessment of the capacity for phosphoenolpyruvate carboxykinase to contribute to C4 photosynthesis. Plant Sci., 235: 70-80.
CrossRefDirect Link - Taub, D.R., 2000. Climate and the U.S. distribution of C4 grass subfamilies and decarboxylation variants of C4 photosynthesis. Am. J. Bot., 87: 1211-1215.
CrossRefDirect Link - Cabido, M., E. Pons, J.J. Cantero, J.P. Lewis and A. Anton, 2007. Photosynthetic pathway variation among C4 grasses along a precipitation gradient in Argentina. J. Biogeography.
CrossRefDirect Link
حمید هویزهHamid hoveizeh Reply
Hi
I am from Iran.
I saw this plant in south costal iran.
I am so glad if you refer me complet paper.
scincerly.
Hamid hoveizeh