
Wanraya Suthumchai
Not Available
Toshiyuki Matsui
Not Available
Kazuhide Kawada
Not Available
Yusuke Kosugi
Not Available
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 4 | Page No.: 568-576







ABSTRACT
This study investigated the changes in activities of sugar metabolizing enzymes (acid invertase, sucrose synthase (SS) and sucrose phosphate synthase (SPS)) and sugar contents during storage at low temperature in lettuce head (cvs. ‘Shizuka’ and ‘Shisuko’). The acid invertase activity decreased while the SS activity showed a continuous increase during storage. In both cultivars and portions, sugar contents declined after 2 days of storage especially sucrose content. The SS activity and sugar contents had a significant negative correlation in both portions of ‘Shisuko’. However, a significant positive correlation was observed between acid invertase activity (soluble fraction) and sugar contents in the leaf portion of both cultivars. SPS activity did not show specific inclining or declining pattern throughout the storage time. Among the two cultivars, ‘Shizuka’ showed lower percentage of weight loss, degree of yellowing and respiration rate than ‘Shisuko’ which might be the reason for longer storage life of the latter.
PDF Abstract XML References Citation
How to cite this article
Wanraya Suthumchai, Toshiyuki Matsui, Kazuhide Kawada and Yusuke Kosugi, 2007. Sugar Metabolizing Enzymes Activities in Lettuce Head During Low Temperature Storage. Asian Journal of Plant Sciences, 6: 568-576.
DOI: 10.3923/ajps.2007.568.576
URL: https://scialert.net/abstract/?doi=ajps.2007.568.576
DOI: 10.3923/ajps.2007.568.576
URL: https://scialert.net/abstract/?doi=ajps.2007.568.576
INTRODUCTION
Vegetables are living tissues that their composition and physiology continue to change after harvest. Postharvest losses in fresh products is estimated 5 to 25% in developed countries and 20 to 50% in developing countries, depending upon the commodity, cultivar and handling conditions (Kader, 2002). Their conditions and marketable life are influenced by such things as temperature, humidity, air composition and microorganisms (Thompson, 2003). However, the single most important factor that affects fresh product storage life is temperature, which influences enzymatic and other chemical reaction rates (Bolin and Huxsoll, 1991). Low temperature storage maintains quality after harvest and prolongs storage life of fresh products by reducing respiration rate, moisture loss and senescence, as well as reducing growth of spoilage microorganisms and therefore delays senescence. (Watada et al., 1996; Jobling, 2002). In addition, low temperature protects non-appearance quality attributes such as texture, nutrition, aroma and flavor (Paull, 1999). Maintaining freshness and quality during storage is mainly a thing of temperature control but the optimum temperatures for keeping quality of various products vary considerably (Nilsson, 2000). Fresh fruits and vegetables need low temperatures (0 to 13°C) and high relative humidity (80 to 95%) to lower respiration rate and to slow down metabolic and transpiration rates (Gast, 1991). For temporary storage, however, temperatures of 0 to 2°C and 90 to 95% RH are recommended. For marketing within 1 to 2 weeks, storage temperatures consistently below 5°C are generally adequate to maintain good quality (Cantwell and Kasmire, 2002).
Iceberg lettuce (Lactuca sativa) is a popular fresh salad vegetable in many countries. It is a well recognized source of minerals, vitamins and dietary fiber (Pirovani et al., 2005). Head lettuce is a perishable commodity that requires immediate refrigerated storage after harvest to prolong its shelf life (Rennie et al., 2001). Lettuce can be stored up to 2 to 3 weeks when held at optimum conditions of 0°C and 98-100% RH (Gast, 1991). Moreover, Stanley (1989) suggested that lettuce quality was retained longer at 5 than 10°C or ambient temperatures.
Storage temperature can influence compositions (vitamins, sugar contents and other nutrients) in many fruits and vegetables (Paull, 1999). For instance, lamb’s lettuce stored for 8 days at 0 to 30°C showed increasing degradation of sucrose, glucose and fructose with increasing storage temperatures (Enninghorst and Lippert, 2003). The decrease in sugar contents, especially sucrose, leads to quality deterioration and senescence of fresh products. Sucrose is the major sugar transported through the phloem from source tissues to sink tissues in higher plants. Sucrose is synthesized by sucrose phosphate synthase (SPS; UDP-glucose: D-fructose-6-phosphate-2-α-glucosyltransferase; EC 2.4.1.14), a soluble enzyme located in the cytoplasm. SPS plays a key role in the synthesis of sucrose, which catalyzes the formation of sucrose-6-phosphate from uridine 5’-diphosphate-glucose (UDPG) and fructose-6-phosphate (Lee et al., 2003). On the other hand, the cleavage of sucrose is catalyzed by two enzymes with different properties, invertase and sucrose synthase (Pavlinova et al., 2002). Invertase (β-D-fructofuranosidase; EC 3.2.1.26) is a irreversible hydrolase and cleaves sucrose into glucose and fructose. By contrast, sucrose synthase (SS; UDP-glucose: D-fructose-2-α-glucosyltransferase; EC 2.4.1.13) catalyses the cleavage of sucrose on the presence of UDP to give UDP-glucose (UDPG) and fructose (Kingston-Smith et al., 1999). The reaction is reversible and SS is believed to act mainly as a hydrolytic enzyme in most plant tissue. SS, along with invertase, which also hydrolyzes sucrose, has an important impact on the steady level of sucrose in plant cells and their activity is particularly high in sink organs (Castonguay and Nadeau, 1998).
In the previous studies, it is found that lettuce quality rapidly deteriorated owing to wilting, high respiration rate and low sugar contents during storage at ambient temperature (Suthumchai et al., 2006). To further understand the changes in some postharvest physiological traits and activities of sugar related enzymes in lettuce stored at low temperature, this study was carried out.
MATERIALS AND METHODS
Plant materials: The crisphead lettuce cultivars ‘Shizuka’ and ‘Shisuko’ were harvested from a commercial farm in Kagawa Prefecture, Japan in December, 2004 and transported to the Postharvest laboratory. Harvested lettuce heads were stored in perforated plastic bags at 5°C for 10 days. Initial weight, color and respiration rate were measured prior to storage. After 48 h intervals, the heads of each cultivar were taken out from storage. The leaves were separated from the midribs and immediately stored at -30°C until analysis.
Weight loss determination: Weight loss was measured by weighing the heads at random after 48 h intervals until 10 days of storage at 5°C. The decrease in weight was revealed as percentage of sample weights determined every 48 h intervals.
Color assessment: Color change in lettuce heads were determined using a Chromameter (model CR-200, Minolta, Japan), equipped with an 8 mm measuring head and C illuminant. The meter was calibrated using the manufacturer’s standard white plate. Color changes were quantified in the L, a, b colorimetric system. L parameter refers to the lightness index scale of the head and ranges from 0 (black) to 100 (white). The ‘a’ parameter indicates the degree of red-purple (+a) or green (-a) color, while the ‘b’ parameter indicates the degree of yellow (+b) or blue (-b) color (McGuire, 1992). The CIE L, a, b parameters were revealed as hue angle (h° = tan-1(b/a) when a>0 and b>0, or h° = 180° + tan-1(b/a) when a<0 and b>0) was calculated from the a and b values (Lancaster et al., 1997). On each head, three readings were taken from different portions.
Respiration rate measurement: Respiration rate was determined using a Gas Chromatograph (GC); GC-8A. The lettuce head was weighed and carefully placed in a 10 L glass jar held at 5°C. Carbon dioxide production was measured at 48 h intervals by taking 10 mL gas sample from the glass jar sealed for 1 h and injected into a thermal conductive detector (TCD) gas chromatograph equipped with a 1 m activated charcoal column at 70°C (GC-8 AIT, Shimadzu Co., Ltd.). The result was revealed as mL CO2 kg-1 h-1.
Enzyme extraction: Approximately 5 g of fresh sample (plus % weight loss of the initial fresh weight of the tissue) from each portion were added with 1% of polyvinylpolypyrrolidone (PVPP) and 1 g of sea sand. The sample mixture was homogenized using a cooled mortar and pestle with 5 mL of 0.2 M citrate-phosphate buffer (C-P buffer, pH 5.0) for acid invertase. Whereas the extraction of SS and SPS used 10 mL of 0.3 M potassium-phosphate buffer (K-P buffer, pH 7.8) containing 1 M ascorbate, 1 M MgCl2, 1 M DTT and 0.1 M Na-EDTA. The resulting homogenate was filtered through cotton cloth and was centrifuged at 12,000 x g, at 2°C for 10 and 20 min for acid invertase and SS and SPS, respectively. For acid invertase analysis, the total supernatant was dialyzed with 40 times diluted 0.2 M C-P buffer (pH 5.0), for 12 h with frequent stirring and the inner solution was designated as ‘soluble fraction’. The residual tissues were reextracted after incubation with 5 mL of 0.2 M NaCl C-P buffer (pH 5.0) for about 24 h at 4°C. The supernatant was dialyzed as described above. The dialyzed solution was designated as ‘cell wall-bound fraction’. On the other hand, the total supernatant for SS and SPS determination was dialyzed with 40 times diluted 0.3 M K-P buffer (pH 7.8), for 12 h and the inner solution was used as the crude enzyme. All extraction procedures were carried out at 4°C.
Enzyme assay: The standard assay mixture for acid invertase consisted of 0.2 mL of 0.2 M C-P buffer (pH 5.0), 0.1 mL of 0.5 M sucrose, 0.1 mL of distilled water and 0.1 mL of crude enzyme solution. The blank experiment contained distilled water instead of sucrose. The assay mixture was incubated at 45°C for 15 min and neutralized with 0.1 N NaOH or 0.1 N HCl. A coloring Somogyi’s copper reagent was added and the mixture was heated for 10 min in boiling water. After cooling, the mixture was added with 1 mL of Nelson’s reagent. The amount of reducing sugars was estimated by the method of Somogyi (1952). Soluble protein content was determined according to the method of Lowry et al. (1951) using bovine serum albumin as the standard. The enzyme activity was revealed as the amount of glucose produced per min per milligram of protein.
SS and SPS activities were assayed at 37°C following the method described by Hubbard et al. (1989) with slight modifications. For SPS determination, we used a mixture of 70.75 μL consisting 50 mM Heps-NaOH solution (pH 7.5), 15 mM MgCl2, 25 mM fructose-6-phosphate, 25 mM glucose-6-phosphate, 25 mM UDP-glucose, distilled water and crude enzyme solution. For SS determination, 25 mM of fructose was used instead of fructose-6-phosphate and glucose-6-phosphate. The blank experiment contained distilled water instead of UDP-glucose and crude enzyme. The reaction mixtures were incubated at 37°C for 30 min and added with 70 μL of 30% KOH to terminate the reaction. After that, the assay mixtures were kept in boiling water (approximately 100°C) for 10 min to destroy any unreacted fructose or fructose-6-phosphate. After cooling, 2 mL of anthrone reagent (150 mg anthrone in 100 mL of 70% H2SO4) was added and incubated at 40°C for 10 min. The enzyme activities were determined by the absorbance of the sample at 620 nm using a spectrophotometer (UV-1200, Shimadzu Co., Ltd.). The soluble protein content was estimated using the method of Lowry et al. (1951) using bovine serum albumin as the standard. The SS and SPS activities were revealed as micromole of sucrose or sucrose-6-phosphate produced per min per milligram of protein, respectively.
Determination of soluble sugar contents by high performance liquid chromatography (HPLC): The sucrose, glucose and fructose contents were determined using HPLC. Approximately 2 g of lettuce sample (plus % weight loss of the initial fresh weight of the tissue) from each portion were mixed with 1 g of sea sand and homogenized in a cooled mortar and pestle. Ten milliliter of distilled water was added to the homogenate and centrifuged at 12,000 x g, at 2°C for 10 min. The mixture was filtered through a cellulose nitrate membrane filter (0.45 μm pore size). Soluble sugar contents were analyzed by HPLC using a stainless steel column (10.7 mm IDx30 cm) packed with silica gel (gel pack C610). The mobile phase (filtered water) was pumped through the column at a flow rate of 1.0 mL min-1. The pressure was adjusted to 28-29 kg cm-2 and the temperature to 60°C. A refractive index monitor (Hitachi L-3300) was used to record the peak areas. Sucrose, glucose and fructose were identified by their retention times and were quantified according to standards.
Statistical analysis: A Randomized Complete Block Design (RCBD) was adopted in this experiment with three replications. The data were treated by analysis of variance with Duncan’s Multiple Range Test (DMRT) between means, determined at the 5% level of significance. Linear correlation analysis was used to evaluate the relationships between sugar contents and enzyme activities.
RESULTS
Weight loss: Highest weight loss was observed during the first 2 days of storage in both cultivars (Fig. 1). After that, weight loss was minimal until the end of the experimental period. ‘Shisuko’, however, showed higher weight loss than ‘Sizuka’.
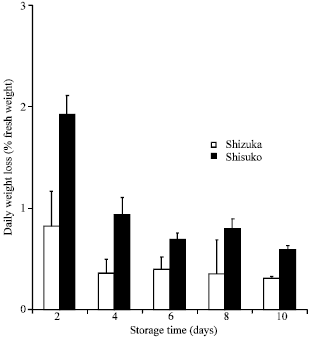 | |
| Fig. 1: | Weight loss of two lettuce cultivars (Shizuka and Shisuko) during storage at 5°C for 10 days. Each point represents the mean of 3 replications. Vertical bars indicate standard errors (SE). Between ‘Shizuka’ and ‘Shisuko’ = highly significant difference at 99% level, all days. Between days of storage of two cultivars = highly significant difference at 99% level, day 2 |
Color assessment: A continuous decrease in hue angle value was observed in ‘Shizuka’ except for a transient increase on day 8. On the other hand, there was no remarkable change in hue angle value in ‘Shisuko’ until the end of the storage period. Also, no significant difference was found between ‘Shizuka’ and ‘Shisuko’ (Fig. 2).
Respiration rate: In both cultivars, respiration rate rapidly declined after 2 days of storage. After that, CO2 production fluctuated until the end of the storage period (Fig. 3). ‘Shizuka’ showed higher initial CO2 production than ‘Shisuko’. However, after day 2 of storage, ‘Shisuko’ produced higher CO2 than ‘Shizuka’ except on day 10.
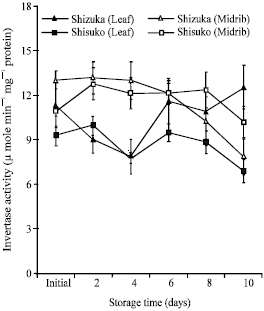
Acid invertase activity: Acid invertase (soluble fraction) activity in the midrib portion was significantly higher than that in the leaf portion (Fig. 4). The acid invertase activity decreased with few fluctuations except for a transient increase on day 6. In the leaf portion, the activity slightly declined in both cultivars. The acid invertase activity in cell wall-bound fraction also decreased with some fluctuations in the midrib portion of both cultivars (Fig. 5). However, there was no specific inclining or declining pattern observed in the leaf portion. The activity decreased until day 4 of storage. Thereafter, a transient increase was observed on day 6 in ‘Shisuko’ while a continuous increase was found in ‘Sizuka’.
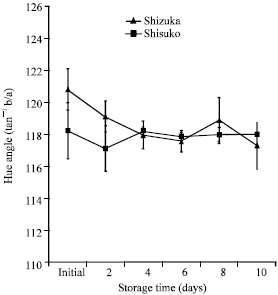 | |
| Fig. 2: | Color change of two lettuce cultivars (Shizuka and Shisuko) during storage at 5°C for 10 days. Each point represents the mean of 3 replications. Vertical bars indicate standard errors (SE). No significant difference throughout the experimental period |
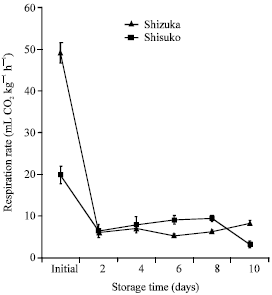 | |
| Fig. 3: | Respiration rate of two lettuce cultivars (Shizuka and Shisuko) during storage at 5°C for 10 days. Each point represents the mean of 3 replications. Vertical bars indicate standard errors (SE). Between ‘Shizuka’ and ‘Shisuko’ = significant difference at 95% level, days 8 and 10. Between days of storage of two cultivars = highly significant difference at 99% level, initial day |
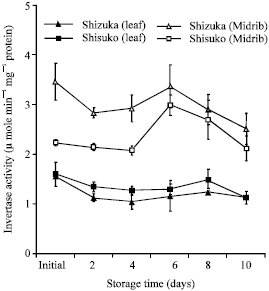 | |
| Fig. 4: | Changes in acid invertase activity in the soluble fraction in the leaf and midrib portions of two lettuce cultivars (Shizuka and Shisuko) during storage at 5°C for 10 days. Each point represents the mean of 3 replications. Vertical bars indicate standard errors (SE). Between leaf and midrib of ‘Shizuka’ = highly significant difference at 99% level, all days. Between leaf and midrib of ‘Shisuko’ = highly significant difference at 99% level, days 2, 4, 6 and 10 |
 | |
| Fig. 5: | Changes in acid invertase activity in the cell wall-bound fraction in the leaf and midrib portions of two lettuce cultivars (Shizuka and Shisuko) during storage at 5°C for 10 days. Each point represents the mean of 3 replications. Vertical bars indicate standard errors (SE). Between leaf and midrib of ‘Shizuka’ = significant difference at 95% level, days 2 and 4. Between leaf and midrib of ‘Shisuko’ = significant difference at 95% level, day 4 |
Sucrose synthase (SS) activity: In midrib portion, SS activity in both cultivars continuously increased with time (Fig. 6). There was only a slight increase in the SS activity in the leaf portion until the end of the storage period. In both cultivars, midrib showed higher enzyme activity than the leaf portion.
Sucrose phosphate synthase (SPS) activity: The SPS activity did not markedly change in both cultivars and portions throughout the storage period. However, the highest SPS activity was observed in the midrib of ‘Shisuko’ cultivar (Fig. 7).
Soluble sugar contents: Values for soluble sugars were estimated based on the water content of the sample at 5°C. In both cultivars, sucrose content rapidly declined after 2 days of storage in the leaf and midrib portions and almost unchanged until the end of the storage period (Fig. 8 and 9). Also, fructose and glucose contents declined with few fluctuations as storage progressed. The sugar contents in midrib portion were higher than that in the leaf portion.
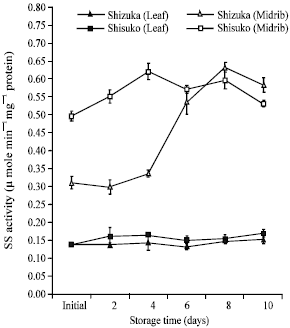 | |
| Fig. 6: | Changes in sucrose synthase (SS) activity in the leaf and midrib portions of two lettuce cultivars (Shizuka and Shisuko) during storage at 5°C for 10 days. Each point represents the mean of 3 replications. Vertical bars indicate standard errors (SE). Between leaf and midrib of two cultivars = highly significant difference at 99% level, all days |
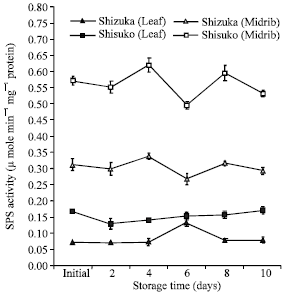 | |
| Fig. 7: | Changes in sucrose phosphate synthase (SPS) activity in the leaf and midrib portions of two lettuce cultivars (Shizuka and Shisuko) during storage at 5°C for 10 days. Each point represents the mean of 3 replications. Vertical bars indicate standard errors (SE). Between leaf and midrib of two cultivars = highly significant difference at 99% level, all days |
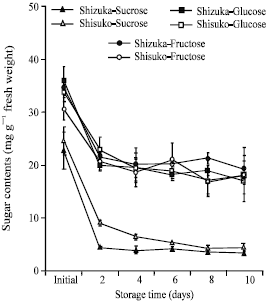 | |
| Fig. 8: | Changes in soluble sugar contents in the leaf portion of two lettuce cultivars (Shizuka and Shisuko) during storage at 5°C for 10 days. Each point represents the mean of 3 replications. Vertical bars indicate standard errors (SE). Between sucrose, glucose and fructose of ‘Shizuka’ = sucrose, highly significant difference at 99% level, all days. Between sucrose, glucose and fructose of ‘Shisuko’ = sucrose, highly significant difference at 99% level, except initial and day 10 |
| Table 1: | Correlation coefficients (r) between acid invertase (soluble and cell wall-bound fractions), SS, SPS activities and sugar concentrations in the leaf and midrib portions of two lettuce cultivars (’Shizuka’ and ’Shisuko’) held at 5°C for 10 days |
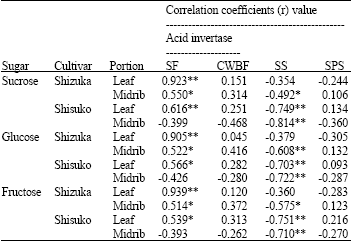 | |
| SF = soluble fraction, CWBF = cell wall-bound fraction SS = sucrose synthase, SPS = sucrose phosphate synthase *, ** denote significant correlation at 0.01<p≤0.05 and p≤0.01, respectively, n = 18 | |
Correlation coefficients (r) between enzyme activities and sugar contents: There was a significant positive correlation between the acid invertase activity of the soluble fraction and sugar contents in the leaf portion of two cultivars (Table 1). Also, a significant positive correlation was observed between the soluble invertase activity and sugar contents in the midrib of ‘Sizuka’. No significant correlation was found between the invertase (cell wall-bound fraction) and SPS activities and soluble sugars in both portions of each cultivar. In addition, there was a highly significant negative relationship between SS activity and sugar contents in both portions of ‘Shisuko’.
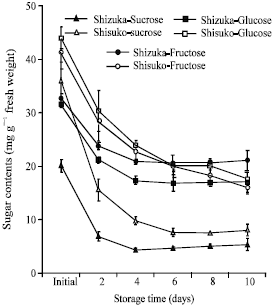 | |
| Fig. 9: | Changes in soluble sugar contents in the midrib portion of two lettuce cultivars (Shizuka and Shisuko) during storage at 5°C for 10 days. Each point represents the mean of 3 replications. Vertical bars indicate standard errors (SE). Between sucrose, glucose and fructose of ‘Shizuka’ = highly significant difference at 99% level, days 2, 4 and 8. Between sucrose, glucose and fructose of ‘Shisuko’ = sucrose, highly significant difference at 99% level, except initial day |
DISCUSSION
Temperature management is a one of the most important method to maintain quality of fresh products. The head lettuces kept at 5°C had still good appearance after 10 days of storage. However, physiological and biochemical changes which included weight loss, color, respiration and sugar metabolizing enzymes activities were observed. Water loss is one of the main causes of deterioration that reduces the marketability of leaf vegetables. Water loss resulted to wilting, shriveling and softening, thus, accelerates senescence (Ben-Yehoshua, 1985). We found only 2.22 and 4.85% of weight loss at the end of the storage period in the cultivar ‘Shizuka’ and ‘Shisuko’, respectively (Fig. 1). There was a slight change in the color of lettuce head at the end of the experimental
period (Fig. 2). The respiration rate was low throughout the storage duration except for the initial day (Fig. 3). The high CO2 production on day 0 could be due to the stress imposed by harvest. The respiration rate of commodities is directly related to storage temperature. Lettuce heads produced 5, 8, 13 mL CO2 kg-1 h-1 when storage at 1, 4.5 and 10°C, respectively (Behrsing et al., 1998).
Invertase exists in several isoforms with different biochemical properties accumulated in the cytoplasm (neutral or alkaline invertase), vacuole (vacuolar invertase) and extracellular spaces (cell wall invertase). Vacuolar and cell wall invertases have acidic pH (4.5-5.0) and are, hence, referred to as acid invertases (Sturm, 1999). In this study, we found that the acid invertase activity decreased (Fig. 4 and 5) with few fluctuations, while the sugar contents declined in both cultivars and portions (Fig. 8 and 9). In addition, there was a significant positive correlation between soluble acid invertase activity and sugar contents in the leaf portion of both cultivars (Table 1). This result suggests that the decrease of sugars was associated with the decrease in the invertase activity. Although, low temperature decreases the activity of invertase, it could still break sucrose into its component monosaccharides, glucose and fructose (Bhowmik et al., 2001). In potato tuber ‘White Fryer’ cultivar, sucrose content decreased even with low invertase activity during storage at 4°C (Matsuura-Endo et al., 2004). Moreover, there are some factors, such as enzyme inhibitors, which could lower the activity of acid invertase during storage in the lettuce head. Pressey (1994) reported that proteins co-extracted with invertase may interact with the enzyme and reduce activity in vitro. The interaction may be affected either by the presence of substrate (Krausgrill et al., 1996) or by the reaction products, with glucose acting as a non-competitive inhibitor and fructose as a competitive inhibitor (Sturm et al., 1999). It was further studied on the purified enzyme from ryegrass leaves that fructose partially inhibits invertase at concentrations as low as 2 mM. Hence, if sucrose was cleaved and the fructose was not used, they would subsequently tend to inhibit hydrolysis (Foyer et al., 1997). Walker et al. (1997) observed in the purified vacuolar enzyme from leaves of Lolium temulentum that fructose was shown to be a non-competitive inhibitor and the interreaction between the rate of hydrolysis and potential inhibition may be species-specific.
Interestingly, SS activity showed a continuous increase with time during storage (Fig. 6), whereas sugar contents decreased in both cultivars and portions that led to quality deterioration of head lettuce. From Table 1, SS activity and sugar contents had a significant negative correlation in both portions of ‘Shisuko’. This result suggests that SS was in the direction of sucrose catabolism. Although SS is able to synthesize sucrose under suitable conditions, there is evidence that SS in vivo is prominently involved in the breakdown of sucrose (Sturm and Tang, 1999). Lerchl et al. (1995) reported that the enzyme has been localized in the sieve tube-companion cell complex of leaves. It also involved in the degradation of sucrose. In developing seeds of lima bean and tubers of potato, SS was found to be the primarily sucrose-breaking enzyme (Sung et al., 1989). In addition, Riffkin et al. (1995) found that SS had consistent higher activity than invertase throughout endosperm development in wheat and invertase activity did not apparently change. Furthermore, Eldan and Mayer (1974) suggested that during the germination of lettuce seeds, sucrose content immediately decrease with the onset of germination, but acid invertase activity was observed after 10-15 h. Thus, acid invertase is appreciably not the only enzyme which could be involved in the sucrose hydrolysis but SS may have role in sucrose metabolism.
There was no remarkable change observed in SPS activity in both cultivars and portions (Fig. 7). This pattern was reported by Pramanik et al. (2004) in broccoli stored at 5°C. From this study, the sucrose-hydrolyzing enzyme activities vary with leaf and midrib portions; the midrib showed higher activities than leaf. This could be due to the difference in their physiological function; the leaf tissue serves as source while the midrib acts as sink organ. The activities in sink organs (developing tissues) were higher than in source organs (mature leaves) (Pavlinova et al., 2002). Additionally, Foyer et al. (1997) observed in leaf of ryegrass that the invertase activity was high in young leaves, while the activity fell as the leaves mature.
Base on the above discussion, it can be concluded that low temperature can maintain the quality and extend the shelf life of head lettuce. ‘Shizuka’ showed lower weight loss, degree of yellowing and respiration rate than ‘Shisuko’ which might the reason for the better appearance of the prior. SS plays a key role in sucrose hydrolysis. The SS activity had a continuous increase, while the acid invertase activity decreased throughout the experimental period. Further studies in the molecular level are required for understanding the genes expression that involved in sucrolysis process.
ACKNOWLEDGMENTS
We are thankful to the Government of Japan through its Ministry of Education, Culture, Sports, Science and Technology (Monbukagakusho: MEXT) for the financial support.
REFERENCES
- Behrsing, J.P., R.B. Tomkins, J.M. Hutchin and P. Franz, 1998. Effect of temperature and size reduction on respiratory activity and shelf-life of vegetables. Acta Hortic., 464: 500-500.
Direct Link - Ben-Yehoshua, S., 1985. Individual seal-packaging of fruit and vegetables in plastic film-a new postharvest technique. HortScience, 20: 32-37.
Direct Link - Bhowmik, P.K., T. Matsui, K. Kawada and H. Suzuki, 2001. Seasonal changes of asparagus spears in relation to enzyme activities and carbohydrate content. Sci. Hortic., 88: 1-9.
Direct Link - Bolin, H.R. and C.C. Huxsoll, 1991. Effect of preparation procedures and storage parameters on quality retention of salad-cut lettuce. J. Food Sci., 56: 60-62.
CrossRef - Castonguay, Y. and P. Nadeau, 1998. Enzymatic control of soluble carbohydrate accumulation in cold-acclimated crowns of alfafa. Crop Sci., 38: 1183-1189.
Direct Link - Eldan, M. and A.M. Mayer, 1974. Acid invertase in germinating Lactuca sativa seeds: Evidence for de novo synthesis. Phytochemistry, 13: 389-395.
CrossRef - Enninghorst, A. and F. Lippert, 2003. Postharvest changes in carbohydrate content of lambs lettuce (Valerianella locusta). Acta Hortic., 604: 553-558.
Direct Link - Hubbard, N.L., S.C. Huber and D.M. Pharr, 1989. Sucrose phosphate synthase and acid invertase as determinants of sucrose concentration in developing muskmelon (Cucumis melo L.) fruits. Plant Physiol., 91: 1527-1534.
PubMed - Kingston-Smith, A., R.P. Walker and C.J. Pollock, 1999. Invertase in leaves: Conundrum or control point?. J. Exp. Bot., 50: 735-743.
Direct Link - Krausgrill, S., A. Sander, S. Greiner, M. Weil and T. Rausch, 1996. Regulation of cell wall invertase by a proteinaceous inhibitor. J. Exp. Bot., 47: 1193-1198.
Direct Link - Lancaster, J.E., C.E. Lister, P.F. Reay and C.M. Triggs, 1997. Influence of pigment composition on skin color in a wide range of fruit and vegetables. J. Am. Soc. Hortic. Sci., 122: 594-598.
Direct Link - Lee, M.H., C.C. Yang, H.L. Wang and P.D. Lee, 2003. Regulation of sucrose phosphate synthase of the sweet potato callus is related to illumination and osmotic stress. Bot. Bull. Acad. Sin., 44: 257-265.
Direct Link - Lerchl, J., P. Geigenberger, M. Stitt and U. Sonnewald, 1995. Impaired photoassimilate partitioning caused by phloem-specific removal of phyrophosphate can be complemented by a phloem-specific cytosolic yeast-derived invertase in transgenic plants. Plant Cell, 7: 259-270.
PubMed - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Matsuura-Endo, C., A. Kobayashi, T. Noda, S. Takigawa, H. Yamauchi and M. Mori, 2004. Changes in sugar content and activity of vacuolar acid invertase during low-temperature storage of potato tubes from six Japanese cultivars. J. Plant Res., 117: 131-137.
CrossRef - McGuire, R.G., 1992. Reporting of objective color measurements. HortScience, 27: 1254-1255.
CrossRefDirect Link - Paull, R.E., 1999. Effect of temperature and relative humidity on fresh commodity quality. Postharvest Biol. Technol., 15: 263-277.
CrossRefDirect Link - Pavlinova, O.A., E.N. Balakhontsev, M.F. Prasolova and M.V. Turkina, 2002. Sucrose-phosphate synthase, sucrose synthase and invertase in sugar beet leaves. Russian J. Plant Physiol., 49: 68-73.
CrossRefDirect Link - Pramanik, B.K., T. Matsui, H. Suzuki and Y. Kosugi, 2004. Changes in carbohydrates content and activities of acid invertase, sucrose synthase and sucrose phosphate synthase in broccoli during short term storage at low temperature. Asian J. Plant Sci., 3: 449-454.
Direct Link - Rennie, T.J., C. Vigneault, G.S.V. Raghavan and J.R. DeEll, 2001. Effects of pressure reduction rate on vacuum cooled lettuce quality during storage. Can. Biosyst. Eng., 43: 3.39-3.43.
Direct Link - Riffkin, H.L., C.M. Duffus and I.C. Bridges, 1995. Sucrose metabolism during endosperm development in wheat (Triticum aestivum). Physiol. Plant., 93: 123-131.
Direct Link - Stanley, R., 1989. The influence of temperature and packaging material on the post harvest quality of iceberg lettuce. Acta Hortic., 244: 171-180.
Direct Link - Sturm, A., 1999. Invertase. Primary structures, functions and roles in plant development and sucrose partitioning. Plant Physiol., 121: 1-8.
Direct Link - Sturm, A. and G.Q. Tang, 1999. The sucrose-cleaving enzymes of plants are crucial for development, growth and carbon partitioning. Trends Plant Sci., 4: 401-407.
PubMedDirect Link - Sturm, A., D. Hess, H.S. Lee and S. Lienhard, 1999. Neutral invertase is a novel type of sucrose-cleaving enzyme. Physiol. Plant., 107: 159-165.
CrossRef - Sung, S.J.S., D.P. Xu and C.C. Black, 1989. Identification of actively filling sucrose sinks. Plant Physiol., 89: 1117-1121.
PubMed - Suthumchai, W., T. Matsui, K. Kawada and Y. Kosugi, 2006. Changes in acid invertase activity and sugar contents in lettuce during storage at ambient temperature. Asian J. Plant Sci., 5: 304-310.
CrossRefDirect Link - Walker, R.P., A.L. Winters and C.J. Pollock, 1997. Purification and characterization of invertases from leaves of Lolium temulentum L. New Phytologist, 135: 259-266.
Direct Link - Watada, A.E., N.P. Ko and D.A. Minott, 1996. Factors affecting quality of fresh-cut horticultural products. Postharvest Biol. Technol., 9: 115-125.
CrossRef