
B. Vafias
Not Available
C. Goulas
Not Available
G. Lolas
Not Available
E. Tamoutsidis
Not Available
P.N. Deligeorgidis
Not Available
C.G. Ipsilandis
Not Available
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 3 | Page No.: 463-469







ABSTRACT
Mass selection during early stages of genotype evaluation is essential for maize breeding programs. The purpose of this study was to determine the proper conditions for mass selecting useful genotypes in segregating maize genetic materials. F1/F2 comparisons were made to evaluate the genetic materials and F1/F2 and F2 mixtures of genotypes were formed to ensure allocompetition conditions. Moreover, two severe stresses were applied: plant density and low inputs, in addition to full-inputs conditions. Prisma F1 and F2 were better yielding materials than Funo F1 and F2, respectively. Until 13.33 plants m-2 there was an increase in F1 field yields, followed by lower yields in increased plant density, since modern hybrids tolerate greater plant populations. Significant interaction between genetic materials and plant density found in full-inputs experiments, was eliminated under the strong low-inputs stress. The indications from Half-sib progeny evaluation showed strong relationship between progeny yield and both selected plants’ yield and selection differential. High selection differential was realized under low-inputs condition due to low performance of original genetic materials. Strong allocompetition conditions may have a negative effect on HS progeny yielding performance, since mixtures of different genetic materials lead to lower yields. As a final conclusion, present findings indicated that selection of genotypes must be practiced under conditions of high selection differential. The original population mean is not as important as the selection differential. Extended experimentation must include S1 and Full-sib progenies and selection of progeny plants must include not only the highest yielding plants, but plants from the upper, mean and lower yielding areas of yield distribution, as well.
PDF Abstract XML References Citation
How to cite this article
B. Vafias, C. Goulas, G. Lolas, E. Tamoutsidis, P.N. Deligeorgidis and C.G. Ipsilandis, 2007. Stress Conditions for Improvement of Mass Breeding Method in Maize
. Asian Journal of Plant Sciences, 6: 463-469.
DOI: 10.3923/ajps.2007.463.469
URL: https://scialert.net/abstract/?doi=ajps.2007.463.469
DOI: 10.3923/ajps.2007.463.469
URL: https://scialert.net/abstract/?doi=ajps.2007.463.469
INTRODUCTION
The contribution of maize breeding on hybrid yield per plant is summarised in Duvick’s findings (1997), that during the past 70 years, breeding had no effect on hybrid yield per plant improvement, as it was estimated under low stress conditions (1 plant m-2). The most important result is that, maize breeding has contributed to improvement of hybrid yield per unit area (Troyer, 1995; Duvick, 1997), when hybrid grain yield was estimated at the typical plant density of 7.9 plants m-2. These results indicated that higher grain yield productivity of modern maize hybrids resulted indirectly by improving tolerance to various biotic and abiotic stresses and by improving the efficiency of exploiting the available resources (Derieux et al., 1987; Russell, 1991; Tollenaar and Wu, 1999). A successful breeding program in developing new hybrids, depends not only on the germplasm but also on the procedure for developing inbred lines and consequently new promising crosses (Ipsilandis and Koutsika-Sotiriou, 2000). Maize breeders lead their breeding programs to an excessive exploitation of heterosis, which is rendered on the function of alleles showing dominance effects (Smith, 1984; Falconer, 1989). According to Kearsey and Pooni (1992), heterosis is caused by dispersed genes showing mainly directional dominance and not by heterozygote superiority or complementary epistasis. Additive gene action is of great importance, because this kind of action insures heritable and stable performance (Fasoulas, 1993; Hallauer and Miranda, 1988; Ipsilandis and Koutsika-Sotiriou, 2000) and may insure high yields for crosses developed under genetically narrow-based crossing programs (Ipsilandis and Koutsika-Sotiriou, 1997, 2000).
Maize population improvement is based on recurrent selection schemes between and within populations (Moll and Stuber, 1971). Furthermore, maize population improvement may include mass selection (Gardner, 1961) or modified ear-to-row (Lonnquist, 1964). Maize breeders may use combined selection methods based on S1/HS evaluation (Goulas and Lonnquist, 1976; Coors, 1988; Ipsilandis and Koutsika-Sotiriou, 2000) in order to keep high combining ability in elite genetic materials with high per se yielding performance. Mass selection resulted in 4% improvement (Hallauer and Sears, 1969) and pedigree selection schemes achieved greater improvement up to 13.6% (Lonnquist, 1964; Paterniani, 1967; Dudley et al., 1974). S1 selection resulted in satisfactory improvement in the past (Moll and Stuber, 1971; Genter and Eberhart, 1974). Pedigree selection schemes combined with selfings may result in considerable improvement of maize lines (Fasoulas, 1981, 1988; Ipsilandis and Koutsika-Sotiriou, 2000; Ipsilandis et al., 2006). Second-cycle lines may also considerably improve yields (Jenkins, 1978; Smith, 1988; Ipsilandis and Koutsika-Sotiriou, 2000; Ipsilandis et al., 2006), substituting lines from open-pollinated populations (Jenkins, 1978; Hallauer, 1990). Test crosses contributed in an up to 4.5% improvement (Russell et al., 1973) resulting in elite line development such as B73. Population improvement under favourable conditions in absence of competition, found to be more effective in comparison to competition conditions (Onenanyoli and Fasoulas, 1989). For maximizing phenotypic expression, Fasoulas (1973, 1981, 1988, 1993) proposed selection under favourable conditions in absence of competition in honeycomb designs. Honeycomb method was expanded by Fasoula and Fasoula (1997, 2000), based also on favourable evaluation conditions. Tokatlidis et al. (2001) also proposed low-stress conditions for maize improvement and especially for development of density-independent hybrids. Low plant density during the first stages of evaluation, was found effective for the development of high performing inbred lines (Ipsilandis and Koutsika-Sotiriou, 2000; Ipsilandis et al., 2005b; 2006). On the other hand, the necessity of higher plant densities for optimal productivity of modern maize hybrids led Troyer and Rosenbrook (1983) to suggest that selection under higher plant densities was a means to improve grain yield of maize, since this may lead to better yielding genotypes under the density stress. This possibility was also mentioned indirectly by Tetio-Kagho and Gardner (1988).
The purpose of this study was to determine the proper conditions for mass selecting useful genotypes in segregating genetic materials. In that way, F1/F2 comparisons were made to evaluate the genetic materials. Additionally, F1/F2 and F2 mixtures of genotypes were formed to ensure allocompetition conditions. Moreover, two severe stresses were applied: plant density and low inputs, in addition to normal (full-inputs) conditions.
MATERIALS AND METHODS
In year 2000, in the experimental farm of Technological Education Institute of Larissa, Greece, two experiments were conducted. The first (full-inputs experiment) was based on the availability of fertiliser (about 24 units of N) and irrigation. The second (low-inputs experiment) was based on lack of fertiliser (about only 10 units of N) and irrigation (half the water given in the first experiment).
The genetic materials used were the F1 generations of commercial maize hybrids Prisma and Funo, the F2 generations of the two hybrids, the balanced mechanical mixture F1+F2 of hybrid Prisma and the balanced mechanical mixture of the F2 generations of the two hybrids (F2+F2 Prisma and Funo). The plant densities used in both experiments were 5.33, 6.66, 8.88, 13.33 and 19.0 plants m-2.
The experimental design was based on a complete blocks design, with four replications of each treatment. The two factors (plant densities and genetic materials) were arranged in a split-plot design. The main factor (genetic materials) formed the subplots and the second factor (plant densities) formed the main plots. Each plot consisted of double rows, 5 m long and 75 cm row to row spacing. Plants on the rows were sown on the proper distances (25, 20, 15, 10 and 7 cm) to form plant densities. Factorial analysis was based on Snedecor and Cochran (1980). Yield estimation was based on individual plant yield in grammars (g) and field yield in kg m-2. Maximum and mean yields of individual plants were also estimated to calculate theoretical selection differential. Finally, standard deviation of individual values was estimated. Humidity was found below 15%.
The experimental procedure was continued for one more year (2002) in the experimental farm of Technological Education Institute of Florina. For this purpose, the half-sib (HS) progenies of 16 materials from the first year’s experiment were used. These materials were selected in random and according to selection differential calculated (only progenies from F2 plants exhibiting maximum yield values), especially if there were great differences between selection differential theoretical values. These progenies were sown in 16 separate lines, with 40 plants per line. Prisma and Funo F1s were sown in two additional lines. Plant-to-plant distances were 0.2 m and the distance between these lines was 1 m. Experiment was based on the availability of fertiliser (about 24 units of N) and irrigation. Individual plant yield (g) and standard deviation were calculated. Correlations were made between original genetic materials and their selected progeny yield estimations.
RESULTS
Yield data from full-inputs and low-inputs first year’s experiments are presented in Table 1 and 2, involving yield per unit area (field yield) and individual plant yield.
| Table 1: | Measurements of field yield in kg m-2, mean individual plant yield in g plant-1, standard deviation (Dev.), maximum (Max) individual plant yield in g plant-1 and selection differential, across five plant densities (5.33, 6.66, 8.88, 13.33 and 19.0 plants m-2) for full-inputs field, including F1s and F2s of commercial hybrids Prisma and Funo, mixture of F1 and F2 seed from Prisma and mixture from both F2s of Prisma and Funo |
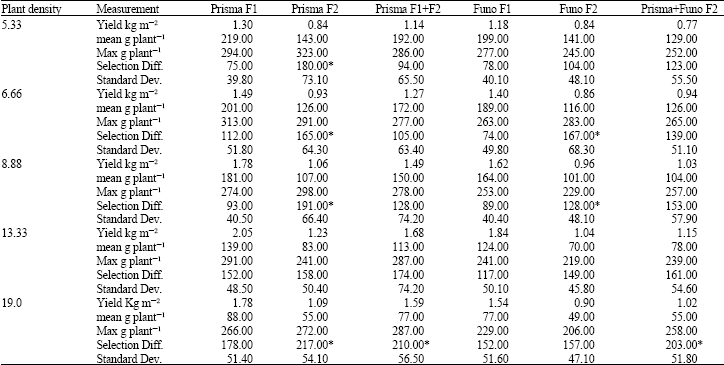 | |
| Statistically significant differences (for yield estimations) were found between different genetic materials and plant density levels, at p<0.001 and interaction between the two factors, at p<0.01 (CV exp = 8.5%, MS error = 0.6). * Selected original materials for progeny selection | |
| Table 2: | Measurements of field yield in kg m-2, mean individual plant yield in g plant-1, standard deviation (Dev.), maximum (Max) individual plant yield in g plant-1and selection differential, across five plant densities (5.33, 6.66, 8.88, 13.33 and 19.0 plants m-2) for low-inputs field, including F1s and F2s of commercial hybrids Prisma and Funo, mixture of F1 and F2 seed from Prisma and mixture from both F2s of Prisma and Funo |
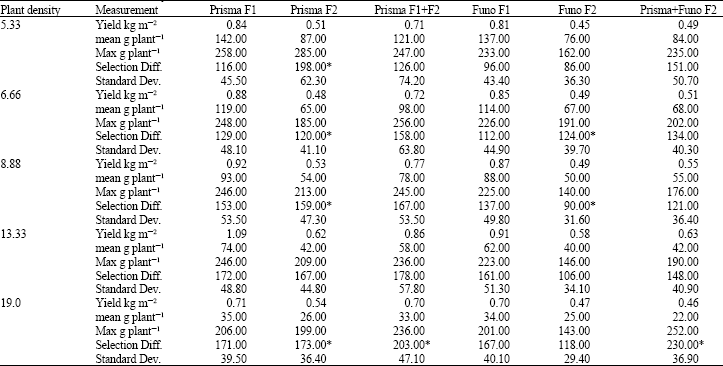 | |
| Statistically significant differences (for yield estimations) were found between different genetic materials, at p<0.001 and only for high density level, at the limit of p = 0.05 (CV exp = 13.5%, MS error = 0.4). * Selected original materials for progeny selection | |
In the full-inputs experiment, F1 hybrid Prisma yielded better than other materials (Table 1), increasing field yield from lower to higher plant densities until the optimum of 13.33 plants m-2 was reached (2.05 kg m-2). The rest of the genetic materials showed almost the same behaviour (Table 1). The F2 mixture (Prisma+Funo), was the lowest yielding material in low plant densities, but in higher densities over yielded F2 Funo and reached yield of F2 Prisma. F1 and F2 from single maize hybrid Prisma were better performing in comparison to F1 and F2 from Funo across all densities (Table 1 and 2). Concerning yield, significant interaction was found between the two factors (genetic materials and plant density). The individual plant yield was decreasing as plant density was increasing (Table 1), but the decreasing rate was slower until 13.33 plants m-2. At this density level the decreasing rate of individual plant yield became rapid, leading to lower field yield.
| Table 3: | Measurements of mean individual plant yield in g per plant and standard deviation (Std Dev.) of selected original genetic materials and their progeny lines and individual (maximum) yield in g per plant of selected plants, with Selection Differential |
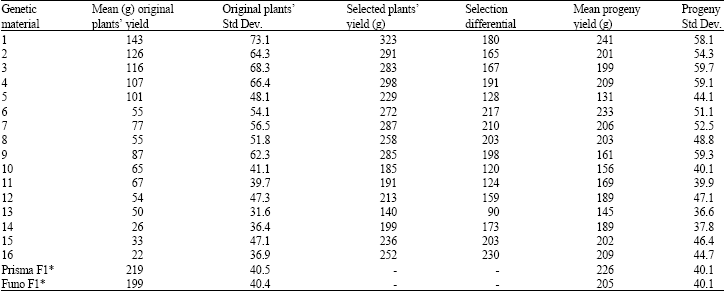 | |
| * F1 yields in first year’s experiment, with full inputs at 5.33 plants m-2 | |
| Table 4: | Correlations between the six measurements in Table 3 |
 | |
| Correlations coefficients statistically significant * at p<0.05 and ** at p<0.01 | |
In the low-inputs experiment, F1 hybrid Prisma yielded better than other materials (Table 2), increasing field yield from lower to higher plant densities until the optimum of 13.33 plants m-2 was reached. Prisma F1+F2 was almost the mean of F1 and F2 separately and all the F2s exhibited the same performance (Table 2). All yields were lower than the first experiment (Full-inputs) and the same was found for individual plant yield (Table 1 and 2). Concerning yield, differences were found between all genetic materials and only for high density level (p = 0.05).
Maximum yield values, were not always accompanied by high selection differential due to high mean plant yielding performance (Table 1 and 2). These adverse results allowed better selection, in random, for the genotypes used in progeny evaluation (Table 1-3).
Progeny evaluation showed that there were genotypes with good and stable performance and sometimes progeny performance was better than original plant’s performance (Table 3). Correlations between measurements in Table 4 revealed that, original mean plant yield is correlated to standard deviation (r = 0.835) but not to its progeny’s yield or to selection differential. On the other hand, progeny yield depends mainly on selection differential (r = 0.737) and additionally on maximum values of selected plants (r = 0.682). Only for full-inputs experiment, original plant mean yield was correlated to progeny yield (r = 0.898). Correlations between yields of corresponding original genetic materials in full-inputs and low-inputs experiments were calculated and found very significant (r = 0.983), but the progenies of such plants showed no relation.
DISCUSSION
In full-inputs experiment, there was a significant interaction between genetic materials and plant densities, indicating that different materials respond in a different way under the stress of density. F2 generations were affected lesser than F1 hybrids. Until 13.33 plants m-2 there was an increase in F1 field yields, followed by lower yields in increased plant density, since modern hybrids tolerate greater plant populations (Duvick, 1992; Vafias et al., 2006). Optimum plant density for some maize hybrids was found between 70,000 and 100,000 plants per hectare (Vafias et al., 2006) indicating density dependence of maize hybrids (Tokatlidis, 2001; Tokatlidis et al., 2001). In the present study, significant interaction between genetic materials and plant density was eliminated under the low-inputs stress. To overcome density effects in farmer’s field Fasoula and Fasoula (1997, 2000) emphasized the importance of low stress conditions in optimizing the effectiveness of selection for new cultivars, aiming in improved potential yield per plant, tolerance to stresses and responsiveness to inputs. Genotype mixing, plant density and allocompetition are major stresses only under high-inputs or normal conditions and in agreement with the findings reported by Fasoulas (1981, 1988, 1993), Fehr (1987), Vafias et al. (2000) and Ipsilandis et al. (2005a). In this case, plant size variability is directly related to neighbourhood competition in which the growth of an individual depends on the number, size and proximity of neighbours (Bonan, 1991).
Genetic purity proved to be a greater stress condition than density effects and in agreement with the findings reported by Ipsilandis et al. (2005a). This was more apparent in the low-inputs experiment where differences between genetic materials were much more significant and plant density was a limited stress. Soil heterogeneity effects were found only in low-inputs experiments (Vafias et al., 2007), which are considered to be more significant in low density and poor field conditions (Fasoulas, 1981, 1988). Low inputs resulted in significant soil heterogeneity, maybe stronger as a stress condition than plant density effects and allocompetition. These adverse conditions obviously affected in a different way genotype expression, generating different conditions of selection. Mass selection cannot depict the best conditions for genotype evaluation, but progeny evaluation can make selection more reliable (Fehr, 1987; Fasoulas, 1988).
F1 and F2 from single maize hybrid Prisma were better performing in comparison to F1 and F2 from Funo across all densities, indicating better adaptation of hybrid Prisma in the specific environments. Prisma and Funo HS progenies had similar behaviour without any relation to original genetic materials’ yielding performance. The indications from Half-sib progeny evaluation showed strong relationship between progeny yield and both selected plants’ yield and selection differential. High selection differential was found under low-inputs condition due to low performance of original genetic materials. Strong allocompetition conditions in the evaluation field may have a negative side effect on HS progeny yielding performance, since mixtures of different genetic materials (F1+F2 and especially F2+F2) lead to lower yields in comparison to pure F2s.
Rodriguez and Hallauer (1991) referred that low-yielding Full-sib (FS) progenies can easily be distinguished from early stage evaluation. Even though high-yielding Full-sib progenies may not preserve their vigour in late stages, low-yielding Full-sib progenies would never become high yielding and promising materials. Extended experimentation must include S1 and Full-sib progenies and selection of progeny plants must include not only the highest yielding plants (forming an artificial high selection differential), but plants from the upper, mean and lower yielding areas of yield distribution, as well. Many original plants must be selected to represent better the different growing conditions and avoid soil heterogeneity. S1 progenies may proved better for progeny evaluation because of extensive gene fixation and heritable additive effects (Genter and Alexander, 1966; Genter, 1973; Fasoulas, 1981; 1988). The only difficulty for such progeny plants may be the availability of seed, especially in low-inputs and high density conditions. Tokatlidis et al. (2001), stated that low stress conditions during genotype evaluation might lead to density-independent hybrids in maize, but this may not always be the target (Ipsilandis et al., 2005a). If response to selection is based on additive effects (Fehr 1987; Fasoulas, 1988, 1993; Falconer, 1989; Ipsilandis and Koutsika-Sotiriou, 2000), then high line performance is an “one-way” target for breeding maize. Accumulation of favourable additive genes is essential, but finding of high specific combining ability may become very difficult (Baker and Verhalen, 1975).
As a final conclusion, our findings showed that selection of genotypes must be practiced under conditions of high selection differential. The original population’s mean is not as important as the selection differential. Low inputs (high stress conditions) contributed to high selection differential and indirectly, to better progeny performance, indicating a more reliable procedure for selecting individual plants. In certain cases, this is in contrast with the statement for using low stress conditions in order to optimise the effectiveness of selection for new cultivars (Fasoulas, 1988; Fasoula and Fasoula, 2000).
REFERENCES
- Bonan, G.B., 1991. Density effects on the size of annual plant populations: An indication of neighbourhood competition. Ann. Bot., 68: 341-347.
Direct Link - Duvick, D.N., 1992. Genetic contributions to advances in yield of U.S. maize. Maydica, 37: 69-79.
Direct Link - Falconer, D.S., 1989. Introduction to Quantitative Genetics. 3rd Edn., Longman Scientific and Technical, New York, USA Pages: 438.
Direct Link - Fasoula, D.A. and V.A. Fasoula, 1997. Competitive ability and plant breeding. Plant Breed. Rev., 14: 89-138.
Direct Link - Fasoula, V.A. and D.A. Fasoula, 1999. Honeycomb breeding: Principles and applications. Plant Breed. Rev., 18: 177-250.
Direct Link - Gardner, C.O., 1961. An evaluation of effects of mass selection and seed irradiation with thermal neutrons on yield of corn. Crop Sci., 1: 241-245.
Direct Link - Genter, C.F. and M.W. Alexander, 1966. Development and selection of productive S1 inbred lines of corn (Zea mays L.). Crop Sci., 6: 429-431.
Direct Link - Genter, C.F., 1973. Comparison of S1 and testcross evaluation after two cycles of recurrent selection in maize. Crop Sci., 13: 524-527.
Direct Link - Goulas, C.K. and J.H. Lonnquist, 1976. Combined half-sib and S1 family selection in a maize composite population. Crop Sci., 16: 461-464.
Direct Link - Ipsilandis, C.G. and M. Koutsika-Sotiriou, 2000. The combining ability of recombinant S-lines developed from an F2 maize population. J. Agric. Sci. Cambridge, 134: 191-198.
Direct Link - Ipsilandis, C.G., B.N. Vafias, A. Karagiozopoulou and C.K. Goulas, 2005. F1 single-cross maize hybrid performance under low purity conditions. Asian J. Plant Sci., 4: 75-82.
CrossRefDirect Link - Ipsilandis, C.G., P.N. Deligeorgidis, L. Giakalis, M. Koutsika, A. Papadopoulou and V. Xanthopoulos, 2005. Breeding for homozygotic superiority and stability in maize without losing combining ability. Asian J. Plant Sci., 4: 499-506.
CrossRefDirect Link - Ipsilandis, C.G., I.S. Tokatlidis, B. Vafias and D. Stefanis, 2006. Criteria for developing second-cycle hybrids in maize. Asian J. Plant Sci., 5: 680-685.
CrossRefDirect Link - Snedecor, G.W. and W.G. Cochran, 1980. Statistical Methods. 7th Edn., Iowa State University Press, Iowa, USA., ISBN-10: 0813815606, Pages: 507.
Direct Link - Tetio-Kagho, F. and F.P. Gardner, 1988. Response of maize to plant population density. II. Reproductive development, yield and yield adjustments. Agron. J., 80: 935-940.
CrossRef - Tokatlidis, I.S., 2002. The effect of improved potential yield per plant on crop yield potential and optimum plant population in maize hybrids. J. Agric. Sci., 137: 299-305.
Direct Link - Tokatlidis, I.S., M. Koutsika-Sotiriou and A.C. Fasoulas, 2001. The development of density-independent hybrids in maize. Maydica, 46: 21-25.
Direct Link - Tollenaar, M. and J. Wu, 1999. Yield improvement in temperate maize is attributable to greater stress tolerance. Crop Sci., 39: 1597-1604.
CrossRefDirect Link - Troyer, A.F. and R.W. Rosenbrook, 1983. Utility of higher plant densities for corn performance testing. Crop Sci., 23: 863-867.
Direct Link - Vafias, B., C.G. Ipsilandis, C. Goulas and P.N. Deligeorgidis, 2006. An approach on yielding performance in maize under varying plant densities. Asian J. Plant Sci., 5: 690-694.
CrossRefDirect Link - Vafias, B., C. Goulas, G. Lolas and C.G. Ipsilandis, 2007. A triple stress effect on monogenotypic and multigenotypic maize populations. Asian J. Plant Sci., 6: 29-35.
CrossRefDirect Link