
A. Manickavelu
Not Available
N. Nadarajan
Not Available
S.K. Ganesh
Not Available
R.P. Gnanamalar
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 1 | Page No.: 33-36







ABSTRACT
The experiment was conducted to develop and evaluate the biparental progenies for drought tolerance breeding in rice. Genetic analysis of biparental progenies resulted that, the traits viz., days to 70% RWC, leaf rolling, leaf drying and plant height were governed by additive gene action and for improvement of these traits, pureline selection, mass selection and/or progeny selection and pedigree breeding method may be followed. Preponderance of non-additive gene action was observed in drought recovery rate, days to flowering, productive tillers per plant, grains per panicle, spikelet fertility, hundred grain weight, biomass yield, root length, dry root weight, root/shoot ratio, harvest index and grain yield. Hence, improvement of these characters could be possible through heterosis breeding or selection of single plants at later generations after hybridization or intermating of selected segregants through recurrent selection techniques.
PDF Abstract XML References Citation
How to cite this article
A. Manickavelu, N. Nadarajan, S.K. Ganesh and R.P. Gnanamalar, 2006. Genetic Analysis of Biparental Progenies in Rice (Orzya sativa L.). Asian Journal of Plant Sciences, 5: 33-36.
DOI: 10.3923/ajps.2006.33.36
URL: https://scialert.net/abstract/?doi=ajps.2006.33.36
DOI: 10.3923/ajps.2006.33.36
URL: https://scialert.net/abstract/?doi=ajps.2006.33.36
INTRODUCTION
Rice is cultivated in a wide range of ecosystems under varying temperature and water regimes. About 27% of world`s rice area is under rainfed lowland without assured water supply during critical periods of crop growth. Under circumstances of forced and unforced situations of moisture stress where rice is grown, it would be very much desirable to have varieties endowed with virtues and values of drought tolerance and stable performance. Development of high yielding drought tolerant rice cultivars will considerably improve rainfed rice production. Varietal improvement still remains the major strategy for increasing production and productivity under rainfed lowland condition. However, the fundamental mechanisms of drought tolerance in rice are poorly understood. Success in any breeding programme is dependent on the knowledge and understanding of the inheritance of the characters of interest. But the main drawback in breeding for drought tolerance is that it is a very complex character. Drought tolerance, as such is not a simple character governed by one or two genes but explained to be controlled by a number of physio-morphological characters being independently controlled by many genes[1]. Most of the drought tolerant traits are quantitative in nature, hence it is necessary to know the inheritance of those traits. Many mating designs were proposed by many authors to know the genetics of quantitative characters. Biparental mating is one of the simplest random mating designs available to effect forced recombination and breaking down undesirable linkages as pointed out by Comstock and Robinson[2]. To develop high yielding genotypes coupled with drought tolerance, population with high variability serves always as prime source for effective selection, particularly the role by F2 segregants in throwing much variability is highly recognized. The F2 are the critical generation in rice breeding and they determine the eventual success or failure of the hybridization programme[3]. The intercrossing in the F2 segregants provides chances of finding superior recombinants in F3 or later generations and a great amount of concealed genetic variations particularly of the additive type would be released thereby improving response to selection[4]. Frederickson and Kronstad[5] stressed that in autogamous crops, intermating among early segregants could open vistas to new levels of genetic variabilities by breaking up of the genetic recombination within the linkage group. Hence the present study was undertaken to know the genetics behind the drought tolerance in rice under lowland condition by following biparental mating.
MATERIALS AND METHODS
The experiment was conducted at Research Farm, Agricultural College and Research Institute, Madurai during October 2003-February 2004 and March 2004-June 2004.
The F2 seeds of Norungan/ASD 16 were obtained from the Department of Plant Breeding and Genetics, Agricultural College and Research Institute, Madurai and it was raised in non-replicated rows of 500 single plants. Two sets of four males and five females were randomly selected and mated in North Carolina Design II (NCD II)[2,6]. Thus, 20 crosses were made in each set and totally 40 crosses (BIPs) were obtained. For crossing, wet cloth method suggested by Chaisang et al.[7] was followed. Forty BIPs were raised in randomized block design with two replications adopting a spacing of 20 cm between rows and 10 cm between plants. In each replication, two sets of BIPs were randomized and in each set, individual BIPs were randomized. In each BIP, 20 plants were raised with a row length of two meters in each replication. IR 50, the most susceptible variety for drought was raised in between the sets and along the borders as an indicator for moisture stress. At peak tillering stage, the irrigation was withheld to impose drought. IR 50, the stress indicator started to show stress symptoms within 2-3 days. In rice, 70% Relative Water Content (RWC) is one of the best indicators of moisture stress[8]. Hence, the RWC was taken at regular intervals in each BIP. When each BIP attain 70% RWC, the drought tolerant parameters viz., leaf rolling and leaf drying were scored and the field was reirrigated. After one week, the drought recovery rate was recorded. At physiological maturity, four drought tolerant traits viz., spikelet fertility, root length, root dry weight and root/shoot ratio and yield and its component traits including days to flowering, plant height, number of productive tillers per plant, panicle length, grains per panicle, hundred grain weight, biomass yield, grain yield per plant and harvest index were recorded on ten plants at random in each BIP. The mean data of 40 BIPs for all 17 characters were subjected to analysis of variance of randomized block design. After ascertaining the significant difference among the BIPs, the data were subjected to analysis of variance appropriate to North Carolina Design II (NCD II)[2,6].
RESULTS AND DISCUSSION
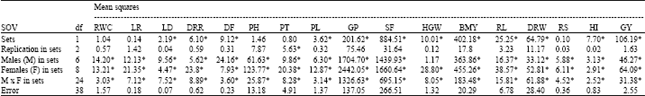
The success of any plant breeding programme depends to a greater extent on the knowledge of genetic architecture of the population handled by the breeder[9]. An overall understanding of the gene action for different characters is possible by making comparison of variance due to additive and/or dominance. The breeding method might be decided based on the relative importance of additive and dominance variances[10]. Higher magnitude of additive variance indicates the predominance of additive gene action and that of dominance variance indicates the non-additive gene action. The choice of the breeding method primarily depends upon the nature and magnitude of gene action. If additive variance is greater and additive gene action forms the principal component of genetic variance, use of pedigree method would be desirable as the gene effects are fixable, then the chance of fixing superior genotypes in early segregating generation will be high. If dominance variance is predominant the selection has to be postponed to later generations and appropriate breeding techniques should be adopted to obtain useful genotypes. The non-additive effect is non fixable, yet it can be exploited through heterosis breeding. In the present study, the genetic architecture of BIPs from the cross Norungan/ASD 16 was studied by employing the NCD II[2,6]. The combining ability analysis of BIPs for different traits exhibited highly significant mean square values for all the traits studied except hundred grain weight in males, all the traits in females and male x females. Sets in BIPs also expressed significant mean square values for all the traits except for days to 70% RWC, leaf rolling, plant height, productive tillers per plant and root/shoot ratio. It was evident from the results that adequate genetic variability was present among the BIPs (Table 1).
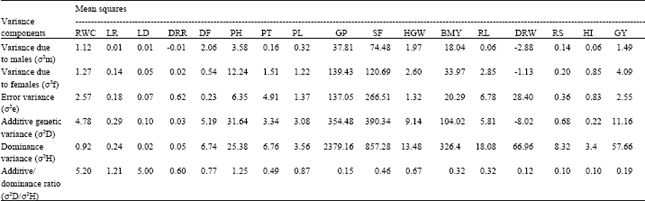
The estimation of variance components in BIPs resulted that some estimates of genetic variance were negative and also observed that negative estimates were obtained only for additive genetic variance (Table 2). This may be due to sampling error or lack of random mating. Similar results of negative estimates of additive genetic variance were also reported by Eberhart et al.[11] and Reddy and Agarwal[12]. Lindsey et al.[13] pointed out that larger estimates of negative variances for some characters might have resulted from some factors other than sampling error such as, lack of random mating in making the half-sib family groups. Hence, the negative sign of the additive genetic variance for dry root weight in the present study were ignored while drawing conclusions.
A perusal of variance estimates revealed that additive genetic variance was higher than dominance genetic variance and also additive /dominance ratio was more than one for four traits viz., days to 70% RWC, leaf rolling, leaf drying and plant height. Additive genetic variance is associated with homozygosity and also fixable in nature. Therefore, selection for these four traits governed by additive genetic variance will be very effective. Existence of additive variance is a prerequisite for improvement through selection because this is the only variance that responds to selection. Additive variance is a measure of additive gene action and this gene action is the chief cause of resemblance between relatives and progress by selection is directly proportional to the degree of resemblance between the parent and its progeny. Thus additive gene action is a measure of breeding value of a genotype.
| Table 1: | Combining ability analysis of BIPs for different traits |
 | |
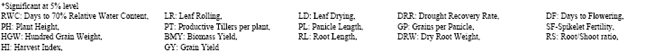 | |
| Table 2: | Estimates of variance components of BIPs for different traits |
 | |
 | |
Hence for the above said four traits, which showed preponderance of additive gene action, reliance should be placed on pure line selection, mass selection and/or progeny selection. This is in agreement with the findings of Ghosh[14] and Lavanya[15]. Further, these four traits could be improved by pedigree breeding method while going for hybridization and selection.
The remaining traits viz., drought recovery rate, days to flowering, productive tillers per plant, panicle length, grains per panicle, spikelet fertility, hundred grain weight, biomass yield, root length, dry root weight, root/shoot ratio, harvest index and grain yield expressed high dominance variance than additive variance. Hence it was concluded that these characters were governed by non-additive gene action. It was in accordance with the findings of Ghosh[14], Anandakumar and Subramanian[16] and Kalita and Upadhaya[17]. Normally dominance variance is associated with heterozygosity and also it is not fixable, therefore, selection for these traits is not effective. Dominance variance is the chief cause of heterosis or hybrid vigour. The preponderance of non-additive gene action for these traits under study indicated that improvement of these characters could be possible through heterosis breeding. But, rice being a self pollinated crop, heterosis breeding is not widely adopted unlike recombination breeding. Therefore, to get better genotypes by way of recombination breeding, hybridization followed by selection at later generations is suggested for exploiting dominance gene action. Alternatively, intermating of selected segregants might not only break the undesirable linkages, if any, but also allow accumulation of favourable alleles for the improvement of traits.
REFERENCES
- Fukai, S. and M. Cooper, 1995. Development of drought resistant cultivars using physio-morphological traits in rice. Field Crops Res., 40: 67-86.
CrossRef - Babu, R.C., M.S. Pathan, A. Blum and H.T. Nguyen, 1999. Comparison of measurement methods of osmotic adjustment in rice cultivars. Crop Sci., 39: 150-158.
Direct Link