
B.N. Vafias
Not Available
C.G. Ipsilandis
Not Available
Asian Journal of Plant Sciences
Year: 2005 | Volume: 4 | Issue: 1 | Page No.: 50-55







ABSTRACT
Recombinant lines developed from combining Half-sib/S1 evaluation on widely-spaced plants in the direction of high yielding per se, were used in combining ability tests in order to determine gene action under three-way crossing and possible commercial exploitation of such crosses. Combining ability tests consisted of crosses between: a) recombinant lines of common pedigree, b) recombinant lines and freely available inbred lines and c) recombinant lines and commercial F1 maize hybrids (three-way crosses). Heterosis was found to be acceptable, since the best three-way crosses reached the performance of corresponding single-cross hybrids and yielding performance of these three-way crosses was stable across years and experimental fields. Rapid line development from combining Half-sib/S1 evaluation may ensure high and stable crossing performance, based on additive gene action. Stability and uniformity of performance of three-way crosses was due to proper breeding incorporated in single-cross hybrid and to high and stable inbred line performance.
PDF Abstract XML References Citation
How to cite this article
B.N. Vafias and C.G. Ipsilandis, 2005. Combining Ability, Gene Action and Yielding Performance in Maize. Asian Journal of Plant Sciences, 4: 50-55.
DOI: 10.3923/ajps.2005.50.55
URL: https://scialert.net/abstract/?doi=ajps.2005.50.55
DOI: 10.3923/ajps.2005.50.55
URL: https://scialert.net/abstract/?doi=ajps.2005.50.55
INTRODUCTION
The starting genetic material of a breeding program is very important, since it determines the potential improvement for traits under selection[1]. Source germplasm used by maize breeders for inbred development includes primarily F2 (elite X elite inbred crosses), backcross and synthetic populations[2]. Modern maize breeding is based primarily on genetically narrow-based populations[2,3] including elite-line synthetics with a restricted genetic base, F2 populations of single crosses and backcross populations. For a successful program of recycling, the choice of the germplasm is the first priority[4]. A successful breeding program in developing new hybrids, depends not only on the germplasm but also on the procedure for developing inbred lines and consequently new promising crosses[5]. Maize breeders lead their breeding programs to an excessive exploitation of heterosis, which is rendered on the function of alleles showing dominance effects[6,7]. According to Kearsey and Pooni[8], heterosis is caused by dispersed genes showing mainly directional dominance and not by heterozygote superiority or complementary epistasis. Additive gene action is of great importance, because this kind of action insures heritable and stable performance[5,9,10] and may insure high yields for crosses developed under genetically narrow-based crossing programs[5,11]. Considering crossing between recombinant lines of common pedigree, general combining ability is of small importance, since heterosis was found low and heterobeltiosis was even lower[5]. These crosses depend their yielding performance on favourable additive gene effects, common in the two parents and consequently the second-cycle hybrids between them must be in a lower level of heterozygosity compared to the original hybrid[5,9,11-14]. Genter and Alexander[15] stated that, if the performance of S1 lines depends mainly on additive effects, then the yield of their crosses would be proportional to their yielding performance per se and thus, it is possible, that selection practiced for improving line performance per se, leads to the accumulation of favourable additive genes. Sotiriou et al.[16] concluded that in such cases the genetic background consist mainly of additive or partially dominant alleles. When crossing unrelated inbred lines, middle-parent heterosis can be more easily realized in comparison to crossing between recombinant lines of common pedigree[5]. A maize breeder has to balance between the three main goals of breeding (productivity, uniformity, stability) and the cost of hybrid seed production because of the low productivity of inbred parents as well as the need of separate plantings under isolation[13]. The practical difficulties associated with the low productivity of inbred lines can be overcome by three different approaches:
| a) | The use of 3-way or 4-way crosses[7,13,17]. |
| b) | The use of the backcross method for step-by-step improvement[7,13]. |
| c) | The improvement of productivity per se of modern inbred lines that serve as parents in crosses[4,5]. The most common practice involves family selection schemes[7,13]. |
The purpose of this study was to explore combining ability in recombinant S-lines in order to determine gene action under three-way crossing and possible commercial exploitation of such crosses, since the S-lines developed from combining Half-sib/S1 evaluation may ensure high and stable per se and crossing performance[5].
MATERIALS AND METHODS
Recombinant lines developed from combining Half-sib/S1 evaluation on widely-spaced plants in the direction of high yielding per se, were used in combining ability tests in order to determine gene action under three-way crossing. The recombinant lines were developed from the F2 generation of F1 commercial single-cross maize hybrid Lorena (PR3183), according to the procedure described by Ipsilandis and Koutsika-Sotiriou[5].
Combining ability tests consisted mainly of crosses between: a) Recombinant Lines (RL) of common pedigree, b) recombinant lines and freely available (FR) inbred lines and c) recombinant lines and commercial F1 maize hybrids (three-way crosses). No balanced design was used.
Evaluation of crosses was conducted in Technological Education Institute farm of Larissa, Greece, in two different fields, in five years (from 1998 to 2002).
In 1998, the materials evaluated were:
| • | Seven F1 single-cross commercial hybrids (Lorena, Dracma, Costanza, Rio Grande, Marista, Prisma and Aris). |
| • | Two F2 from single-cross hybrids (Costanza F2, Prisma F2). |
| • | Thirteen three-way crosses (nine 3-ways with RL lines, four with FR lines). |
| • | One double cross (Prisma X Costanza). |
| • | Five single crosses (one FR X FR, four RL X FR). |
In 1999, the materials evaluated were:
| • | Eight F1 single-cross commercial hybrids (Aligreen, Costanza, CS1251, Prisma, Dracma, LG2360, Golden West 623 and 621). |
| • | Fourteen three-way crosses (seven 3-ways with RL lines, seven with FR lines). |
| • | Three single crosses (RL X FR). |
In 2000, the materials evaluated were:
| • | Six F1 single-cross commercial hybrids (Aligreen, Costanza, CS1251, Prisma, Rio Grande and Dias). |
| • | One double hybrid (Aligreen X Rio Grande). |
| • | Fourteen three-way crosses (seven 3-ways with RL lines, seven with FR lines). |
| • | Five single crosses (one RL X RL, four RL X FR). |
In 2001, the materials evaluated were:
| • | Three F1 single-cross commercial hybrids (Aligreen, Costanza and Volusia). |
| • | One double hybrid (Costanza X Aligreen). |
| • | Ten three-way crosses (six 3-ways with RL lines, seven with FR lines). |
| • | Eight single crosses (five RL X RL, two RL X FR, one FR X FR experimental hybrid). |
In 2002, the materials evaluated were:
| • | One F1 single-cross commercial hybrid (Costanza). |
| • | Four three-way crosses (with RL lines). |
| • | Seven single crosses (three RL X RL, three RL X FR, one FR X FR). |
The experimental design, was the Randomized Complete Block (RCB) with 4 replications for all field trials. In all yield tests the experimental plot consisted of two 5 m long rows, spaced 80 cm apart. All plots contained 50 plants, i.e. 25 plants/row giving a density of 6.25 plants/m2. Plots were overplanted and thinned to desired stand at the seedling stage. Nitrogen and P fertiliser were applied at regular quantities. Trials were regularly irrigated (to avoid drought stress). Grain moisture was adjusted at 15.5%.
The eleven inbred lines used in experiments (single and 3-way crosses) were previously selected in the basis of productivity per se, after comparison to free-release inbred line B73 (in RCB design, similar to the one described above).
Middle-parent Heterosis (MPH%), Heterobeltiosis to second parent in cross (SPHB%) and Heterobeltiosis to best parent in cross (BPHB%) for all types of crosses were computed by formulas proposed by Koutsika-Sotiriou and Bos[18] and applied by Ipsilandis and Koutsika-Sotiriou[5].
RESULTS
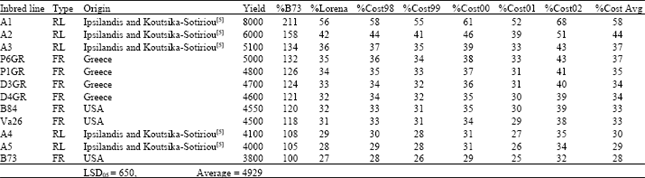
Table 1 clearly demonstrated that the yielding performance per se of all inbred lines used in crosses, was very high (up to 8000 kg ha-1). Yields of best performing line A1 reached 211% of free-release inbred line B73, 56% of F1 hybrid Lorena and 58% of average yield of F1 hybrid Costanza (Table 2-6). The average yield of Costanza is equal to the yielding performance of year 1998, the first year of experimentation (base yield), facilitating comparisons of lines and crosses across years. Nine inbred lines (recombinant and free-release) differed significantly when compared to B73, while two inbred lines although with better performance were at the same level of productivity (no significant difference) when compared to B73.
| Table 1: | The inbred lines used in crosses their type and origin, yield per se in kg ha-1, relative yield as percentage: of B73 (%B73), of F1 hybrid Lorena (%Lorena), of F1 hybrid Costanza in year 1998, 1999, 2000, 2001, 2002 and average of five years, respectively; %Cost98, %Cost99, %Cost00, %Cost01, %Cost02 and %CostAvg |
 | |
| Table 2: | Yield in kg ha-1, relative yield as percentage of F1 hybrid Costanza (%Cost), Middle-parent Heterosis (MPH%), Heterobeltiosis to second parent in cross (SPHB%) and Heterobeltiosis to best parent in cross (BPHB%) for all types of crosses in year 1998 |
 | |
In Table 2, the best yielding F1 hybrid found to be Lorena (14300 kg ha-1). F1 hybrid Costanza reached 13715 kg ha-1.
| Table 3: | Yield in kg ha-1, relative yield as percentage of F1 hybrid Costanza (%Cost), Middle-parent Heterosis (MPH%), Heterobeltiosis to second parent in cross (SPHB%) and Heterobeltiosis to best parent in cross (BPHB%) for all types of crosses in year 1999 |
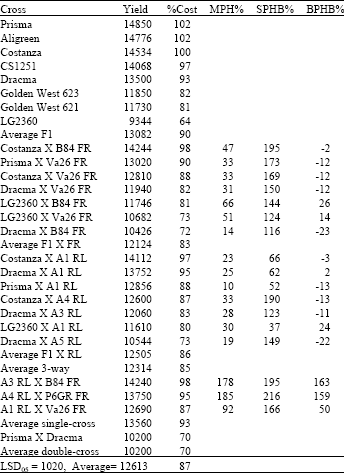 | |
The F2 generation of Costanza reached 55% of original hybrid, while the average F2 performance of Costanza and Prisma reached 52% of Costanza F1 and 57% of average performance between Costanza F1 and Prisma F1 (12610 kg ha-1). The average yield of F1 found to be 12842 kg ha-1.
The average of 3-way crosses found to be 10950 kg ha-1.
| Table 4: | Yield in kg ha-1, relative yield as percentage of F1 hybrid Costanza (%Cost), Middle-parent Heterosis (MPH%), Heterobeltiosis to second parent in cross (SPHB%) and Heterobeltiosis to best parent in cross (BPHB%) for all types of crosses in year 2000 |
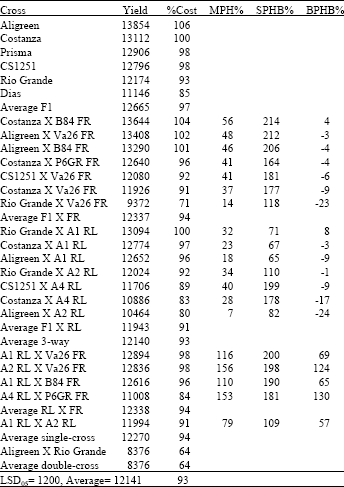 | |
Three-way crosses between F1 and RL lines were better performing when compared to 3-way between F1 and FR lines. Single crosses (in average) yielded close to the average performance of the three-way crosses and close to the performance of the double cross Prisma X Costanza.
Similar data are presented in Table 3-6, where the average performance of F1 was always better than the average of all other type of crosses. In general, three-way crosses between F1 and RL lines were better performing when compared to 3-way between F1 and FR lines (Table 2-6). A few experimental single crosses found to be better than F1 hybrids (the average in year 1999, Table 3). Various three-way crosses yielded near or better than the corresponding F1 hybrid, exhibiting high Heterobeltiosis to second parent (around 200%) and satisfactory Middle-parent Heterosis (over 50%) and in some cases Heterobeltiosis to F1 parent.
The Coefficient of Variation (%) is presented in Table 7. This includes crosses with average CV across years <10%. Costanza X Va26 and Marista X Va26 exhibited the lower Cvs, that is only 3% in average (1% for Costanza X Va26 in 1998).
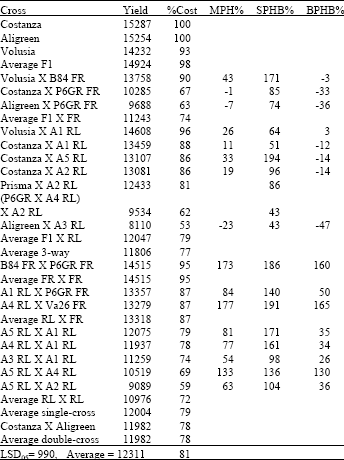
| Table 5: | Yield in kg ha-1, relative yield as percentage of F1 hybrid Costanza (%Cost), Middle-parent Heterosis (MPH%), Heterobeltiosis to second parent in cross (SPHB%) and Heterobeltiosis to best parent in cross (BPHB%) for all types of crosses in year 2001 |
 | |
| Table 6: | Yield in kg ha-1, relative yield as percentage of F1 hybrid Costanza (%Cost), Middle-parent Heterosis (MPH%), Heterobeltiosis to second parent in cross (SPHB%) and Heterobeltiosis to best parent in cross (BPHB%) for all types of crosses in year 2002 |
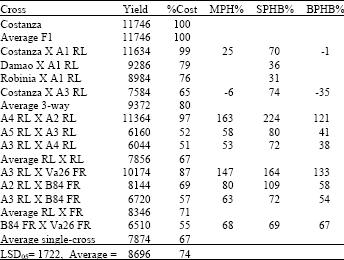 | |
Costanza F1 had a CV = 9% in average (Table 7).
Combining data from results, three-way crosses between Costanza X A1 were high yielding (94.756% of Costanza F1 yield) with a low CV (near 6%) and the same was found for Rio Grande X A1 (90.22% of Costanza F1 yield).
| Table 7: | The Coefficient of Variation % as a parameter of stability for years: 1998 (CV98), 1999 (CV99), 2000 (CV00), 2001 (CV01) and average CV (CV Avg), only for crosses with average CV<10% |
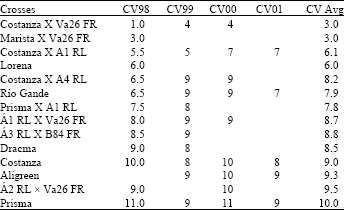 | |
These three-way crosses to the best performing inbred line A1 were always in the top of ranking and the average yield was at 89.5% of Costanza F1. In general, the average yield for all three-way crosses was at 82.1% of Costanza F1. Three-way crosses to RL lines exhibited better performance in comparison to FR lines (83.15% against 81.05% of Costanza F1). Costanza and Rio Grande were crossed successfully to RL lines (87.25% and 85.186% of Costanza F1 in average, respectively), but only Costanza was crossed successfully to FR lines (89.95% of Costanza F1 in average). Prisma and Dracma crossed satisfactory to RL lines (near 84% of Costanza F1) and Aligreen to FR lines (near 89% of Costanza F1). Costanza X Va26 (average yield 87.97% of Costanza F1) and Costanza X B84 (101% of Costanza F1, Middle-parent Hetrosis around 50%) three-way crosses were successful. The average yield of three-way crosses to inbred line B84 was found at 91% of Costanza F1.
DISCUSSION
It is expected, high yielding inbred lines used in crosses to express favorable additive gene action when considering yielding performance per se or in crosses where the level of heterozygosity is low[5,9,11,19]. Another consequence of this fact is that hybrids formed by crossing high yielding inbred lines may depend a portion of their vigor on favorable additive gene action, since such kind of loci may be alike in the two parents especially when they derived from common pedigree[5,9,19,20].
The average of the F2 evaluated in 1988 were the expected one by the theory[10,13], near 55% of the respective F1 yielding performance. Three-way crosses, single crosses and double crosses were found to be at the same level of productivity and in general, three-way crosses between F1s and RL lines were better performing when compared to 3-way between F1s and FR lines. Various three-way crosses yielded near or better than the corresponding F1 hybrid, exhibiting high heterobeltiosis to second parent and satisfactory Middle-parent heterosis and in some cases Heterobeltiosis to F1 parent. These findings indicate that, new combinations formatting three-way crosses could exploit both favorable additive and dominant gene action because of the lower level of heterozygosity in comparison to F1 and the high level of productivity of the inbred parents[5,8,9,11-14].
The use of three-way commercial hybrids is a low-cost technique for seed companies[13] but the lack of genetic uniformity may lead in competition effects and unstable yielding performance[9]. Fasoulas[9,21], Fasoula and Fasoula[22-24] proposed selection of parents in absence of competition and evaluation in many places from the first stages of breeding and of course evaluation of the final crosses in various environments to achieve stability of yielding performance. Fasoulas[9,21] , Fasoula and Fasoula[22-24] consider that Coefficient of Variation (CV) is a meter of uniformity and stability. From this point of view, the low CVs of a number of three-way crosses indicate uniformity and stability. Additionally, many three-way crosses exhibited high and stable yields across years of evaluation. Visual observation revealed satisfactory uniformity within plots.
Since a number of three-way crosses exhibited high yields accompanied by uniformity and stability, depending their performance on the productivity and stability of the two parents, it seems to be a means for exploiting heterosis under low-cost hybrid production programs. F1 hybrid Costanza proved to be a good mother parent for development of three-way crosses with high and stable yielding performance. The RL inbred line A1 was combined easily to F1 hybrids to produce high and stable three-way crosses. It is clear that this line contributed useful additive gene action, which usually leads to stable performance[5,9,11]. The B84 inbred line exhibited considerable specific combining ability, in some cases and Va26 inbred line contributed in stable performance.
The expected Middle-parent Heterosis (MPH%) in three-way crosses involving recombinant lines (RL), may be computed by the equation; MPH= 76.166+0.411Y, where, Y is the relative yield of the cross in comparison to Costanza F1 (r= 0.62, p<0.001). Heterobeltiosis to the best parent (BPHB%) may be computed by the equation; BPHB= 90.516+0.544Y, where, Y is the relative yield of the cross (r= 0.73, p<0.001). No relation between Heterobeltiosis to inbred line parent (SPHB%) and three-way cross performance was found, indicating different combining performance of RL inbred lines.
The expected Middle-parent Heterosis (MPH%) in three-way crosses involving free-release inbred lines (FR), may be computed by the equation; MPH= 27.296+0.515Y, where, Y is the relative yield of the cross in comparison to Costanza F1 (r= 0.77, p<0.001). Heterobeltiosis to the best parent (BPHB%) may be computed by the equation; BPHB= 88.654+0.504Y, where, Y is the relative yield of the cross (r= 0.57, p<0.005). Heterobeltiosis to the inbred line parent (SPHB%) may be computed by the equation; SPHB= 37.891+0.305Y, where, Y is the relative yield of the cross (r= 0.98, p<0.001). In general, heterotic phenomena were alike in the two kinds of three-way crosses. This is not in agreement with that found for single crosses[24] and may be attributed in heterotic gene action contributed by the hybrid parent.
As a conclusion, the development of three-way crosses by using modern maize hybrids, may be an easy method to exploit heterosis and achieve high and stable yielding performance. The cost of this method is considerably low since the parents used in this study (F1 hybrids and inbred lines) are highly productive. In such cases, high and stable yields may depend on both useful additive and dominant gene action.
REFERENCES
- Ipsilandis, C.G. and M. Koutsika-Sotiriou, 2000. The combining ability of recombinant S-lines developed from an F2 maize population. J. Agric. Sci. Cambridge, 134: 191-198.
Direct Link - Fehr, W.R., 1987. Principles of Cultivar Development. Vol. 1, Macmillan Publishing Co., New York.
Direct Link - Koutsika-Sotiriou, M., I. Bos and A. Fasoulas, 1990. Hybrid reconstruction in maize. Euphytica, 45: 257-266.
Direct Link - Genter, C.F. and M.W. Alexander, 1966. Development and selection of productive S1 inbred lines of corn (Zea mays L.). Crop Sci., 6: 429-431.
Direct Link - Koutsika-Sotiriou, M. and I. Bos, 1996. Heterosis after several numbers of cycles of mass honeycomb selection in maize. J. Genet. Plant Breed., 50: 7-13.
Direct Link - Fasoula, D.A. and V.A. Fasoula, 1997. Competitive ability and plant breeding. Plant Breed. Rev., 14: 89-138.
Direct Link - Fasoula, D.A. and V.A. Fasoula, 1997. Gene action and plant breeding. Plant Breed. Rev., 15: 315-374.
Direct Link - Fasoula, V.A. and D.A. Fasoula, 1999. Honeycomb breeding: Principles and applications. Plant Breed. Rev., 18: 177-250.
Direct Link