
A. Soltani
Not Available
B. Torabi
Not Available
E. Zeinali
Not Available
R. Sarparast
Not Available
Asian Journal of Plant Sciences
Year: 2004 | Volume: 3 | Issue: 6 | Page No.: 705-708







ABSTRACT
Chickpea (Cicer arietinum L.) is considered to have a quantitative long-day response to photoperiod. No day-neutrality or qualitative response to photoperiod has been reported. The objective of this study was to report qualitative response of chickpea to photoperiod. Four chickpea genotypes from different geographical areas of Iran were serially sown in 11 sowing dates during 2001 to 2003. Plants sown on August, September, October, November and January sowing dates flowered at about May 10 (day of year 130). Flowering was delayed up to 120-150 days when day length decreased to about 11-12 h, indicating that plants remained unresponsive until photoperiod increased above the critical value. It was concluded that chickpea can also be classified as a qualitative long-day plant.
PDF Abstract XML References Citation
How to cite this article
A. Soltani, B. Torabi, E. Zeinali and R. Sarparast, 2004. Response of Chickpea to Photoperiod as a Qualitative Long-day Plant. Asian Journal of Plant Sciences, 3: 705-708.
DOI: 10.3923/ajps.2004.705.708
URL: https://scialert.net/abstract/?doi=ajps.2004.705.708
DOI: 10.3923/ajps.2004.705.708
URL: https://scialert.net/abstract/?doi=ajps.2004.705.708
INTRODUCTION
Three main categories of photoperiodic response are recognized in plants[1]: photoperiod-insensitive or Day-neutral Plants (DNPs), Short-day Plants (SDPs) and Long-day Plants (LDPs). SDPs and LDPs types are further divided into two groups: one is absolute or qualitative response group with critical photoperiod above (SDPs) or below (LDPs) which plants do not flower; and the quantitative or facultative response group of which flowering is accelerated either by short day or long day but with no distinct critical photoperiod.
Chickpea (Cicer arietinum L.) is an important legume crop. It is grown in ecologically diverse environments in India, the Mediterranean, eastern Africa, the Americas, Europe and Australia[2] and is a source of high quality protein in food and feed. Chickpea is generally considered to be a quantitative long-day plant[3-10]. So far no day-neutrality or qualitative long-day response have been reported in chickpea. The objective of this study was to report qualitative behavior in response of chickpea to photoperiod that has not been reported before.
MATERIALS AND METHODS
A serially sown field experiment was conducted at the Gorgan University of Agricultural Sciences Research Farm, Gorgan (36° 85’ N, 54° 27’ E and 100 m asl), Iran. The experiment started in December 2001 and continued until August 2003. Four kabuli chickpea genotypes (Beauvanij, 9096c, Hashem and Jam) were sown at 11 sowing dates. These genotypes selected from different geographical areas across Iran; 9096c and Hashem are cultivated in the west part of the Caspian Sea Coast of Iran (Golestan province), Jam in the north-west (Azerbaijan province) and Beauvanij in the mid west of the country (Kermanshah and Kordestan provinces). Sowing dates (day of year, DOY) were 12 December (346) 2001 and 15 January (15), 15 February (46), 17 March (76), 16 April (106), 18 May (138), 17 June (168), 16 August (228), 15 September (258), 14 October (287) and 12 November (316) 2002. The soil was a deep silty clay and the experiment was conducted under well-watered conditions. Therefore, there was no effect of flooding or water deficit stresses.
The experimental design was a single split plot with sowing dates in the main plot and genotypes in the sub plot, replicated four times. Plot size was 1.5 m by 4 m. Plots were hand-seeded using row spacing of 25 cm. Target plant density was 50 plant m-2 and two seeds were planted at correct spacing at 4 cm depth and seedlings thinned to one in each position later. Stages of development of emergence (50% of plants with some parts at soil surface), flowering (50% of plants with one flower at any node, R1), pod initiation (50% of plants with 0.5 cm pod at one of the 4 upper nodes with unrolled leaf, R3), pod filling (50% of plants with peas beginning to develop, R5) and maturity (50% of plants with pods yellowing, R7) were recorded every 2 days. During the experiment weeds were hand-controlled and several sprayings were carried out against Ascochyta blight. Hashem and 9096c are resistant to the blight, but Beauvanij and Jam are susceptible and were badly affected. At the end, there were 9 sowing dates for Hashem and 9096c with reliable data and this number was 7 for Beauvanij and 6 for Jam. This study, focussed on flowering data of 9096c and Hashem.
To separate effect of photoperiod and temperature on time to flowering, we also calculated thermal time and examined its relationship to photoperiod. Daily thermal time (DTT, °C d) was computed as:
| (1) |
where, f is a scalar factor that accounts for sub-and supra-optimal temperatures on development rate and Tb and To1 are the base and the lower optimum temperatures for development, respectively. f was calculated as:
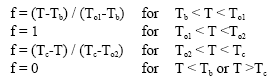 | (2) |
where, T is the average daily temperature and To2 and Tc are the upper optimum and ceiling temperatures for development. All the temperatures are in °C. Tb, To1, To2 and Tc were taken as 1, 22.6, 32.6 and 39.0°C, respectively (Soltani and Torabi, unpublished data). Thermal time from emergence to flowering was calculated by accumulating DTT after emergence till flowering. Daily maximum and minimum temperatures were measured at a weather station a few meters far from the experimental units.
RESULTS AND DISCUSSION
Temperature and photoperiod changes during the experiment are shown in Fig. 1. Temperatures ranged between 5.2 to 40.4°C for maximum temperature and between -3.4 to 27.2°C for minimum temperature. Day length varied between 10.2 and 15.3 h per day. A strong qualitative photoperiod response will be evident in serial sowings of a genotype if photoperiod changes over the response range[11]. For Hashem, days to flowering ranged between 44 and 232 days. This range was between 45 and 225 days for Cv. 9096c (Table 1). In a qualitative long-day response, plants flower only when photoperiod is higher than a critical value. A qualitative long-day response was found for the evaluated cultivars. In Hashem, as an example, flowering occurred at about DOY 130 for sowing dates of September, October, November, December and January, although these sowing dates differed by 120 days (Table 1 and Fig. 1a). It should be noted that plants in sowing date of August 2002 did not flower until nearly DOY 50 when they died as influenced by Ascochyta blight.
| Table 1: | Time to flowering for Cvs. 9096c and Hahsem at different sowing dates. Mean temperature (°C) from emergence to flowering is also included. |
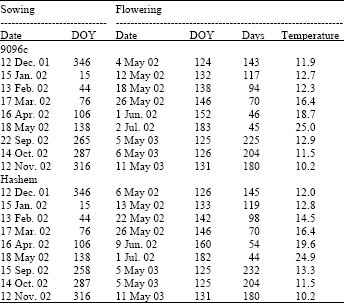 | |
| DOY = Days of year | |
Examination of sowing dates of September, October and November 2002 indicated that with delay in sowing date, days to flowering declined along with decrease in temperature (mean temperature of 13, 12 and 10°C, respectively). Clearly, at these sowing dates plants remained unresponsive until photoperiod increased above the critical value.
Figure 2 shows days to flowering and thermal time from emergence to flowering plotted versus day length for the four cultivars evaluated. With photoperiod of about 11 to 12 h per day, days to flowering and thermal time increased suddenly, indicating day length of 11-12 h per day as a probable critical photoperiod. Response curve of flowering time to photoperiod in this study is similar to the curve of qualitative long-day plants as presented in various texts[1, 12].
The present results contradict the general conclusion that, “chickpea is a quantitative long-day plant but flowers in every photoperiod”[7]. In an early study on chickpea development, the effect of eight photoperiods from 9 to 16 h on flowering of two cultivars, Vilmorin and Lyallpur, was studied[3]. Both cultivars flowered in all photoperiods, suggesting that there was no critical day length. Photoperiod of 9 h delayed flowering of Vilmorin and Lyallpur by 35 and 20 days, respectively, compared to photoperiod of 16 h. Similarly, it was found that, in controlled environments, the Cv. Chafa flowered sooner (27 days) in 16 h and day/night temperatures of 33/18°C than in 11 h days and 22/10°C temperatures (48 days)[4]. Newly, it has been shown that pigeonpea can be classified as a qualitative short-day plant[11], while previously considered to be a quantitative short-day plant[1].
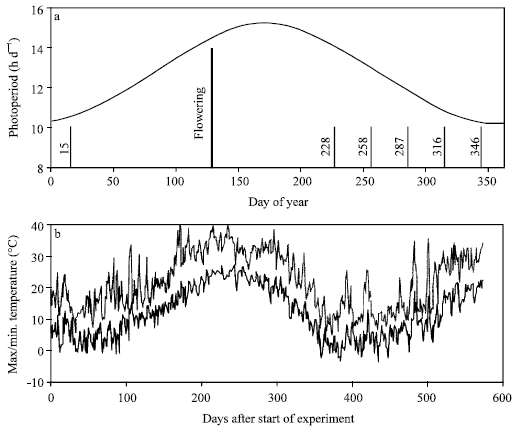 | |
| Fig. 1: | Changes of photoperiod during the year (a) and maximum and minimum temperatures during the experiment (b) at Gorgan. In (a), short lines and their numbers (DOY) indicate some sowing dates and the thick line indicates the time of flowering for these sowing dates |
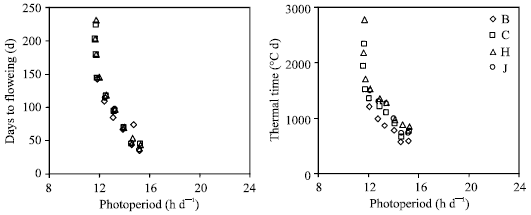 | |
| Fig. 2: | Days from sowing to flowering and thermal time from emergence to flowering for four chickpea cultivars as a function of photoperiod. Cultivars are Beauvanij (B), 9096c (C), Hashem (H) and Jam (J). Each point is mean of four replications |
It has been reported that in some cultivars of chickpea short days vernalization may substitute for the natural long day requirement[13]. Present results do not support this in the evaluated cultivars. At sowing date of September, October, November, December and January, there have been suitable temperatures for vernalization, but they did not substitute for need to long days. Present results also indicate that response to cold (vernalization) is quantitative rather than qualitative in agreement with a previous report[14]. For instance, at sowing date of May 2002, temperatures were not suitable (mean 25°C) for vernalization, but the cultivars flowered in the shortest time.
Overall, the data indicate that chickpea can also be considered as a qualitative long-day plant, in which flowering does not take place at photoperiod lower than a critical value. We are analyzing the data to obtain cardinal temperatures and photoperiods for phenological development of chickpea.