
Zahid Pervaiz
Not Available
Mohammad Afzal
Not Available
Sun Xi
Not Available
Yang Xiaoe
Not Available
Luo Ancheng
Not Available
Asian Journal of Plant Sciences
Year: 2002 | Volume: 1 | Issue: 4 | Page No.: 478-481







ABSTRACT
A solution culture experiment was conducted in wire house of the Department of Soil Science and Agricultural Chemistry, Zhejiang Agricultural University, Hangzhou, P.R.China with natural daylight and day/night temperature of 17/7 oC respectively with the objective to correlate the results of different physiological parameters with salt tolerance of cultivars under 100 and 200 mol m -3 NaCl salinity. The two Chinese cultivars used were Bao-119, CV.86-6 and one Pakistani was FSD-85. The results of this study indicate, that with the gradual increase of salinity dry weight of shoot and root, potassium, calcium contents both in shoot and root and leaf area decreased, where as sodium, chloride and chlorophyll contents increased in all the cultivars except chlorophyll contents decreased at 100 mol m -3 salinity in Bao-119. Photosynthesis increased at 100 mol m -3 salinity and decreased at 200 mol m -3 salinity in all cultivars. The tolerance of cultivars on cumulative dry weight means (3 treatments) was in order of CV.86-6 > FSD-85 > Bao-119 whereas at 200 mol m -3 salinity was as FSD-85 > CV.86-6 > Bao-119. The cultivars FSD-85 and CV.86-6 showed salt inclusion and exclusion mechanisms respectively at 200 mol m -3 salinity in relation to ionic composition. Consistent pattern was found between relative leaf area and dry weight of shoot at 200 mol m -3 salinity and on cumulative mean (3 treatments) basis. In-consistent pattern was found among chlorophyll contents, photosynthesis rates and tolerance of cultivars at both the salinity levels. From this study it was also concluded, that before the detail study of mechanisms of salt tolerance, the following considerations are also essential for the authenticity of the proposed tolerant and sensitive cultivars. The cultivars under test must grow under uniform conditions and must receive their own optimum requirements, and sampling for analysis of different parameters must be done after the same duration of stress and preferably from same plant parts. In the absence of the above, it is difficult to correlate the results obtained from analysis of different physiological parameters.
PDF Abstract XML References Citation
How to cite this article
Zahid Pervaiz, Mohammad Afzal, Sun Xi, Yang Xiaoe and Luo Ancheng, 2002. Physiological Parameters of Salt Tolerance in Wheat. Asian Journal of Plant Sciences, 1: 478-481.
DOI: 10.3923/ajps.2002.478.481
URL: https://scialert.net/abstract/?doi=ajps.2002.478.481
DOI: 10.3923/ajps.2002.478.481
URL: https://scialert.net/abstract/?doi=ajps.2002.478.481
This technology gives economic returns from salt affected soils and provide vegetative covers to soil which reduces evaporation and hence the rate of salinization (Qureshi and Barrett-Lennard, 1998). Study of response of plants/crops to salinity under naturally saline condition is not feasible due to extreme variability in soil salinity both spatially and temporarily (Richards, 1983). To avoid this problem, comparative differences for salt tolerance among crops/ varieties can be studied under artificially salinized control conditions.
Salinity tolerance in wheat has been and is being extensively researched in Pakistan and elsewhere in the world, but still efforts to improve salt tolerance have been hampered by a number of factors, particularly the lack of understanding of the mechanisms of salt tolerance and interaction of salinity with various environmental factors with regard to plant growth. Wheat tolerance to salinity varies with the stage of plant growth, nature and level of salinity, duration of stress etc., (Qureshi et al., 1990) and is affected by soil moisture, climate, nutrition and management practices (Maas and Hoffman, 1977). Different physiological traits such as selectivity for potassium, exclusion and/or compartmentation of sodium and chloride ions, an osmotic adjustment by accumulation of organic solutes have all been related to salt tolerance of crops plants (Wyn Jones and Storey, 1981).
In this study an attempt has been made to study the relationship between yield, ionic contents, leaf area, chlorophyll contents, photosynthesis and salt tolerance of cultivars under consideration.
MATERIALS AND METHODS
Experiment was conducted in wire house of the Department of Soil Science and Agricultural Chemistry, Zhejiang Agricultural University, Hangzhou, P.R.China during 18th November, 1991 to 4th January, 1992, with natural daylight and day/night temperature of 17/7 0C respectively. Sufficient healthy seeds of three wheat cultivars {2 Chinese (Ba0-119, CV.86-6) and 1 Pakistani (FSD-85)} were soaked in 0.2% fungicide solution for 18 hours. After draining fungicide solution, the seed were washed thrice with water. Then seeds were sown in quartz sand in iron trays. The condition in trays kept moist with water and trays remained covered until the sprout came out and waited for nine days. Thirteen days old 20 seedlings of each cultivars were transferred to 1 cm plugged holes in wooden covers over 32 liters of 2 strength Hoagland nutrient solution (Hoagland et al., 1950) in plastic containers. Ten holes were used for each cultivars and each hole having 2 seedlings. Fifteen days old seedlings were subjected to incremental salt stress.
Salt concentrations were increased by 25 mol m-3 after every 12 hours by adding NaCl to nutrient solution until the required salinity levels (100, 200 mol m-3) were obtained in respective containers. Thirty-eight days old seedlings were subjected to full strength Hoagland nutrient solution. Solutions were renewed after every 7 days and pH 6.0-6.5 was daily maintained and loss of water was made regularly. Solutions were aerated for 9 hours every day with air pump by splitting in to 3 equal parts and intervals. Twelve plants were harvested 30 days after salinization. The plants were washed for five minutes in running tap water followed by a quick rinse in distilled water. The plants tissue dried at 70 °C were weight and ground in mortar with pestle and stored in polyethylene bags and redried at the time of weighing for analysis. The ground plant materials were digested with 1N HCl for 24 hours at 40 °C, then the required volume was shaked for one and half-hours and filtered. In the digest, sodium, potassium and calcium were determined by “ICP” model, Jarrel-Ash, ICPA-9000. Chloride was determined by Cl electrode method. Statistical analysis was done by the method given by Steel and Torrie (1980). Leaf area was measured after 32 days of salinization by portable area meter, model, LI. 3000. Li-COR, U.S.A. Photosynthesis after 36 days with the help of LI-6200 Portable photosynthesis system. (LI-COR, INC. U.S.A.). Chlorophyll contents were determined after 34 days by using the method of Cheng Fuming and Cheng Shengwei (1984).
Reagents: Mixture solution (extractant for chlorophyll) = Acetone (4.5 ml)+ Ethyl alcohol absolute (4.5ml)+ Distilled water (1ml).
Small pieces (0.1 gm) of homogenized fresh leaf were taken in 20 ml test tube. Added 10 ml mixture solution. Led the tube and put in absolute dark condition till leaves become white. After leaves turned to white colour, chlorophyll a and b were determined at 645 and 663 nm, respectively with photometer.
| 1 | Chlorophyll “a” mg dm-2 | = | 12.7 x D663 B 2.69 x D645 x |
| V / 1000 x wt | |||
| 2 | Total chlorophyll mg dm-2 | = 20.2 x D645 + 8.42 x | |
| D663 x V / 1000 x wt | |||
RESULTS AND DISCUSSION
The results of this study indicate, that with the gradual increase of NaCl salinity, gradual decrease in yield of shoot and root, K+ and Ca++ contents both in shoot and root and leaf area of all the cultivars was observed, whereas Na, Cl¯ and chlorophyll contents increased in all the cultivars except decrease of chlorophyll in Bao-119 at 100 mol m-3 salinity. Photosynthesis was increased at 100 mol m-3 where as decreased at 200 mol m-3 salinity. Data in Table 1 revealed that cumulative mean (3 cultivars) shoot dry yields were 89% and 62% of control at 100 and 200 mol m-3 salinity respectively. At 100 mol m-3 salinity CV.86-6 gave the maximum (95 % of control) but at 200 mol m-3 salinity, FSD-85 produced maximum relative yield (67% of control). On the basis of cumulative means absolute shoot yield, CV.86-6 gave the better yield (1.27 gm) followed by FSD-85 (1.19 gm) and Bao-119(1.01 gm), but trend of relative yield was different at both the salinity levels. It was also observed that the cultivars selected from screening experiments changed their behavior in relation to tolerance. It was postulated that this change was mainly due to the differences in duration of salt stress, concentration of media and environment conditions such as light, temperature etc. As concerned the root yield, CV.86-6 gave the better root yield (100% and 63% of control) at both the salinity levels as compared to other cultivars, while on cumulative mean basis, dry weight of roots indicated, that CV.86-6 had the least and FSD-85 had the better root yield.
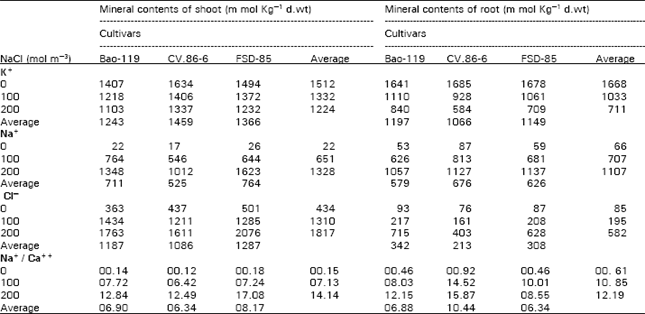
The K+ contents of shoot showed a marked decreased with the increase of salinity both in shoots and roots (Table 2). Highly significant differences were observed among treatments and cultivars. The CV.86-6 showed the maximum cumulative mean of three treatments (1459 m mol Kg-1 d.wt) and Bao-119 had lowest K+ contents in its shoots (1243 m mol Kg-1 d.wt). Trend of K+ contents in shoot had positive correlation (r= 0.990) with dry cumulative mean of absolute shoot yield. In roots, CV.86-6 kept the minimum K+ at both salinity levels, while Ba0-119 had the maximum K+ contents. It indicated that low yield of Bao-119 was due to the retention of high K+ contents in roots as compared to other cultivars, opposite trend was found in CV.86-6. Roots of all cultivars had lower K+ than shoots. It appears that most of the absorbed K+ was translocated to the shoot. As regard Na+ contents of shoot, CV.86-6 had the lowest Na+ contents in its shoot at both the salinity levels as compared to other cultivars. In roots CV.86-6 had the highest Na+contents and was followed by FSD-85 and Bao-119. Maintaining maximum K+ and minimum Na+ contents in shoot of CV. 86-6 was the main mechanism of salt tolerance and cultivars can be said “salt excluder”. Keeping high Na+ in the shoot of FSD-85 and giving high absolute and relative dry yield at 200 mol m+ salinity was also the main mechanisms of salt tolerance in this cultivar and can be said as “salt includer” because it has the capability to produce high yield under adverse conditions as compared to the other cultivars. Less shoot yield of Bao-119 was due to keeping more Na+ contents in its shoot, which disturbed osmotic adjustment of this cultivars. Inconsistent pattern were found while compared shoot and root Na+ contents. As regard Cl¯ contents of shoot, the trend was same as was in case of Na+ contents of shoot, but in case of roots, CV.86-6 had least cumulative means (3 treatments) Cl ¯contents and Bao-119 had maximum. Minimum Cl¯ contents in shoot and root of CV.86-6 and maximum in Bao-119 caused the increase and decrease of shoot yield of these cultivars respectively. It seemed that Cl¯ toxicity had also influenced the growth of shoot and root. The cultivars FSD-85 also had highest Cl¯ contents in its shoot and even then it produced the maximum absolute and relative dry yield at 200 mol m-3 salinity. This was might be due to its genetic character and nature of maintaining high salt for osmotic adjustment under high saline condition by absorbing higher quantity of salt. Sodium calcium ratios were also less in case of shoot of CV.86-6 and highest in FSD-85, which is other evidence in relation to their tolerance.
In saline environment where salts are present in higher concentrations, plant growth is affected negatively in various ways such as osmotic effects, specific ion effect and nutritional imbalance; probably all occurring simultaneously (Flowers et al., 1991). Initial growth inhibition in saline environment is induced by the decreased water potential of rooting medium due to higher salt concentration (Munn et al., 1995). A secondary effect of high concentrations of Na+ and Cl- in the root medium is the suppression of uptake of essential nutrients such as K+, Ca++, NO3 etc. (Gorham and Wyn Jones, 1993). The decrease in shoot and root yield of all the cultivars with the addition of salt could be due to the reduction of physiology availability of water with the increase in solute suction from saline media or accumulation of toxic ions in plants. With the gradual increase of NaCl salinity, decrease in shoot and root yield, potassium and calcium contents and increase in sodium and chloride contents is in agreement with those of Roth (1989), Sharma (1989), Ehert et al. (1990), Brugnoli and Lauter (1991), Saneoka et al. (1999) and Akhtar et al. (2001).
The leaf area was progressively decreased with the increase of salinity (Table 3). Similar results were reported by Yaseen (1987), Gupta (1989) and Saneoka et al. (1999). The reduction in leaf area was probably due to decrease in cell size (Peter et al., 1987) and due to the decrease in the number and volume of cells in barley crops (Yaseen, 1987). On cumulative means basis, leaf area was highest in case of CV. 86-6 and was followed by FSD-85 and Bao-119 and at both salinity levels, FSD-85 had the highest leaf area. Inconsistent and consistent pattern was found at 100 mol m-3 and 200 mol m-3 NaCl concentration respectively with dry weight in relation to their tolerance but on cumulative means basis, non-significant correlation (r= 0.926) was found with dry weight of all cultivars. The cultivars Bao-119 and CV.86-6 showed highly significant differences among all treatments, whereas FSD-85 showed this difference only between saline treatments. Chlorophyll contents were progressively increased with the increase of salinity except in case of Bao-119 at 100 mol m-3 NaCl concentration that gave 93% of control. On cumulative means basis, chlorophyll was highest in Bao-119 and was followed by FSD-85 and CV.86-6. Nonsignificant negative correlation (r= - 0.995) was found between chlorophyll contents and leaf area. Among cultivars at both salinity levels in-consistent pattern was found between chlorophyll contents and tolerance of cultivars. There is very few data available on the increase of chlorophyll contents. The exact reason of increase of chlorophyll contents is not clear, but this increase might be due to the decrease in the activity of enzyme chlorophylls. The increase in chlorophyll under saline condition was reported by Passera and Albuzio (1978) in durum wheat and Reddy (1986) in bajra. Photosynthesis contents were increased at 100 mol m-3 and decreased at 200 mol m-3 salinity in all the cultivars and this is in agreement with those of Passera and Albuzio (1978). At 100 mol m-3 salinity, Bao-119 gave the maximum photosynthesis rate and was followed by CV.86-6 and FSD-85, but at 200 mol m-3 salinity CV.86-6 had the better photosynthesis rate.
| Table 1: | Effect of sodium chloride salinity on dry weight of shoot and root of three cultivars |
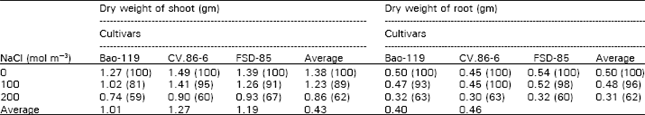 | |
| Values in parenthesis are percentage of their respective control. | |
| Table 2: | Effect of sodium chloride salinity on ionic compositions of shoot and root of three wheat cultivars |
 | |
| Table 3: | Effect of sodium chloride salinity on leaf area, chlorophyll contents and photosynthesis of three wheat cultivars |
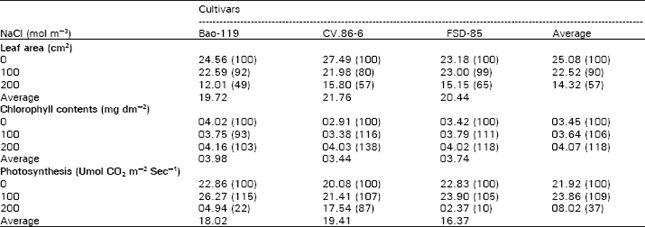 | |
| Values in parenthesis are percentage of their respective control | |
Generally on cumulative mean basis, CV.86-6 gave the highest photosynthesis rate and was followed by Bao-119 and FSD-85. Photosynthesis rate had negative correlation with Na+ and Cl¯ contents in shoots of all the cultivars. It seemed that accumulation of Na+ and Cl¯ in shoots affected the water status of plant and disturbed turgor pressure. Abnormal concentrations of Na+ and Cl¯ in the cytoplasm have been associated with the inhibition of photosynthesis and stomatal closure (Hasson et al., 1983 and Rawson, 1986). Higher photosynthesis rate in CV.86-6 at 200 mol m-3 salinity as compared to other cultivars was probably due to the maintenance of higher turgor due to higher turgor potential in its leaves, (Uprety and Sirohi, 1985). Ba0-119 and FSD-85 showed significant differences while CV.86-6 showed non-significant difference at both salinity levels.
The findings of this study indicate, that the cultivars selected from screening experiment, changed their behavior in relation to salt tolerance. It is postulated that this change was mainly due to the differences in duration of salt stress, concentration of media and environmental conditions such as light, temperature etc. Therefore, it is suggested that before the detail study of mechanism of salt tolerance, the following consideration are very essential for the authenticity of the proposed tolerant and sensitive cultivars. The cultivars under test must grow under uniform conditions and must receive their on optimum requirements such as temperature, time of sowing etc. Long duration and short duration should preferably be tested separately. Screening should be carried out at the critical as well as other ages of growth and conditions used for selection should be as close to the field condition as possible. From this study it was also concluded, that if any meaningful study on mechanism of salt tolerance has to be done, sampling for analysis of different parameters must be done after the same duration of stress period and preferably from same plant parts. In the absence of the above, it is difficult to correlate the results obtained from analysis of different parameters. Much more work needs to be done on this subject in order to improve the results.
REFERENCES
- Brugnoli, B. and M. Lauter, 1991. Effect of salinity on stomatal conductance, phtosythetic capacity and carbon isotope discrimination of salt tolerant cotton and Phaseolus vulgaris. Plant Physiol., 95: 628-635.
Direct Link - Ehret, D.L., R.E. Redman, B.L. Harvey and A. Cipywnyk, 1990. Salinity induced calcium deficiencies in wheat and barley. Plant Soil, 128: 143-151.
Direct Link - Flowers, T.J., M.A. Hajibagheri and A.R. Yeo, 1991. Ion accumulation in the cell wall of rice plants growing under saline conditions: Evidence for Oertli hypothesis. Plant Cell Environ., 14: 319-325.
Direct Link - Hasson, E., A. Poljakoff-Mayber and J. Gale, 1983. The Effect of Salt Species and Concentrations on Photosynthesis and Growth of Pea Plants (Pisum sativum L. CV. Alaska). In: Effects of Stress on Photosynthesis, Marcelle, R., H. Clijsters and M. van Poucke (Eds.). W. Junk Publishers, The Hague, The Netherlands.
- Maas, E.V. and G.J. Hoffman, 1977. Crop salt tolerance-current assessment. J. Irrig. Drainage Div., 103: 115-134.
Direct Link - Munns, R., D.P. Schachtman and A.G. Condon, 1995. The significance of a two-phase growth response to salinity in wheat and barley. Aust. J. Plant Physiol., 22: 561-569.
Direct Link - Passera, C. and A. Albuzio, 1978. Effect of salinity on photosynthesis and photorespiration of two wheat species (Triticum durum CV. PEPE 2122 and Triticum aestivum CV. Marzotto). Can. J. Bot., 56: 121-126.
Direct Link - Rawson, H.M., 1986. Gas exchange and growth in wheat and barley grown in salt. Aust. J. Plant Physiol., 13: 475-489.
Direct Link - Reddy, M.P. and A.B. Vora, 1986. Changes in pigment composition, hill reaction activity and saccharides metabolism in bajra (Pennisetum typhoides S and H) leaves under NaCl salinity. Photosynthetica, 20: 50-55.
Direct Link - Richards, R.A., 1983. Should selection for yield in saline conditions be made on saline or non-saline soils? Euphytica, 32: 431-438.
CrossRefDirect Link - Saneoka, H., K. Shiota, H. Kurban, M. Chaudhary, G.S. Premachandra and K. Fujita, 1999. Effect of salinity on growth and solute accumulation in two wheat lines differing in salt tolerance. Soil Sci. Plant Nut., 45: 873-880.
Direct Link