
Mumtaz Ali Sethar
Not Available
Vajanti Mala Pahoja
Not Available
Qamaruddin Chachar
Not Available
Asian Journal of Plant Sciences
Year: 2002 | Volume: 1 | Issue: 3 | Page No.: 261-263







ABSTRACT
The research work was conducted to elucidate the heat acclimation potential of two promising cotton cultivars Rehmani and S-12 . The plants were hardened for 1 to 7 days at 43ºC for 2h. After every hardening treatment the leaves were heat stressed at 45ºC for 2h. The chlorophyll fluorescence (Fv/Fm) ratio was recorded at pre-incubation, during heat stress and recovery after heat stress. The results showed that cv. Rehmani gave better results only after 1 day hardening treatment but after 2 to 7 days treatments the Fv/Fm ratio declined. However, cultivar S-12 showed good acclimation response as the treatment days were increased to 7-days hardening treatment the Fv/Fm ratio increased to 83% of the control treatment (no hardening).
PDF Abstract XML References Citation
How to cite this article
Mumtaz Ali Sethar, Vajanti Mala Pahoja and Qamaruddin Chachar, 2002. Photosynthetic Acclimation of Cotton Genotypes at Higher Temperatures. Asian Journal of Plant Sciences, 1: 261-263.
DOI: 10.3923/ajps.2002.261.263
URL: https://scialert.net/abstract/?doi=ajps.2002.261.263
DOI: 10.3923/ajps.2002.261.263
URL: https://scialert.net/abstract/?doi=ajps.2002.261.263
INTRODUCTION
The summers of Pakistan can produce exceptionally high temperatures (45-50°C). Under these conditions, even summer crops including cotton will have great difficulty in maintaining photosynthetic activity and growth. Maintenance of metabolic balance under such conditions requires that the plant should possess a high photosynthetic acclimation potential. Burke et al. (1976) have stated that some species acclimate extensively in response to environmental stress while others acclimate only a small extent, and some species do not acclimate at all. One of the main factors determining the ability of plants to adapt to different growth temperatures is their ability to modify their membrane constituents, in order to function normally under the new conditions. Heat damage to biomembranes in these circumstances may be prevented by the synthesis and accumulation of protective compounds in or near to the membranes.
Many workers have reported that various water soluble compounds such as sugars and proteins are able to protect sensitive cell structures against heat inactivation (Santarius, 1973; Kraus and Santarius, 1975; Santarius and Muller, 1979). Feldman and Ageeva (1973) found that hardening increased the thermostability of membrane bound (photosynthetic) ferredoxin in pea plants. It has been suggested (Quinn and Williams, 1985; Terzaghi et al., 1989) that high temperature adaptation occurs due to alteration in the lipid and /or protein composition of the photosynthetic membranes. Muller and Santarius (1979) also suggested that increase in thermostability depends on ultrastructural changes in the thylakoid membrane, rather than biochemical changes in membrane composition. Taguchi (1984) concluded that the seeds of wheat sown in field some times suffer dehydration, and the emergence and establishment were unstable varying with the soil water condition. Even if the seed germinates, the seedling becomes coiled in the soil, and were unable to emerge. In this context Andoh and Kobata (2000) stated that seed hardening of rice is subjection of seeds to wetting and redrying before sowing increased the germinability and/or yields under low soil-moisture conditions. Furthermore, Andoh and Kobata (2001) revealed that seed hardening, imbibition followed by redrying, accelerated seed germination and seedling emergence of wheat in desiccated soils.
The objectives of investigation was to reveal the effectiveness of short temperature treatments on the hardening capabilities of two promising Pakistani cotton cultivars at sub-lethal high temperatures.
MATERIALS AND METHODS
The cotton seed of cultivars Rehmani and S-12 was obtained from cotton section A.R.I. Tandojam (Pakistan). The research work was undertaken at School of Biological Sciences, University of Wales, Bangor (UK) during 1992-93. The seeds were germinated in a temperature and light controlled incubator (Fitotran 600) with 30/27°C day/night and 16/8h light/dark period. After about 8-12 days of sowing the leaves were transferred for hardening treatments to another incubator set at 43°C for a period of 2h in the dark. The plants were then returned to the growth cabinet at 30°C. Transfers between growth cabinet and incubator were made using an insulated box to avoid light and unwanted temperature fluctuations. These hardening treatments were arranged so that the plants were not older than 16 days at the end of the hardening treatments. Finally, leaves from the heat hardened plants were harvested and subjected to heat stress for 2h at 45°C before, being allowed to recover for up to 24h at 30°C. The stress temperature of 45°C was selected because at this temperature the photosynthetic activities were almost ceased in these cultivars (Sethar et al., 2001). Chlorophyll fluorescence technique was used to estimate the changes in leaves during the experiment. The fluorescence was recorded at preincubation (30 min at 30°C), during the temperature stress (45°C) and at recovery of 2, 6, 12, 24h at 30°C after stress. The Fv/Fm ratio is reported because this ratio indicates the oxygen evolution in the photosynthesis process. The procedure of chlorophyll fluorescence measurement has been mentioned some where else (Sethar et al., 1995).
RESULTS AND DISCUSSION
The results from cultivars Rehmani and S-12 are presented in Tables 1 and 2 and Fig. 1 and 2. The fluorescence data are presented as the percentage of the Fv/Fm values recorded at the end of the dark pre-incubation period.
| Table 1: | Effect of whole plant hardening on leaf fluorescence in cv. Rehmani. |
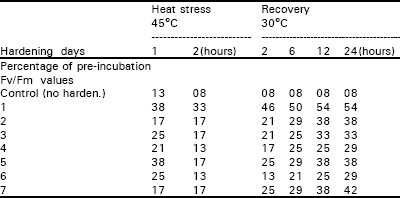 | |
| Table 2: | Effect of whole plant hardening on leaf fluorescence in cv. S-12. |
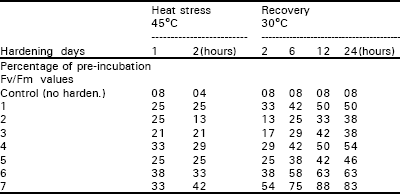 | |
The results in Table 1 showed that cv. Rehmani displayed an improved heat resistance following one day of the hardening treatment. The Fv/Fm values decreased to only 38 and 33% of the pre-incubation values at the end of 1 and 2h, respectively of heat stress in comparison to decreases of 13 to 8% in the control. The fluorescence values then increased to 46% after 2h of recovery and continued to increase to 50% after 6h of recovery.
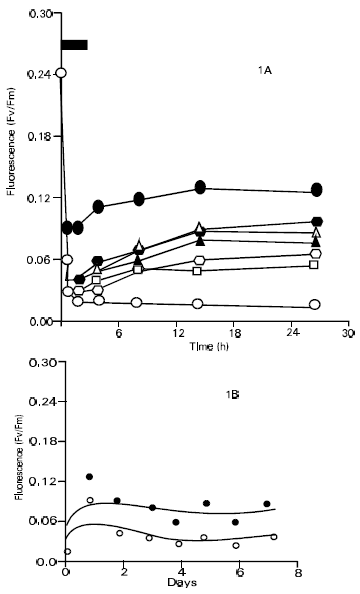 | |
| Fig. 1: | Effect of hardening of leaf fluorescence in cv. Rehamani 1a, time course of fluorescence during and after heat hardening for 0 to 7 days. ● , 1 day; Δ, 2 Days ▲ , 3 Days; □ , 4 Days; ■ , 5 Days |
After 12h of recovery the Fv/Fm value remained unchanged up to the end of the 24h recovery period. Further hardening treatments, in which leaves were hardened between 2 and 7 days, produced markedly lower fluorescence ratios compared with the 1-day treatment. In the 2-7 days hardening treatments the Fv/Fm ratio decreased to 13-17% of the pre-incubation value at the end of 2h heat stress period. During the recovery period the Fv/Fm value increased slightly to 25-38% of the pre-incubation value after 12h. There was little change in these values after 24h recovery. These results revealed that cv. Rehmani has limited capability of hardening under the particular conditions used in this study. Table 2 showed that cultivar S-12 behaved differently from cv. Rehmani. Following a single hardening treatment the leaves had lost about 75% of their fluorescence at the end of the 2h heat stress period. The value then gradually increased, however, to 50% of the pre-incubation value after 12h recovery. The 2 and 3 day treatments showed a poorer performance compared with 1-day treatment. Leaves which had been hardened over 4 or 5 days produced patterns of response which were more similar to that recorded for the 1-day treatment.
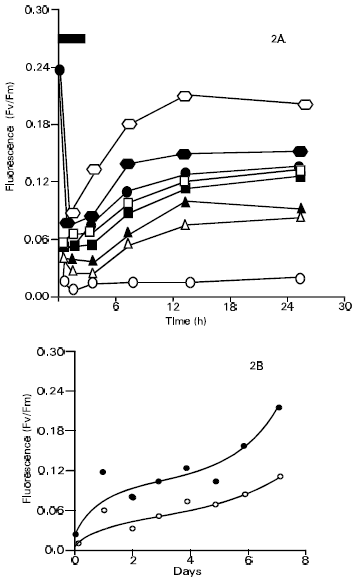 | |
| Fig. 2: | Effect of hardening of leaf fluorescence in cv. S-12 2a, time course of fluorescence during and after heat hardening for 0 to 7 days. ● , 1 day; Δ, 2 Days ▲ , 3 Days; □ , 4 Days; ■ , 5 Days |
It displayed markedly better results after 6 days of hardening treatments, however, where it retained about one third of its starting fluorescence after 2h of heat stress and it regained about two third of its fluorescence after 12h recovery. The 7-day hardening treatment was even more effective in preventing heat injury, which shows that 7 days hardened leaves retained more than 40% of their initial fluorescence at the end of 2h of heat stress. After the heat stress, the recovery in Fv/Fm values was very good reaching 75% after 6h of recovery and 88% after 12h of recovery. There was however, a slight decrease in the Fv/Fm value after 24h reaching 83% of the pre-incubation value. The Fv/Fm data are presented graphically in Fig. 1a and 2a which shows more clearly the Fv/Fm ratio decrease during the course of the 2h heat stress. This trend was similar in both cultivars. Thus, during the first hour of heat stress the fluorescence ratio dropped sharply and it continued to decline more slowly during the second hour of stress. When the leaves were put back to 30°C for recovery the Fv/Fm values tended to increase for all the hardening treatments. This was in contrast to the control leaves (no hardening) where both the cultivars showed negligible recovery in their Fv/Fm values. To summarize, the results shown in Fig. 1a and 2a, derivative plots were prepared using the Fv/Fm values from 2h. heat stress and from 12hr in the recovery period. These derivative plots are presented in Fig. 1b and 2b, where the Fv/Fm values for each hardening treatment are plotted against the number of hardening days. The Fig. 1b and 2b display clearly the differences in the results obtained for both cultivars. Cultivar Rehmani displayed a poor performance except for 1-day hardening treatment. Both curves in the Fig. 1b showed a progressive declining trend after 1 day. Cultivar S-12 performed very well although the points in Fig. 2b display a considerable scatter. The curves nevertheless showed that an increasing number of hardening treatments increased the plants tolerance to heat stress. The response was greatest following 7 days of hardening treatment. Experiments reported in this research work, the general appearance of the leaves also gave valuable information regarding the effects of hardening. In the leaves given no hardening treatment, about 90-95% of them turned brown either fully or partially following heat stress. Little or no browning was noted in the heat hardened leaves.
In a study (Sether et al., 1995) where the investigation into suitable hardening temperature was carried out, the cotton plants which were acclimated only 1-2°C below or above the hardening range of 42-43°C performed very poorly. These findings are interesting in the light of a similar report by Trione and Metzger (1970) working on cold acclimation of wheat seedlings, that in the hardening temperature range of 5-8°C the seedling acclimated well, but they were unable to elongate their meristems when acclimated either at 3 or 9°C . Hardening treatments repeated over a number of days gave markedly better tolerance than the control treatment (unhardened leaves) in this study, and differences were detected between cultivars. In cv. Rehmani the recovery in leaf fluorescence after heat stress was found to be significantly lower than that in cv. S-12, where cv. Rehmani recovered only about 50% of the Fv/Fm ratio while cv. S-12 regained 83% of Fv/Fm values of the control. This indicates that the hardening process was not working at its full capacity in cv. Rehmani. Significant metabolic changes other than those associated with photosynthesis may also occur during the process of hardening in plants. Several workers, Yelenosky (1979), Lalk and Dorffing (1985) and Sobczyk et al. (1985) noted increases in proline, Adenosine tri phosphate, ABA, glutamic acid and ascorbic acid levels, and in the proportion of unsaturated fatty acids in membrane lipids hardened plants at low temperatures compared to those of no-hardened plants. Similarly, Windle (1988) reported an increase in the degree of fatty acid unsaturation in the phospholipids, phosphatidylcholine and phoshatidylethanolamine in rye plant hardened to high temperatures.
The acclimation of a plant at sub-lethal temperature may also induce the synthesis of heat shock proteins which protect the plant from the potentially damaging effects of the later and more severe high temperature stress (Howarth, 1991). Woledge et al. (1989) reported that temperature acclimation may have increased the canopy photosynthesis of rye grass and white clover. There is evidence regarding the acclimation of photosynthesis at low temperature, that although temperature has a well known direct effect on the rate of leaf photosynthesis so that the light saturated rate in rye grass and white clover grown at 15°C is reduced by about half when measurements were made at 5°C. There was some acclimation, in that the amount by which photosynthetic rate was decreased by low temperature was less in plants which have been grown at low temperature (Woledge and Dennis, 1982; Nosberger et al., 1983).
The research work was carried out to reveal the heat acclimation potential of cotton cultivars Rehmani and S-12. The plants were hardened for 1 to 7 days at 43°C. The chlorophyll fluorescence technique was used to estimate the changes in leaves. After every hardening treatment the leaves were heat stressed at 45°C for 2h. The results showed that cv. Rehmani was better only after 1 day hardening treatment, but the fluorescence ratio declined in 2-7 days hardening treatments. Cultivar S-12 however, showed good hardening potential as the treatment days increased.
REFERENCES
- Andoh, H. and T. Kobata, 2000. Does wetting and redrying the seed before sowing improve rice germination and emergence under low soil conditions?. Plant Prod. Sci., 3: 161-163.
Direct Link - Andoh, H. and T. Kobata, 2001. Effect of seed hardening, wetting and redrying before sowing, on germination and seedling emergence of a japanese wheat variety norin 61 in desiccated soil. Plant Prod. Sci., 4: 50-55.
Direct Link - Burke, M.J., L.V. Gusta, H.A. Quamme, C.J. Weister and P.H. Li, 1976. Freezing injury in plants. Ann. Rev. Plant Physiol., 27: 507-528.
Direct Link - Howarth, C.J., 1991. Molecular responses of plants to an increased incidence of heat shock. Plant Cell Environ., 14: 831-841.
Direct Link - Krause, G.H. and K.A. Santarius, 1975. Relative thermostability of the chloroplast envelope. Planta, 127: 285-299.
CrossRefDirect Link - Lalk, I. and K. Dorffing, 1985. Hardening, abscisic acid, proline and freezing resistance in two wheat varieties. Physiol. Plant., 63: 287-292.
Direct Link - Muller, M. and K.A. Santarius, 1979. Changes in chloroplast membrane lipids during adaptation to barley to extreme salinity. Plant Physiol., 62: 326-329.
PubMedDirect Link - Santarius, K.A., 1973. The protective effects of sugars on chloroplast membrane during temperature and water stress and its relationship to frost, desiccation and heat resistance. Planta, 113: 105-114.
CrossRefDirect Link - Santarius, K.A. and M. Muller, 1979. Investigations on heat resistance on spinach leaves. Planta, 146: 529-538.
CrossRefDirect Link - Sethar, M.A., V.M. Pahoja, Q.I. Chachar and D.L. Laidman, 2001. Impact of optimal and superoptimal temperatures on the photosynthetic apparatus of cotton leaves. J. Biol. Sci., 1: 1088-1092.
Direct Link - Sobczyk, E.A., A. Marszaleka and A. Kacperska, 1985. ATP involvement in plant tissue responses to low temperature. Physiol. Plant., 63: 399-405.
Direct Link - Terzaghi, W.B., D.C. Fork, J.A. Berry and C.B. Field, 1989. Low and high temperature limits to PS-II. Plant Physiol., 91: 1494-1500.
Direct Link - Trione, E.J. and R.J. Metzger, 1970. Wheat and barley vernalization in a precise temperature gradient. Crop. Sci., 10: 390-392.
Direct Link - Windle, J.J., 1988. A comparative spin label study of isolated plasma membrane of whole cells and protoplasts from cold hardened and no hardened winter rye. Plant Physiol., 88: 1388-1396.
PubMedDirect Link - Woledge, J. and W.D. Dennis, 1982. The effect of temperature on the photosynthesis of rye grass and white clover leaves. Ann. Bot., 50: 25-35.
Direct Link - Woledge, J., I.A. Davidson and V. Tewson, 1989. Photosynthesis during winter in rye grass/white clover mixtures in the field. New Phytol., 113: 275-281.
Direct Link - Yelenosky, G., 1979. Accumulation of free proline in citrus leaves during cold hardening of young trees in controlled temperature regimes. Plant Physiol., 64: 425-427.
PubMedDirect Link