
Cuneyt Cirak
Department of Agronomy, Faculty of Agriculture, University of Ondokuz Mayis, Samsun, Turkey
American Journal of Plant Physiology
Year: 2007 | Volume: 2 | Issue: 5 | Page No.: 287-294







ABSTRACT
The availability of wild plants from different Hypericum species in Turkish flora is strongly limited due to uncontrolled harvesting. Thus, seed germination requirements of the four rare Hypericum species, namely H. orientale, H. perfoliatum, H. pruinatum and H. origanifolium were studied by performing some pre-soaking treatments with the aim of describing suitable germination protocols for use in ex situ conservation. Before placing the seeds in Petri dishes, they were soaked in 50, 100 or 150 mg L-1 GA; 0.5, 1 or 1.5% H2SO4; 150 mg L-1 GA + 0.5% H2SO4, 0.01 solutions, tap water, 40, 50 or 60°C hot water for 30 min. The study was performed under a photoperiod of 18 h light/8 h darkness in growth chambers. In H. orientale and H. origanifolium, the highest germination was induced by H2SO4 1.5% in both species (81 and 52%, respectively) while GA 100 and 150 mg L-1 treatments produced the highest germination in H. perfoliatum (43%) and H. pruinatum (80%) seeds. The results indicated that propagation from seed is a viable method for the ex situ conservation of these species and germination response of the seeds to the pre-soaking treatments is variable. The changeable germination responses were discussed as a possible result of the dormancy involving partially dormant embryo for H. perfoliatum and H. pruinatum and hard seed coat for H. orientale and H. origanifolium.
PDF Abstract XML References Citation
How to cite this article
Cuneyt Cirak, 2007. Seed Germination Protocols for Ex situ Conservation of Some Hypericum species from Turkey. American Journal of Plant Physiology, 2: 287-294.
DOI: 10.3923/ajpp.2007.287.294
URL: https://scialert.net/abstract/?doi=ajpp.2007.287.294
DOI: 10.3923/ajpp.2007.287.294
URL: https://scialert.net/abstract/?doi=ajpp.2007.287.294
INTRODUCTION
The conservation of genetic resources of medicinal plants has been of increasing interest and seed storage is the simplest and most economical method for ex situ conservation of plant genetic resources (Perez-Garcia et al., 2006). However, each species has particular requirements for seed germination as a result of adaptive radiation into patchy and changing environments (Cerabolini et al., 2004). Germination is a critical stage in the life cycle of weeds and crop plants and often controls population dynamics, with major practical implications (Keller and Kollmann, 1999). Generally, seed germination is restricted by different kind of dormancy in many plant species. Plant growth regulators such as GA (gibberellic acid) and IAA (indoleacetic acid) (Hilhorst and Karssen, 1992; Iglesias and Babiano, 1997); chemicals such as sulphuric acid (H2SO4) and hot water treatments (Tomer and Maguire, 1989; Baes et al., 2002) have been recommended to break dormancy and enhance germination.
Hypericum is a large genus of herbs or shrubs which grow in temperate regions of the world (Patocka, 2003). Species belonging to this genus have been used as traditional medicinal plants due to their wound-healing, bactericide, anti-inflammatory, diuretic and sedative properties for the last two hundred years (Dias et al., 2000). There are about 400 species of Hypericum in relatively dry temperate zones of the world. In Turkey, the genus is represented by 89 species of which 43 are endemic (Davis, 1988).
Today, some Hypericum species such as H. perforatum (Kirakosyan et al., 2004), H. brasiliense (Abreu et al., 2003), H. androsaemum (Valentao et al., 2003), H. perfoliatum (Couladis et al., 2001) and H. scabrum (Matsuhisa et al., 2002) have become a valuable commodity to wild-crafters who supply these species for the herbal industry due to their phytomedicinal properties and the market for only H. perforatum has exceeded $570 million worldwide annually (Sirvent et al., 2002). The increased market demand for Hypericum derived products has led to uncontrolled harvesting of plant materials from wild populations. In Turkey, wild-crafters and some companies have collected Hypericum plants excessively for export to herbal industry. As a result, the availability of wild plants from different Hypericum species in Turkish flora is strongly limited. Thus, the present study focuses on the determination of ex situ requirements for seed germination in the four Hypericum species, namely H. orientale L., H. perfoliatum L., H. pruinatum Boiss. and Bal. and H. origanifolium Willd. by performing some pre-soaking treatments to promote germination as an initial step in their conservation.
MATERIALS AND METHODS
Brief Description of Plant Material
Hypericum orientale: Stems 7-45 cm, erect or decumbent and sometimes rooting. Leaves 10-40 mm, oblong or elliptic-oblong to oblanceolate or linear. Sepals unequal, narrowly oblong and ovate or broadly elliptic to obovate. Petals 10-18 mm, entire, without black dotes. Capsule 7-14 mm, ovoid to ovoid-cyclindric. Hypericum pruinatum: Stems 15-35 cm, erect or ascending from a rooting and branching base, pruinose. Leaves on main stems 10-35 mm, oblong to elliptic, pruinaose. Inflorescence pyramidal to cyclindric. Sepals broadly oblong to broadly elliptic, rounded, entire or minutely black-glandular-denticulate. Petals 9-14 mm. Capsule 7-10 mm, ovoid. Hypericum perfoliatum: Stems 15-80 cm, erect or decumbent. Leaves 13-60 mm, ovate to triangular-lanceolate or rarely linear, the upper most sometimes black-glandular-ciliate, usually densely pellucid-dotted, with obscure reticulate venation. Sepals oblong, subacute to rounded, densely and irregularly dark-glandular-denticulate. Petals 9-14 mm, with or without black dotes. Capsule 5-6 mm, broadly ovoid, with dorsal vittae and lateral vesicles. Hypericum origanifolium: Stems 5-30 cm, suberect to ascending, branching but not usually rooting at the base. Leaves 5-30 mm, ovate or obovate with intramarginal and sometimes superficial black dots. Sepals narrowly oblong to oblong-spathulate, usually with some superficial black dots. Petals 9-15 mm, with black dots. Capsule 7-12 mm, with dorsal vittae and lateral vesicles (Fig. 1) (Davis, 1988). The four Turkish species of Hypericum grow wild in Black Sea region of Turkey at high altitudes. The plant materials were identified by Dr. Hasan Korkmaz, Department of Biology, University of 19 Mayis, Samsun, Turkey. Voucher specimens were deposited in the herbarium of Ondokuz Mayis University Agricultural Faculty (OMUZF # 55 for H. orientale, OMUZF # 107 for H. pruinatum, OMUZF # 109 for H. origanifolium and OMUZF # 101 for H. perfoliatum).
Seed Collection
The seeds were handpicked from at least ten randomly selected individuals during October of 2005 and stored at 4±2°C in sealed plastic bags until used for germination tests.
Experimental Procedures
The present study was conducted at laboratory of Ondokuz Mayis University, Agricultural Faculty. In preliminary testing, seeds placed in Petri dishes did not germinate effectively under normal laboratory conditions. The pre-soaking treatments used in the study were different GA and H2SO4 doses, hot water and tap water. Before placing the seeds in Petri dishes, they were soaked in 50, 100 or 150 mg L-1 GA; 0.5, 1 or 1.5% H2SO4 solutions, tap water, 40, 50 or 60°C hot water for 30 min.
 | |
| Fig. 1: | H. perfoliatum (a), H. pruinatum (b), H. origanifolium (c) and H. orientale (d) plants at full flowering |
Seeds also were soaked in 150 mg L-1 GA and then 0.5% H2SO4 solutions for 30 min to evaluate double effect of these chemicals (Cirak et al., 2004a). The treated seeds were placed in individual, sterilised Petri dishes containing moisture-retaining paper liners. Paper liners in the Petri dishes were kept moist throughout the germination period. The study was performed under a photoperiod of 18 h light/8 h darkness in growth chambers. Temperature was set at 20°C, recommended temperature for germination in H. brasiliense and H. perforatum seeds (Bertelle et al., 2004). Germination was measured as a percentage, 20 days after the experiment was initiated. The seeds showing radicle emergence were recorded as germinated (Come, 1970).
Data Analysis
The experimental design was a factorial randomized block arrangement with three replications with 100 seeds in each. Germination percentages from the original data were transformed for statistical analysis (arcsine of square root of percent germination x 0.01). The transformed data were analyzed using ANOVA and differences among treatments were tested using the Duncan Multiple Range Test (level of significance p<0.01) (Yurtsever, 1984).
RESULTS
According to the results of variance analysis (Fig. 2), the pre-soaking treatments tested had a significant (p<0.01) effect on germination rates depending on species.
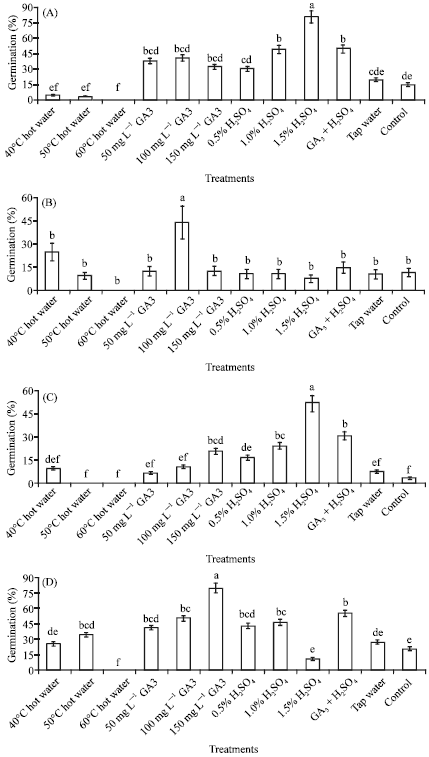 | |
| Fig. 2: | The germination rates of H. orientale (A) H. perfoliatum (B) H. origanifolium (C) and H. pruinatum (D) seeds exposed to different pre-soaking treatments. (Values with different letters within columns for each species differ significantly at the level of p<0.01; bars are±SE) |
In H. orientale and H. origanifolium, H2SO4 treatment was found to be most effective to improve seed germination depending on doses while other treatments were efficient to a lesser degree. The highest germination was induced by H2SO4 1.5% in both species (81 and 52%, respectively), followed by GA + H2SO4 and H2SO4 1% (50 and 49% for H. orientale, 30 and 24% for H. origanifolium, respectively). It is interesting to note that hot water treatments in increasing levels deteriorated germination when compared to control in both species.
GA treatment was the only effective one in promoting germination for H. perfoliatum seeds and GA 100 mg L-1 resulted in the highest germination (43%). The other treatments were all generally ineffective in improving germination and were found in the same statistical group as control.
Germination rates elevated linearly with GA treatment in increasing doses in H. pruinatum seeds. The highest germination was induced by GA 150 mg L-1 (80%), followed by GA + H2SO4 (55%) and GA 100 mg L-1 (50%). On the contrary, germination was suppressed by hot water 60°C and H2SO4 10% treatments.
DISCUSSION
Light has been recognized since the mid-nineteenth century as a germination-controlling factor and it is frequently found to be a requirement in plant species native to arid lands (Baskin, 2004). In general, absence of light has a negative effect on germination in several Hypericum species such as H. perforatum (Campbell, 1985), H. gramineum (Ash et al., 1998), H. brasiliense (Bertelle et al., 2004) and H. aviculariifolium (Cirak et al., 2007). In a previous study, it was found that 18/6 h light/dark cycle was the most effective to meet light requirement for germination in H. perforatum seeds (Cirak et al., 2004b). Thus, the present study was performed under this photoperiod to supply the probable light requirement.
Seeds of many wild members of different genus have hard seed coats which restrict water absorption by the embryo. Failure to imbibe limits O2 to the embryo and leaching of inhibitors, therein effectively enforcing dormancy of the embryo. For applied uses, dormancy-breaking treatments are required to provide more uniform and rapid seed germination responses. Permeability may be improved by scarifying the seed coat by mechanical means (e.g., clipping, abrasion or immersion in hot water) or chemically with strong oxidative agents (e.g., sulphuric acid or sodium hypochlorite) (Abdallah et al., 1989). In the present study, H2SO4 treatment was found to be most effective to induce seed germination in H. orientale and H. origanifolium. The results indicate the presence of physical dormancy related to hard seed coat and overcame by only acid scarification. The high germination rates obtained with H2SO4 treatments agree with those obtained from other species of Hypericum such as H. lydum and H. tetrapterum (Cirak et al., 2006) as well as some legumes (Teketay, 1996; Grouzis and Danthu, 2001), Prosopis ferox (Baes et al., 2002), Acacia origena, Acacia pilispina and Pterolobium stellatum (Teketay, 1998), Erythrina brucei and Erythrina burana (Teketay, 1994).
Studies of genetics and physiology have shown the important roles of the plant hormones abscisic acid and gibberellin in the regulation of dormancy and germination (Koornneef et al., 2002). Gibberellins comprise the class of hormones most directly implicated in the control and promotion of seed germination. Endogenously applied gibberellins can relieve certain types of dormancy, including physiological dormancy, photodormancy and thermodormancy acting as a substitute for low temperatures, long days, or red light (Seiller, 1998). In this study, GA increased germination rate significantly, depending on concentration in H. perfoliatum and H. pruinatum indicating the presence of physiological dormancy related to partially dormant embryo. Similar results were obtained from studies carried out on other species, such as Sesamum indicum (Kyauk et al., 1995), Rumex dentatus (Ali and Helal, 1996), Zea mays and Glycine max (Wang et al., 1996), Opuntia tomentosa (Carrillo et al., 2003) and Physoplexis comosa (Cerabolini et al., 2004).
Hot water treatments have been reported to enhance germination of hard coated seeds by elevating water and O2 permeability of the testa (Teketay, 1998; Aydin and Uzun, 2001). In present case, hot water treatment in the lowest level induced germination in H. perfoliatum, H. origanifolium and H. pruinatum while it was not effective in promoting germination in H. orientale. The negative effect of the thermal pretreatments on H. orientale seeds was probably due to the combination of high temperature and time, which may cause a damage to the embryo tissue as observed in several species (Khosh-Khui and Bassiri, 1976; Masamba, 1994).
Chemicals that accumulate in the fruit and seed-coat during development and remain in the seed after harvest can act as germination inhibitors. Some of the substances associated with inhibition are various phenols, coumarin and abscisic acid which can be leached out by soaking in water (Booth and Sowa, 2001). In case of H. perforatum and H. aviculariifolium, soaking the seeds in tap water resulted in a significant increase in germination (Cirak et al., 2004c and 2007). In the present study, similarly, tap water treatment slightly increased germination rates in H. orientale and H. pruinatum seeds, but this increase was found to be insignificant.
CONCLUSIONS
Seed germination behavior is an integral part of ex situ conservation, especially for developing standard viability monitoring protocols and to ensure sufficient populations for germplasm regeneration. However, germination requirements for native species are often unknown. Here, it is the first time we have described the seed germination requirements of H. orientale, H. perfoliatum, H. origanifolium and H. pruinatum, which are under threat due to uncontrolled harvesting together with other species of Hypericum from Turkish flora. According to the results, propagation from seed is a viable method for the ex situ conservation of these species. However, it should be noted that seed storage and germination are only the first steps in the reinforcement of populations of these species. Further studies based on field experiments to attain baseline data on ex situ plant development and establishment are currently underway.
ACKNOWLEDGMENT
Author is grateful to Dr. Dursun Ali Tokel, Ondouz Mayis University, Faculty of Science and Art, for his valuable comments and technical assistance.
REFERENCES
- Abreu, I.N., M.T.A. Azevedo, V.M. Solferini and P. Mazzafera, 2003. In vitro propagation and isozyme polymorphism of the medicinal plant Hypericum brasiliense. Biol. Planta., 47: 629-632.
CrossRefDirect Link - Al-Helal, A.A., 1996. Studies on germination of Rumex dentatus L. seeds. J. Arid Environ., 33: 39-47.
CrossRefDirect Link - Ash, J.E., R.H. Groves and A.J. Willis, 1998. Seed ecology of Hypericum gramineum, an Australian forb. Aust. J. Bot., 45: 1009-1022.
Direct Link - Aydin, I. and F. Uzun, 2001. The effects of some applications on germination rate of gelemen clover seeds gathered from natural vegetation in Samsun. Pak. J. Biol. Sci., 4: 181-183.
Direct Link - Baes, P.O. M.L. de Viana and S. Suhring, 2002. Germination in Prosopis ferox seeds, effects of mechanical, chemical and biological scarificators. J. Arid Environ., 1: 185-189.
Direct Link - Baskin, J.M. and C.C. Baskin, 2004. A classification system for seed dormancy. Seed Sci. Res., 14: 1-16.
CrossRefDirect Link - Bertelle, F.M.L., P.M. Beatriz and L.A. Augusto, 2004. Light, temperature and potassium nitrate in the germination of Hypericum perforatum L. and H. Brasiliense choisy seeds. Bragantia, 63: 193-199.
Direct Link - Booth, D.T. and S. Sowa, 2001. Respiration in dormant and non-dormant bitterbrush seeds. J. Arid Environ., 48: 35-39.
CrossRefDirect Link - Campbell, M.H., 1985. Germination, emergence and seedling growth of Hypericum perforatum. Weed Res., 25: 259-266.
CrossRefDirect Link - Carrillo, Y.O., J.M.B. Guzman, S.C. Victor, O. Esther and S. Alma, 2003. Germination of the hard seed coated Opuntia tomentosa, a cacti from the Mexico valley. J. Arid Environ., 55: 29-42.
Direct Link - Cerabolini, B., A. Rossella, M. Roberta, S.P. Ceriani and R. Barbara, 2004. Seed germination and conservation of Endangered species from the Italian Alps, Physoplexis comosa and Primula glaucescens. Biol. Conserv., 117: 351-356.
Direct Link - Cirak, C., K. Kevsero-lu and B. Sa-lam, 2004. Physical and physiological dormancy in black henbane (Hyoscyamus niger L.) seeds. J. Plant Biol., 47: 391-395.
Direct Link - Cirak, C., A. Ayan, K. Kevseroglu and O. Caliskan, 2004. Germination rate of St. John's wort (Hypericum perforatum L.) seeds exposed to different light intensities and illumination periods. J. Biol. Sci., 4: 279-282.
CrossRefDirect Link - Cirak, C., A.K. Ali and K. Kudret, 2004. The effects of light and some presoaking treatments on germination rate of St. John's worth (Hypericum perforatum L.) seeds. Pak. J. Biol. Sci., 7: 182-186.
CrossRefDirect Link - Cirak, C., K. Kevseroglu and A.K. Ayan, 2006. Breaking of seed dormancy in a Turkish endemic Hypericum species: Hypericum aviculariifolium subsp. depilatum var. depilatum by light and some pre-soaking treatments. J. Arid Environ., 68: 159-164.
Direct Link - Couladis, M., P. Baziou, P.V. Petrakis and C. Harvala, 2001. Essential oil composition of Hypericum perfoliatum L. growing in different locations in Greece. Flavour Fragrance J., 16: 204-206.
Direct Link - Dias, A.C.P., R.M. Seabra, P.B. Andrade, F. Ferreres and M. Fernandes-Ferreira, 2000. Xanthone biosynthesis and accumulation in calli and suspended cells of Hypericum androsaemum. Plant Sci., 150: 93-101.
Direct Link - Sy, A., M. Grouzis and P. Danthu, 2001. Seed germination of seven Sahelian legume species. J. Arid Environ., 49: 875-882.
CrossRefDirect Link - Hilhorst, H.W.M. and C.M. Karssen, 1992. Seed dormancy and germination, the role of abscisic acid and gibberellins and the importance of hormone mutants. Plant Growth Regul., 11: 225-238.
Direct Link - Iglesias, R.G. and M.J. Babiano, 1997. Endogenous abscisic acid during the germination of chickpea seed. Physiol. Plant., 100: 500-504.
Direct Link - Keller, M. and J. Kollmann, 1999. Effects of seed provenance on germination of herbs for agricultural compensation sites. Agric. Ecosyst. Environ., 72: 87-99.
CrossRefDirect Link - Khosh-Khui, M. and A. Bassiri, 1976. Physical dormancy in myrtle seed. Scientia Hortic., 5: 363-366.
CrossRefDirect Link - Kirakosyan, A., T.M. Sirvent, D.M. Gibson and P.B. Kaufman, 2004. The production of hypericins and hyperforin by in vitro cultures of St. John's wort (Hypericum perfotarum). Biotechnol. Applied Biochem., 39: 71-81.
Direct Link - Kyauk, H., N.W. Hopper and R.D. Brigham, 1995. Effects of temperature and pre-soaking on germination, root length and shoot length of sesame (Sesamum indicum L.). Environ. Exp. Bott., 35: 345-351.
Direct Link - Masamba, C., 1994. Presowing seed treatments on four African Acacia species, appropriate technology for use in forestry for rural development. For. Ecol. Manage., 64: 105-109.
Direct Link - Matsuhisa, M., Y. Shikishima, Y. Takaishi, G. Honda and M. Ito et al., 2002. Benzoylphloroglucinol derivatives from Hypericum scabrum. J. Nat. Prod., 65: 290-294.
Direct Link - Patocka, J., 2003. The chemistry, pharmacology and toxicology of the biologically active constituents of the herb Hypericum perforatum L. J. Applied Biomed., 1: 61-70.
Direct Link - Perez-Garcia, F., M. Huertas, E. Mora, B. Pena, F. Varela and M.E. Gonzalez-Benito, 2006. Hypericum perforatum L. seed germination: Interpopulation variation and effect of light, temperature, presowing treatments and seed desiccation. Gen. Res. Crop Evol., 53: 3387-3398.
Direct Link - Seiller, G.J., 1998. Seed maturity, storage time and temperature and media treatment effects on germination of two wild sunflowers. Agron. J., 90: 221-226.
Direct Link - Sirvent, T., L. Walker, N. Vance and G. Donna, 2002. Variation in hypericins from wild populations of Hypericum perforatum L. in the Pasific Northwest of the USA. Econ. Bot., 56: 41-48.
CrossRefDirect Link - Teketay, D., 1994. Germination ecology of two endemic multipurpose species of Erythrina from Ethiopia. For. Ecol. Manage., 65: 81-87.
CrossRefDirect Link - Teketay, D., 1996. Germination ecology of twelve indigenous and eight exotic multipurpose Leguminous species from Ethiopia. For. Ecol. Manage., 80: 209-223.
Direct Link - Teketay, D., 1998. Germination of Acacia origena, A. pilispina and Pterolobium stellatum in response to different pre-sowing seed treatments, temperature and light. J. Arid Environ., 38: 551-560.
CrossRefDirect Link - Valentao, P., A. Dias, M. Ferreira, B. Silva, P.B. Andrade, M.L. Bastos and R.M. Seabra, 2003. Variability in phenolic composition of Hypericum androsaemum. Nat. Prod. Res., 17: 135-140.
Direct Link - Wang, Q., Z. Feng and D. Smith, 1996. Application of GA and kinetin to improve corn and soybean seedling emergence at low temperature. Environ. Exp. Bot., 36: 377-383.
Direct Link