
El-Sayed Ali El-Sherbiny
Department of Botany, Faculty of Science, Zagazig University, Zagazig, Egypt
Asian Journal of Biological Sciences
Year: 2011 | Volume: 4 | Issue: 1 | Page No.: 15-24







ABSTRACT
Streptomyces cyaneogriseus capable of utilizing chitosan as sole carbon source was isolated from soil samples. Chitosanase enzyme produced in the culture filtrate of Streptomyces cyaneogriseus was purified to homogeneity through ammonium sulfate precipitation (80%), ultrafiltration and Sephdex G-200 gel filtration. Molecular weight of the enzyme was 46 kDa. An optimum pH value was 5.0 and temperature was 50°C, respectively. The enzyme was stable (in absence of substrate) at temperature range from 40 to 60°C and at pH range from 3.0 to 8.0. The activity of chitosanase enzyme increased by the addition of 10 mM Mn2+ and Co2+. However, 10 mM Hg2+ and Cd2+ strongly inhibited the enzyme activity. Other metallic ions used (Na+, K+, Ca2+, Fe2+, Mg2+ and EDTA) have little effect on chitosanase activity. The enzyme also showed activity for hydrolysis of crystalline and colloidal chitosan but did not hydrolyze chitin, colloidal chitin, cellulose and carboxymethyl cellulose.
PDF Abstract XML References Citation
Received: January 20, 2010;
Accepted: May 06, 2010;
Published: August 18, 2010
How to cite this article
El-Sayed Ali El-Sherbiny, 2011. Purification and Characterization of Chitosanase Enzyme from Streptomyces cyaneogriseus. Asian Journal of Biological Sciences, 4: 15-24.
URL: https://scialert.net/abstract/?doi=ajbs.2011.15.24
URL: https://scialert.net/abstract/?doi=ajbs.2011.15.24
INTRODUCTION
Chitosan is one of the most abundant renewable polysaccharides prepared from chitin through chemical N-deacetylation. There is a growing interest in exploiting renewable resources for the production of biodegradable polymers (Hejazi and Amiji, 2003; Senel et al., 2000; Singla and Chawla, 2001; Senel and McClure, 2004). Chitosan, which is obtained by N-deacetylation of chitin (Roberts, 1992) has attracted some attention since chitin is the second most abundant polysaccharide. Chitosan oligosaccharides (pentamers, hexamers and glucosamine) are produced by partial hydrolysis of chitosan as reaction intermediates. There are two methods for hydrolysis of chitosan: chemical and enzymatic. Chemical hydrolysis is performed at high temperatures under highly acidic conditions and produces a large amount of glucosamine (chitosanmonomer), low yields of pentamers and hexamers are produced by this method due to difficulties in controlling the progress of the reaction. The advantages of using enzymatic hydrolysis for the production of chitosan oligosaccharides are that some chitosanases can catalyze the hydrolysis of chitosan under mild conditions. Chitosanolytic enzymes are gaining increasing importance as Low Molecular Weight Chitosans (LMWC) and chitooligomers, the products of chitosan hydrolysis, show various applications in various fields such as medicine, agriculture and biotechnology (Uchida et al., 1989; Suzuki et al., 1989; Suzuki, 1996; Somashekar and Joseph, 1996; Chatelet et al., 2001; Jeon et al., 2001; Zhang and Neau, 2002; Savard et al., 2002; Hejazi and Amiji, 2003; Mi et al., 2003).
Chitosanases (EC 3.2.1.132) are glycosyl hydrolases produced by many microorganisms, including fungi (Shimosaka et al., 1991; Zhang et al., 2000; Cheng and Li, 2000; Chen et al., 2005) actinomycetes (Price and Storck, 1975; Boucher et al., 1992; Sakai et al., 1991; Ngo et al., 2005), bacteria (Akiyama et al., 1999; Chiang et al., 2003; Omumasaba et al., 2000; Yoon et al., 2000; Choi et al., 2004; Chen et al., 2006; Yun et al., 2006) and plants (Yun et al., 2005; Somashekar and Joseph, 1996). Chitosanases are generally endo-splitting enzyme and could degrade chitosan into chitooligosaccharides and glucosamine as they catalyze the hydrolysis of the β-1,4-glycosidic linkage of chitosan (Fenton and Eveleigh, 1981).
In this study, Streptomyces cyaneogriseus was able to produce chitosanase enzyme and degrade chitosan into low molecular weight chitosan. This research was designed to study purification procedures and protocols for characterizing physicochemical properties of the enzyme.
MATERIALS AND METHODS
Materials: Chitosan and glucosamine were purchased from Sigma (St-Louis, MO, USA). Commercial chitosan was prepared from shrimp chitin in our laboratory as described by Hejazi and Amiji (2003). Colloidal chitosan was prepared by the method of Fen et al. (2006). Folin reagent was purchased from LOBA chemie (Mumbai, India). All other chemicals used were of analytical grade.
Strain and cultural condition: The strain used in this study was Streptomyces cyaneogriseus which was isolated and identified by the author El-Sherbiny (1998). The culture medium used through out this study was composed of: 0.5 g L-1 colloidal chitosan, 3.0 g L-1 KNO3, 1.0 g L-1 K2HPO4, 0.5 g L-1 NaCl, 0.5 g L-1 MgSO4 and 1000 mL tap water. The culture medium (pH 5.0) was inoculated with 1 mL spore suspension of S. cyaneogriseus at 30°C with agitation rate 150 rpm for 5 days. Then, the culture supernatant was obtained after the cells were sedimented by centrifugation at 5000 rpm for 15 min.
Chitosanase assay: Assay of chitosanase activity was carried out as described by Uchida and Ohtakara (1988) as well as Shimosaka et al. (1995) with some modification. The assay mixture containing 1 mL of crude enzyme filtrate and 1 mL of 1% colloidal chitosan in sodium acetate buffer (pH 5.0) was incubated at 37°C for 1 h. The concentration of reducing sugar was measured by the modified Schales’s method which was described by Imoto and Yagishita (1971) with d-glucosamine as a reference compound. One unit of chitosanase activity was defined as the amount of enzyme that produced 1 μmol of D-glucosamine equivalent in 1 min.
Determination of protein: The protein content of the crude enzyme preparation was estimated colorimetrically according to the method adapted by Lowry et al. (1951).
Purification of chitosanase: The cell-free supernatant was partially purified by fractional precipitation with Ammonium sulphate (80%). The precipitated protein was obtained by centrifugation for 15 min at 15000 rpm and tested for both chitosanase activity and protein content. The partially purified chitosanases precipitated at 80% ammonium sulphate saturation (dissolved in 0.1 M citrate phosphate buffer, pH 5.0) were dialyzed overnight against acetate phosphate buffer pH 5.0. The dialyzed chitosanase enzymes preparations were dissolved in a known volume (50 mL) of Citrate-Phosphate buffer at pH 5.0. These samples were loaded on the pharmacia column (60x2 cm) packed with Sephadex G200, which had been previously equilibrated with the same buffer (pH 5.0), used to dissolve the enzyme solution. The elute was obtained in 5 mL fractions. The protein content and chitosanase enzymes of each fraction were determined as described before. All purifications steps were carried out at 4.0°C.
Molecular weight of chitosanase: The molecular weight of chitosanase enzyme was measured by Sodium dodecyl sulfate-poly-acrylamide gel electrophoresis (SDS-PAGE) in10% gel was carried out by using the method of Laemmli (1970).
Characterization of purified enzyme: Effect of pH on enzyme activity and stability: The optimum pH of the chitosanolytic activity for the purified enzyme was assayed in citrate-phosphate buffer from 3.0 to 9.0 using colloidal chitosan as the substrate and incubated in 40°C for 60 min. For pH stability, the enzyme was treated at various pHs (without substrate) at 40°C for 90 min. The pH of enzyme solution was then adjusted to 5.0 and the remaining activities were measured.
Effect of temperature on enzyme activity and stability: The optimum temperature of the chitosanolytic activity of the purified enzyme was assayed at pH 5.0 from 40 to 80°C for 60 min. The enzyme was treated at different temperatures (40-80°C) for 90 min at pH 5.0 without the addition of substrate and was withdrawn at precise intervals to investigate thermal stability. The remaining activities were measured.
Effect of metal ions and some other reagents: The purified enzyme was incubated with 10 mM metal ions dissolved in the buffer and the activity was then measured at standard conditions.
RESULTS
Streptomyces cyaneogriseus was grown on culture medium containing colloidal chitosan as the sole carbon source and KNO3 as nitrogen source had produced chitosanase enzyme and secrete it in their culture fluid. The, partially purified chitosanase precipitated at 80% ammonium sulphate saturation (dissolved in 0.1 M citrate phosphate buffer, pH 5.0) was subjected to ultrafiltration. Partially purified chitosanase enzyme solution prepared by dialysis (ultrafiltration) was applied to gel filtration on Sephadex G200 column.
As shown in Fig. 1, gel filtration of partially purified chitosanase enzyme of Streptomyces cyaneogriseus yielded 60 fractions, the active 7 chitosanase fractions were collected and concentrated by dialysis for further investigation. The summary of purification steps were summarized in Table 1.
Molecular mass: The molecular mass of the chitosanase was 46 kDa, as estimated by SDS-PAGE Fig. 2.
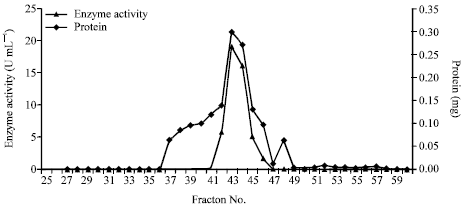 | |
| Fig. 1: | Gel filtration of chitosanase enzyme from S. cyaneogriseus on Sephadex G200 |
| Table 1: | Purification steps of chitosanase enzyme produced by Streptomyces cyaneogriseus |
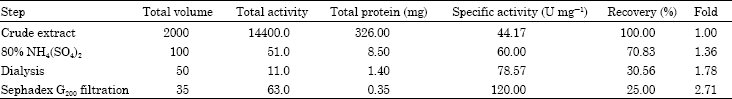 | |
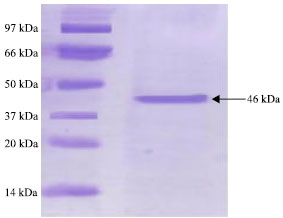 | |
| Fig. 2: | Analysis of the purified chitosanase enzyme from by SDS-Polyacrylamide Gel Electrophoresis (10%) |
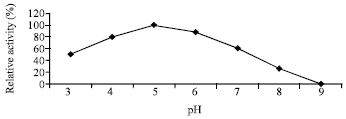 | |
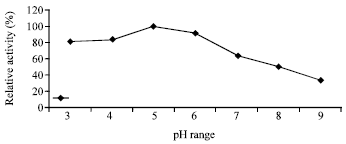
| Fig. 3: | Effect of different pH-values on the enzyme activity |
 | |
| Fig. 4: | Effect of pH-values on the stability of purified chitosanase |
Effect of pH on the activity and stability of enzyme: The purified chitosanase had an optimal pH of 5.0 for chitosan hydrolysis Fig. 3. When the enzyme was kept at 25°C in a Citrate-Phosphate buffer from pH 3 to 9 for 90 min, the enzyme was stable at pH 5-8 Fig. 4.
Effect of temperature on the activity and stability of enzyme: The purified chitosanase had an optimal temperature of 50°C for chitosan hydrolysis Fig. 5. When the enzyme was incubated without substrate at temperatures from 40 to 80°C for 30-90 min, the enzyme was stable at 40 and 50°C, gradually inactivated at 60 and 70°C and significantly lost activity above 70°C Fig. 6.
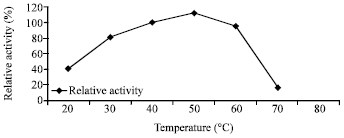 | |
| Fig. 5: | Effect of different temperatures on the enzyme activity |
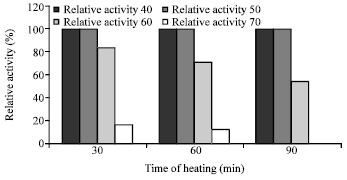 | |
| Fig. 6: | Effect of different temperatures on the enzyme stability |
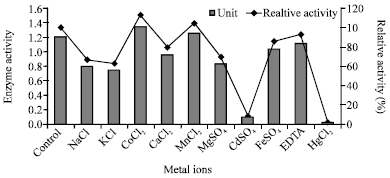 | |
| Fig. 7: | Effect of some metallic ions on the enzyme activity |
| Table 2: | Substrate specificity of S. cyaneogriseus chitosanase |
 | |
Substrate specifity: The activities of the purified chitosanase up on chitosan, colloidal chitosan, chitin, colloidal chitin, cellulose and CMC are presented in Table 2.
Effect of metal ions: As shown in Fig. 7, the chitosanase was significantly inhibited by 10 mM of Hg2+and Cd2+ but not significantly affected by other metal ions. Co2+ and Mn2+ had a stimulatory effect on chitosanase enzyme produced by S. griseous. Other metallic ions had slight remarkable effect on chitosanase activity.
DISCUSSION
This study described the purification and characterization of chitosanase from Streptomyces cyaneogriseus. The purification of the crude chitosanase produced in culture filtrates of Streptomyces cyaneogriseus were carried out by fractional precipitation with 80% saturation ammonium sulfate showing 1.36 fold and specific activity of chitosanase (60 U mg-1 protein). The protein fractions recovered from ammonium sulphate precipitation were dialysis against citrate-phosphate buffer pH (5.0). The dialysis technique led to an increase in the specific activity of chitosanase enzyme up to 78.57 U mg-1 protein with purification folds 1.78 (fold). The next step on purification of this dialyzed enzyme preparation was gel filtration on a column of sephadex G200 elution with citrate-phosphate buffer at pH 5.0. Active fractions of the sharp peaks of fractional purification curve were collected and concentrated by dialysis against the same buffer for having a concentrated preparation of the purified- chitosanase. This led to the recovery of 25% of chitosanase activity. The molecular mass of the purified enzyme was 46 kDa as estimated by SDS-PAGE. This suggested that the purified enzyme was a single protein. Most microbial chitosanases had molecular masses in the range of 10-50 kDa (Fenton and Eveleigh, 1981; Somashekar and Joseph, 1996; Cheng and Li, 2000; Zhang et al., 2001; Tanabe et al., 2003; Kim et al., 2004; Chen et al., 2005). The molecular mass of this enzyme is in the range of reported values. The enzyme had its maximum activity at pH 5.0 when assayed with colloidal chitosan as substrate for enzyme reaction. These results were comparable to those obtained for other microbial chitosanase which showed optimum pH ranges between 5 to 5.5 (Somashekar and Joseph, 1996). The activity of enzyme increase with increasing pH then followed by slightly decrease in activity after optimum pH (5.0) but there is no activity detected at pH 9.0. Chitosanase enzyme was stable in the pH range of 3 to 7 and lost about 50 and 66.7% at pH 8.0 and 9.0. When enzyme was assayed at different temperatures, the optimum activity was achieved at 50°C. The enzyme was stable at temperature range from 40 to 60°C. This is in agree with (Somashekar and Joseph, 1996; Chiang et al., 2003; Kim et al., 2004; Chen et al., 2005; Liu and Xia, 2006; Su et al., 2006; Yun et al., 2006). The chitosanase of various species were found to be stable up to 50 C (Somashekar and Joseph, 1996). The activity of S. cyaneogriseus chitosanase lost its activity above 70°C. In this study we found that the optimum concentration of the substrate required for maximum enzyme activity was 5.0 mg L-1. This might be due to saturation with higher concentration of chitosan or to product inhibition (Abdel-Aziz, 1994). Other results reported for other microbial chitosanases are from 0.5 to 1.0 mg (Abdel-Aziz, 1999; Cheng and Li, 2000; Aktuganov et al., 2003; Chen et al., 2005).
The substrate specificity of the enzyme was studied by monitoring the hydrolysis of various substrates. Most microbial chitosanases were specific for chitosan degradation (Fenton and Eveleigh, 1981; Yoshihara et al., 1992; Price and Storck, 1975; Tominaga and Tsujisaka, 1975). However, some of these enzymes also showed activity for hydrolysis of carboxymethyl cellulose (Hedges and Wolfe, 1974) or colloidal chitin (Yamasaki et al., 1993). The enzyme from S. cyaneogriseus hydrolyzed chitosan and colloidal chitosan, but did not hydrolyze chitin, colloidal chitin, cellulose and carboxymethyl cellulose at all. The effect of deacetylation of chitosan on the activity of chitosanases has been reported for the enzymes from P. islandicum (Fenton and Eveleigh, 1981), Bacillus circulans MH-K1 (Yabuki et al., 1988) and Bacillus sp. 7I-7S (Seino et al., 1991). Chitosanases show differing specificities for substrates. Schindler et al. (1977) have hypothesized that evolutionary changes in substrate structure may have influenced the development of the active site of lysozyme so that it could function most efficiently with the particular natural substrate encountered by each species. S. cyaneogriseus chitosanse was activated by Mn+2 and Co+2.these results are similar to those obtained by Liu and Xia (2006). Also, Chitosanase of Rhodotorula gracilis and A. fumigatus KH 94 was enhanced by Mn2+ and greatly inhibited by Hg2+ and Cu2+ (Somashekar and Joseph, 1992; Kim et al., 1998). Hg2+ and Cd2+ were significantly or completely inhibited the enzyme thus, cysteine is essential for the active site of the enzyme (Somashekar and Joseph, 1996). Cheng and Li (2000) reported that the purified chitosanase of A. sp Y2K greatly enhanced by Mn2+, Cu2+ and Cu+ ions at low concentration and strongly inhibited by Hg2+ and Cd2+ ions. The chitosanase activity of Bacillus sp. MET 1299 was increased by addition of 1 mM Mn2+, Ca2+, Mg2+ and Ba2+. However, Hg2+ and Fe2+ ions in the assay solution strongly decreased the enzyme activity (Kim et al., 2004).
Finally, the purified chitosanase hydrolyzed colloidal chitosan and crystalline chitosan into a biofunctional oligomer at a high-level. Therefore, this enzyme is a good candidate for biotechnological application to produce chitooligosaccharides, which may be used as antifungal, antibacterial and antitumer activities. However, further studies are required for other practical application of this enzyme.
REFERENCES
- Abdel-Aziz, S.M., 1999. Production and some properties of two chitosanases from Bacillus alvei. J. Basic Microbiol., 39: 79-87.
CrossRefDirect Link - Akiyama, K., T. Fujita, K. Kuroshima, T. Sakane, T. Yokota and R. Takata, 1999. Purification and gene cloning of a chitosanase from Bacillus ehimensis EAG1. J. Biosci. Bioeng., 87: 383-385.
PubMed - Aktuganov, G.E., A.V. Shirokov and A.I. Melent`ev, 2003. Isolation and characterization of chitosanase from the strain Bacillus sp. 739. Applied Biochem. Microbiol., 39: 469-474.
CrossRef - Boucher, I., A. Duppy, P. Vidal, W.A. Neugebauer and R. Brzezinski, 1992. Purification and characterization of a chitosanase from Streptomyces N174. Applied Microbiol. Biotechnol., 38: 188-193.
CrossRef - Chatelet, C., O. Damour and A. Domard, 2001. Influence of the degree of acetylation on some biological properties of chitosan films. Biomaterials, 22: 261-268.
CrossRef - Chen, Xe, X. Wenshui and Y. Xiaobin, 2005. Purification and characterization of two types of chitosanase from Aspergillus sp. CJ22-326. Food Res. Int., 38: 315-322.
Direct Link - Chen, Y-L., C. Su and B.H. Chiang, 2006. Optimization of reversed micellar extraction of chitosanases produced by Bacillus cereus. Process Biochem., 41: 752-758.
CrossRef - Cheng, C.Y. and Y.K. Li, 2000. An Aspergillus chitosanase with potential for large-scale preparation of chitosan oligosaccharides. Biotechnol. Applied Biochem., 32: 197-203.
Direct Link - Chiang, C.L., C.T. Chang and H.Y. Sung, 2003. Purification and properties of chitosanase from a mutant of Bacillus subtilis Enzyme Microbiol. Technol., 32: 260-267.
CrossRef - Choi, Y.J., E.J. Kim, Z. Piao, Y.C. Yun and Y.C. Shin, 2004. Purification and characterization of chitosanase from Bacillus sp. strain KCTC 0377BP and its application for the production of chitosan oligosaccharides. Applied Environ. Microbiol., 70: 4522-4531.
CrossRefDirect Link - Fen, L.L., R.M. Illias, K. Kamaruddin, M.Y. Maskat and O. Hassan, 2006. Development of rapid screening method for low-yielding chitosanase activity using Remazol Brilliant Blue-chitosan as substrate. Enzyme Microbiol. Technol., 38: 215-219.
CrossRef - Fenton, D.M. and D.E. Eveleigh, 1981. Purification and mode of action of a chitosanase from Penicillium islandicum. J. Gen. Microbiol., 126: 151-165.
CrossRefDirect Link - Hedges, A. and R.S. Wolfe, 1974. Extracellular enzyme from Myxobacter AL-1 that exhibits both β-1,4-glucanase and chitosanase activities. J. Bacteriol., 120: 844-853.
Direct Link - Hejazi, R. and M. Amiji, 2003. Chitosan-based gastrointestinal delivery systems. J. Control Release, 89: 151-165.
CrossRefPubMedDirect Link - Imoto, T. and K. Yagishita, 1971. A simple activity measurement of lysozyme. Agric. Biol. Chem., 35: 1154-1156.
Direct Link - Jeon, Y.J., P.J. Park and S.K. Kim, 2001. Antimicrobial effect of chitooligosaccharides produced by bioreactor. Carbohydr. Polym., 44: 71-76.
CrossRef - Kim, P.I., T.H. Kang, K.J. Chung, I.S. Kim and K.C. Chung, 2004. Purification of a constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and expression of the gene. FEMS Microbiol. Lett., 240: 31-39.
PubMed - Kim, S.Y., D.H. Shon and K.H. Lee, 1998. Purification and characteristics of two types of chitosanase from Aspergillus fumigatus KH-94. J. Microbiol. Biotechnol., 8: 568-574.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Liu, J. and W. Xia, 2006. Purification and characterization of a bifunctional enzyme with chitosanase and cellulase activity from commercial cellulase. Biochem. Eng. J., 30: 82-87.
CrossRef - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mi, F.L., Y.B. Wu, S.S. Shyn, A.C. Chao, J.Y. Lai and C.C. Su, 2003. A symmetric chitosan membranes prepared by dry/wet phase separation: A new type of wound dressing for controlled antibacterial release. J. Membr. Sci., 212: 237-254.
CrossRef - Ngo, K.X., H. Umakoshi, T. Shimanouchi, H.S. Jung, S. Morita and R. Kaboi, 2005. Heat-enhanced production of chitosanase from Streptomyces griseus in the presence of liposome. J. Biosci. Bioeng., 100: 495-501.
PubMed - Omumasaba, C.A., N. Yoshida, Y. Sekiguchi, K. Kariya and K. Ogawa, 2000. Purification and some properties of a noval chitosanase from Bacillus subtillus KH1. J. Gen. Applied Microbiol., 46: 19-27.
Direct Link - Price, J.S. and R. Storck, 1975. Production, purification and characterization of extra-cellular chitosanase from Streptomyces. J. Bacteriol., 124: 1574-1585.
PubMedDirect Link - Sakai, K., R. Katsumi, A. Isobe and F. Nanjo, 1991. Purification and hydrolytic action of a chitosanase from Norcardia orientalis. Biochim. Biophys. Acta, 1079: 65-72.
PubMed - Savard, T., C. Beaulieu, I. Boucher and C.P. Champagne, 2002. Antimicrobial action of hydrolyzed chitosan against spoilage yeasts and lactic acid bacteria of fermented vegetables. J. Food Prot., 65: 828-833.
PubMed - Schindler, M., D. Mirelman and S. Sharon, 1977. Substrate induced evolution of lysozymes. Biochim. Biophys. Acta (BBA)-Enzymol., 482: 386-392.
CrossRef - Senel, S. and J.S. McClure, 2004. Potential application of chitosan in veterinary medicine. Adv. Drug Deliv. Rev., 56: 1467-1480.
CrossRef - Senel, S., M.J. Kremer, S. Kas, P.W. Wertz, A.A. Hincal and C.A. Squier, 2000. Enhancing effect of chitosan on peptide drug delivery across buccal mucosa. Biomaterials, 21: 2067-2071.
PubMed - Shimosaka, M., M. Nogawa, X.Y. Wang, M. Kumehara and M. Okazaki, 1995. Production of two chitosanases from a chitosan-assimilating bacterium, Acinetobacter sp. Strain CHB101. Applied Environ. Microbiol., 61: 438-442.
Direct Link - Shimosaka, M., M. Nogawa, Y. Ohno and M. Okazaki, 1991. Chitosanase from the plant pathogenic fungus, Fusarium solani f. sp. phaseoli: Purification and some properties. Biosci. Biotechnol. Biochem., 57: 231-235.
Direct Link - Singla, A.K. and M. Chawla, 2001. Chitosan: Some pharmaceutical and biological aspects-an update. J. Pharm. Pharmacol., 53: 1047-1067.
CrossRefPubMedDirect Link - Somashekar, D. and R. Joseph, 1992. Partial purification and properties of a novel chitosanase secreted by Rhodotorula gracilis. Lett. Applied Microbiol., 14: 1-4.
CrossRefDirect Link - Somashekar, D. and R. Joseph, 1996. Chitosanases-properties and applications. Bioresour. Technol., 55: 35-45.
Direct Link - Su, C., D. Wang, L. Yao and Z. Yu, 2006. Purification, characterization and gene cloning of chitosanase from Bacillus species strain S65. J. Agric. Food Chem., 54: 4208-4214.
CrossRefDirect Link - Tominaga, Y. and Y. Tsujisaka, 1975. Purification and some enzymatic properties of the chitosanase from Bacillus R-4 that lyses Rhizopus cell walls. Biochem. Biophys. Acta, 410: 145-155.
PubMedDirect Link - Uchida, Y. and A. Ohtakara, 1988. Chitosanase from Bacillus species. Methods Enzymol., 161: 501-505.
CrossRef - Yabuki, M., A. Uchiyama, K. Suzuki, A. Ando and T. Fujii, 1988. Purification and properties of chitosanase from Bacillus circulans MH-K1. J. Gen. Applied Microbiol., 34: 255-270.
Direct Link - Yamasaki, Y., I. Hayashi, Y. Ohda, T. Nakagawa, M. Kawamukai and H. Matsuda, 1993. Purification and mode of action of chitosanolytic enzyme from Enterobacter sp. G-1. Biosci. Biotechnol. Biochem., 57: 444-449.
Direct Link - Yoon, H.G., H.Y. Kim, Y.H. Lim, H.K. Kim, D.H. Shin, B.S. Hong and H.Y. Cho, 2000. Identification of essential amino acid residues for catalytic activity and thermostability of novel chitosanase by site-directed mutagenesis. Applied Microbiol. Biotechnol., 56: 173-180.
PubMed - Yoshihara, K., J. Hosokawa, T. Kubo, M. Nishiyama and Y. Koba, 1992. Purification and properties of a chitosanase from Pseudomonas sp. H-14. Biosci. Biotech. Biochem., 56: 972-973.
Direct Link - Yun, C.S., D. Amakata, Y. Matsuo, H. Matsuda and M. Kawamukai, 2005. New chitosan-degrading strains that produce chitosanase similar to ChoA of Mitsuaria chitosanitabdia. Applied Environ. Microbiol., 71: 5138-5144.
Direct Link - Yun, C., H. Matsuda and M. Kawamukai, 2006. Directed evolution to enhance secretion efficiency and thermostability of chitosanase from Mitsuaria chitosanitabida 3001. Biosci. Biotechnol. Biochem., 70: 559-563.
PubMed - Zhang, H. and S.H. Neau, 2002. In vitro degradation of chitosan by bacterial enzymes from rat cecal and colonic contents. Biomaterials, 23: 2761-2766.
CrossRef