
Y. Smith
Department of Zoology and Entomology,
H.O. Waal
Department of Animal, Wildlife and Grassland Sciences,
University of the Free State, P.O. Box 339, Bloemfontein 9300, South Africa
O. Kok
Department of Zoology and Entomology,
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 11 | Page No.: 2149-2152







ABSTRACT
Feed intake and digestibility trials were conducted on captive African lions (Panthera leo). An adult female and sub-adult male were used in a zoo environment and a sub-adult male and large cub female under reserve conditions. The lions were fed large chunks of fresh animal carcasses under controlled conditions. Afterwards, refusals and feces were collected over several days, analysed and weighed. Apparent digestibilities were fairly high ranging from 77.4 to 94.3% in the zoo and from 83.8 to 95.6% at the nature reserve. Fecal excretion was highly variable and irregular, the lions excreting up to four times following food ingestion and in some cases only twice. Possible reasons for this are discussed. The consistency, moisture content and colour of the scat also varied between the two trial sites. Plant material was regularly ingested in both environments, apparently to assist in purging the digestive tract. Being obligatory carnivores, the gastrointestinal tract of four African lions of different ages and sexes was found to be relatively simple, short and uncomplicated.
PDF Abstract XML References Citation
How to cite this article
Y. Smith, H.O. Waal and O. Kok, 2006. Aspects of Carcass Digestibility by African Lions (Panthera leo Linnaeus, 1758) under Captive Conditions. Pakistan Journal of Biological Sciences, 9: 2149-2152.
DOI: 10.3923/pjbs.2006.2149.2152
URL: https://scialert.net/abstract/?doi=pjbs.2006.2149.2152
DOI: 10.3923/pjbs.2006.2149.2152
URL: https://scialert.net/abstract/?doi=pjbs.2006.2149.2152
INTRODUCTION
A large volume of studies have been conducted on various aspects of the African lion (Panthera leo Linnaeus, 1758), with the primary emphasis on behaviour and social organisation (Guggisberg, 1961; Bertram, 1975; Packer et al., 2001; Eloff, 2002) and prey selection and hunting ecology (Smuts, 1979; Eloff, 1984; Stander, 1992; Packer and Pusey, 1997; Viljoen, 1997). There is, however, a paucity of quantitative information on the nutritional requirements and digestive capacity of the species. Furthermore, no quantitative data have been published on the excretion rate of these animals. Here we report on the apparent digestibility and rate of fecal excretion of captive African lions under different holding conditions. The gross dimensions of the gastrointestinal tract were also investigated.
MATERIALS AND METHODS
Digestibility trials were performed on four captive African lions at the Bloemfontein Zoological Gardens (29°06.809'S, 26°12.387'E) and Wag-‘n-Bietjie Private Nature Reserve (28°56.007'S, 26°17.421'E), approximately 40 km north of the city in the Brandfort district, central Free State, during winter 2001 and 2002, respectively. Experimental animals included a sub-adult male (27 months) and an adult female (four years) held in enclosures with separate night chambers and a spacious open air leisure yard at the zoo and a sub-adult male (27 months) and a large cub female (15 months) held in an enclosure of 1.1 ha at the nature reserve.
At the outset of the study, fresh lion scats were collected for a period of one month to determine the consistency, colour, moisture content, mass and frequency of fecal excretion following the ingestion of 30 kg carcass component per week, thereby ensuring the efficient identification and collection of fresh scat during subsequent digestibility trials. The latter were performed with large, unprocessed fore- and hindlimbs of donkeys (Equus asinus Linnaeus, 1758), kudu (Tragelaphus strepciseros Pallas, 1766) and red hartebeest (Alcelaphus buselaphus Pallas, 1766) or halved cattle foetuses (Bos taurus Linnaeus, 1758). A whole, fresh donkey skin was used in one trial. In all cases the mirror image, consisting of opposite limb sections or foetuses cut lengthwise along the spine, were sectioned into muscle/fat/tissue, bone and skin components and immediately weighed, carefully covered in plastic cling wrap, sealed in plastic bags and frozen at -10°C for later analyses.
Prior to feeding (for comparative purposes always at 15:00), the holding facilities at the zoo and nature reserve were cleared of all scat. Based on the results attained regarding rates of fecal excretion, the enclosures were revisited between 06:00 and 07:00 the next morning and all refusals and fresh scat collected, sealed in airtight plastic bags (Ziploc®) and frozen at -10°C. Scat, when present, was collected again at 14:00 and between 16:00 and 19:00 and on the third day, at 07:00 and 14:00.
Laboratory analyses involved the thawing of the scat and carcass components while still sealed. For preparation for drying in a force drought oven, all muscle/tissue/fat and skin sections were cut into small strips and suspended with wire hooks from shelves in the oven. Bones were placed upright on specially designed racks to capture dripping oil. The drying process was started by setting the oven at 80°C for at least 48 h after which the temperature was increased to 100°C for a further two to three days, depending on the amount of meat and bone to be dried. After drying, the samples were cooled in a desiccator and weighed again. The apparent digestibilities of the diets were calculated on an as fed (fresh) and dry matter basis as follows:
where Food ingested = Food given-Food refused |
Four lion specimens, an adult (six years) and sub-adult female (3.5 years), a sub-adult male (three years) and a stillborn male, were obtained for examination of the gastrointestinal tract. The abdomen and chest cavity of the carcass was opened and the gastrointestinal tract removed from behind the tongue through to the anus. The mesentery with the mesenteric arteries, veins, lymphatic vessels, nerves and fat were removed and the tract measured. Excepting for the first specimen where the outer curvature of the stomach was erroneously measured, the inner curvature of the stomach was taken for the remaining dissections.
RESULTS
Fecal excretion: A total of 16 feedings were observed at the zoo during the period of one month of which six from the sub-adult male are presented in Fig. 1. The rate of fecal excretion under nature reserve conditions were taken from the digestibility trials in both animals and are shown in Fig. 2. The frequency and mass of the defecation was highly irregular, the number varying from two to four with assorted masses.
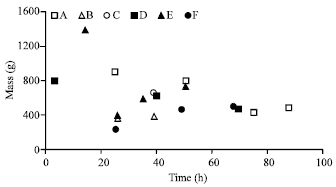 | |
| Fig. 1: | Time interval and mass (g) of feces excreted by the sub-adult male African lion subsequent to the feeding of different carcass components under zoo conditions. Open squares, triangles and circles represent the whole forelimb, dissected forelimb and hindlimb of a red hartebeest, respectively, while solid squares, triangles and circles represent the forelimb of a kudu and fore- and hindlimb of a donkey, respectively |
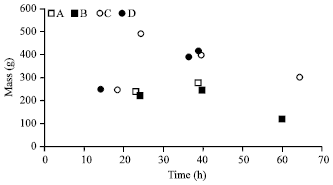 | |
| Fig. 2: | Time interval and mass (g) of feces excreted by African lions subsequent to the feeding of halved Bos taurus carcasses under nature reserve conditions. Squares represent trials with the large cub female and circles, the sub-adult male |
Further, a marked difference occurred between the masses of the scat of the two sub-adult males with the highest recorded in the zoo at 1500 g and in the reserve, 500 g. Both were 27 months at the time of the trials which were both performed during winter. The lions in the reserve were fed foetal carcasses and the number and mass of excretions were generally lower (Fig. 2). In the zoo, the sub-adult male was fed a whole and dissected forelimb of the same red hartebeest carcass (Fig. 1) and in the case of the latter component, the mass and number of the scats was comparable to the results obtained in the nature reserve. Time intervals for defecation also fluctuated, from three to 29 h between scats.
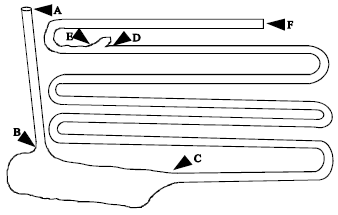 | |
| Fig. 3: | The gastrointestinal tract of an adult African lion with the loops of the intestine unfolded and laid out in one plane. The mesentery (with the mesenteric arteries, veins, lymphatic vessels, nerves and fat) has been removed. A-B, oesophagus; B-C, stomach; C-D, small intestine (duodenum, jejunum, ileum); D-F, large intestine (caecum, colon, rectum); D-E, caecum |
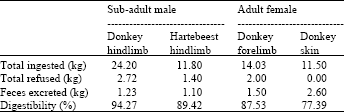
| Table 1: | Apparent digestibility of different carcass components by African lions under zoo conditions |
 | |
| Table 2: | Apparent digestibility of halved Bos taurus foetuses by African lions under nature reserve conditions |
 | |
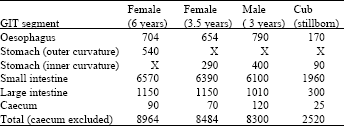
| Table 3: | Dimensions (mm) of the different sections of the gastrointestinal tract (GIT) of four African lions |
 | |
The colour ranged from black to dark or light brown at the first excretion through to light grey or white at the final excretion. Consistencies were also erratic from watery diarrhoea to firm or granular. In general, the first scat was moist and dark, drying and lightening to the final scat which was generally grey and dry. In the nature reserve, the scat was found to have an overall drier consistency than that collected under zoo conditions.
Digestibility trials: The same number of trials was performed at each site, namely two per animal. The details and results of the trials are presented in Table 1 and 2 for the zoo and nature reserve, respectively. The digestibility of the fed carcass components is fairly high, above 80%, with an equally high result attained in the trial using only a fresh skin. The digestibilities for the foetal carcasses are similar to that of adult animal carcasses although the total mass excreted is lower in the former. Refusals were generally low or absent and crushed bone and bleached hair voided in the feces indicate utilization of the nutritional value of these elements.
Gastrointestinal tract: Figure 3 shows a typical gastrointestinal tract of an African lion. The tract was found to be relatively simple, short and uncomplicated. The gross dimensions of the gastrointestinal tracts of the four lions examined are presented in Table 3. The different segment lengths in the adults are similar and roughly three times the length of that measured in the stillborn cub.
DISCUSSION
Nutritional aspects are an important facet in the survival of both individual lions and whole prides when taking into account the effects of migration patterns, climate change, droughts, ecosystem degradation and other environmental pressures on both the African lion and its prey species. Although the African lion has few natural enemies, environmental pressures are high and individuals must be exceptionally fit to survive. Their place at the top of the ecosystem places them in the position that their survival is dependent on entire ecosystem function.
Eloff (1980) reports that in the Kalahari 50% of cubs die prior to reaching the age of two years. In these extreme environments, lion densities are low. The Kgalagadi Transfrontier Park supports one lion/207 ha (www.Scienceinafrica.com), while Etosha National Park has one lion/13500-5400 ha (H.H. Berry, Personal communication). Kruger National Park, roughly half the size of Etosha, can maintain one lion/950 ha (www.Sanparks.com). This demonstrates that prey abundance regulates lion numbers. It is, however, the quality of the prey animal that affords the predator adequate nutrition. This is especially true in the case of lionesses which are suckling cubs. Under zoo conditions where feeding is strictly controlled and environmental factors regulated, lionesses still lose condition while rearing cubs (Personal observation). Further to this, under natural conditions, a hierarchy exists at feeding sites with the males feeding first, primarily from the hindquarters, followed by the females and cubs (Packer et al., 2001).
No comparative data exists on the frequency of excretion in lions. In studies that have included elements found in scat, such as with Eloff (2002) and Müller-Graf et al. (1999), scat were collected on an opportunistic basis. Our results indicate that when a carcass is soft (foetal or dissected), absorption is increased and the mass of fecal matter is decreased. It is probable that the rate of digestion and passage through the tract is influenced by the consistency of the meat and digestion is, therefore, more complete.
The difference in moisture levels of the scat between the two sub-adult males in the different conditions may be due to the fact that although both studies were conducted in the winter season, the lions in the reserve were fully exposed to environmental conditions. These included sub-zero temperatures at night and dawn and heavy frost. The lions in the zoo, however, always had access to the sleeping chambers equipped with infrared light heaters. Further, it was noted that the lions in the nature reserve drank less water than the lions in the zoo although this was not quantitatively measured.
Ingestion of plant material, with grass found in the scat, occurred at both sites. In the case of the zoo, the grass was always Kikuyu (Pennisetum clandestimum) from the ground cover in the leisure yard. In the nature reserve, plant material in the scat varied as several types of grasses grew in the enclosure. The frequency of plant material in the scat increased as hair and bone were voided and the scat became drier. It is possible that the plant material could be used for a bottle brush action, cleaning the rugae which increase in intensity as the stomach shrinks. It must also pass through the stomach quickly since in the case of Kikuyu grass, it was not found to be discoloured.
In the reserve enclosures, feces were consistently voided where the animals were positioned. This indicates that in the case of the research subjects, defecation occurred not at a specific site, but rather according to the movement of the animal involved. This concurs with published data since lions are not known to form latrines (Skinner and Smithers, 1990). Further research is required into nutrient extraction and water consumption and turnover to determine whether lions exposed to natural conditions have more stringent nutrient requirements than those kept under zoo conditions.
The length of the stillborn cub’s gastrointestinal tract, being roughly a third of that of the adults, indicates that although a total body increase of some 200% is required to adulthood, the gastrointestinal tract needs only to triple its initial length. The protein required for the development of an adult male lion indicate the efficacy of the gastrointestinal tract in the digestion and absorption of food and nutrients. The caecum is positioned in the reverse order when compared to that of the human gastrointestinal tract. This no doubt has evolved due to the consumption of large pieces of bone and even porcupine quills (Smuts, 1979). The position of the caecum prevents these elements from becoming lodged.
ACKNOWLEDGMENTS
We thank Sarel van der Merwe and George and Madeleen Croucamp for permission to conduct the study at Bloemfontein Zoological Gardens and Wag-‘n-Bietjie Private Nature Reserve. The Windhoek Observer is thanked for generous financial contributions.