
Saba Haider
Department of Botany and Jinnah University for Women 5-C, Nazimabad 74600, Karachi,
Salma Kanwal
Department of Chemistry, Jinnah University for Women 5-C, Nazimabad 74600, Karachi,
Fahim Uddin
University of Karachi, Pakistan
Rafia Azmat
Department of Chemistry, Jinnah University for Women 5-C, Nazimabad 74600, Karachi,
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 11 | Page No.: 2062-2068







ABSTRACT
Lead concentration is increasing rapidly in the environment due to increased use of its sources by human society. Photosynthesis has been found to be one of the most sensitive plant physiological process and the effect of metal is multi facial. This investigation discussed the toxicity of Pb on chlorophyll content and carbohydrate of Phaseolus mungo and Lens culinaris. Growth and pigments contents of both species were highly decreased and inhibited at 250 ppm of Pb concentration. The absorption spectrum of pigments were obtained by UV/Visible double beam Schimadzo Spectrophotometer. The observed spectral change due to enhancement of Pb dose, led to significant reduction in chlorophyll biosynthesis with values of 1.004 to 0.023 for Phaseolus mungo and 1.415 to 0.426 for Lens culinaris. This may be related with the accumulation of Pb in root and shoot. The absorption spectrum shows the change in structure of chlorophyll, which indicated that absorption of Pb as compare to essential mineral ion was higher. Pb replaces the magnesium and imparts the color due to which dark green leaves of both species were observed and leaves of plant remain green after experience of death. It may be attributed with less concentration of Mg in chlorophyll. Observation revealed that stomata density in the epidermis was noticeably reduced in presence of Pb due to which quantum efficiency for released of oxygen decreased.
PDF Abstract XML References Citation
How to cite this article
Saba Haider, Salma Kanwal, Fahim Uddin and Rafia Azmat, 2006. Phytotoxicity of Pb: II. Changes in Chlorophyll Absorption Spectrum due to Toxic Metal Pb Stress on Phaseolus mungo and Lens culinaris. Pakistan Journal of Biological Sciences, 9: 2062-2068.
DOI: 10.3923/pjbs.2006.2062.2068
URL: https://scialert.net/abstract/?doi=pjbs.2006.2062.2068
DOI: 10.3923/pjbs.2006.2062.2068
URL: https://scialert.net/abstract/?doi=pjbs.2006.2062.2068
INTRODUCTION
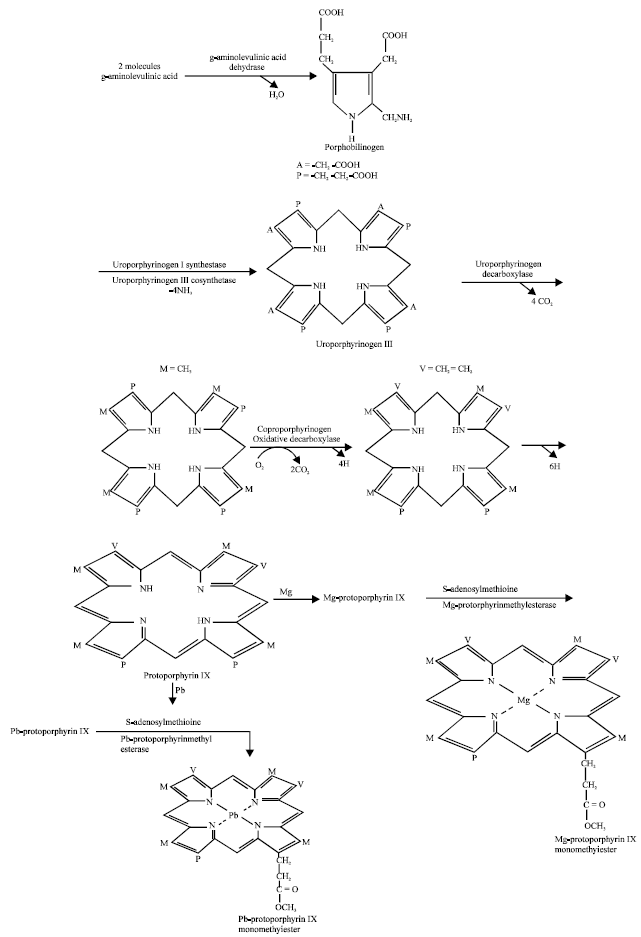
As many of the Pb pollutant are indispensable for modern human life, soil contamination with Pb is not likely to reduce in near future. It is easily taken up by plants from soil and accumulated in different organs (Baryla et al., 2001; Ewais, 1997). Photosynthetic pigments are essential for plant development. The studies on the effect of heavy metals on the photosynthesis apparatus (Csatorday et al., 1984) have raised the question about possible mechanism of their toxicity towards its integrity and functional aspects (Baker, 1981) whereas leaf chlorophyll content (Doring et al., 1969) provides value able information about physiological status of plant. Reflectance measurement makes its possible to quickly and nondestructively asses, in situ, the chlorophyll content in leaves. The process of photosynthesis is adversely affected by toxicity. Plant exposed to Pb show a decline in photosynthesis rate which results from distorted chloroplast ultra structure, restrained synthesis of chlorophyll, plastoquinone and carotenoids, obstructed electron transport, inhibited activities of Calvin cycle enzymes, as well as deficiency of CO2 as a result of stomata closure. Pb inhibits chlorophyll synthesis by impairing uptake of essential elements of chlorophyll such as Mg and Fe. The mechanism proposed for this inhibition is the replacement of magnesium in the chlorophyll molecule. Consequently cells accumulate protoporphyrin and synthesis of chlorophyll is blocked. In this regard the reduction of chlorophyll content is a common symptom of heavy metals toxicity. This may be attributed to inhibition of reduction steps in the biosynthesis pathways of the pigments (Hartmut et al., 1990). It damages the photosynthetic apparatus due to its affinity for protein N-and S- ligands. The chlorophyll content over a wide range of its changes can be assessed during leaf ontogeny using reflectance near 700 nm (Gitelson et al., 2002, 2003). Reflectance spectra of leaves and fruits undergo remarkable changes under deficiency of mineral nutrition, pollution, different stress conditions, during adaptation to variable solar irradiation and in the course of senescence (Merzlyak et al., 2003; Solvchenko et al., 2001). Reduced accumulation of cholorophyll in Cd, Cu, Mn and Ni treated plant have been reported by Siedlecka et al. (2001). The substitution of heavy metal in place of Mg as a central ion in the prophyrine ring of chlorophyll or bacterochlorophyll in vivo, for algae, lichen, water plants and in vitro for isolated pigments were reported by Ewais (1997). Leaf chlorosis was attributed to a marked decrease in the chloroplast density caused by a reduction in number of chloroplast per cell and change in cell size suggesting that Cd interfered with chloroplast (Azmat et al., 2005a) replication and cell division.
Phaseolus mungo and Lens culinaris are widely used by people of south Asia and as well as in Pakistan and these pulses are imported in Pakistan from other countries. Keeping this view, this study has been planned to investigate the pigments and starch contents of both species under Pb stress. Toxic metal is widespread potent pollutant of environment which is present both in air and water of irrigation. Chlorophyll and carbohydrate will be determined by usual method and absorption spectrum will be observed by UV/Visible double beam Schimadzo Spectrophotometer. The results will be discussed in relation with photosynthesis and growth regarding with carbohydrate concentration.
MATERIALS AND METHODS
The experiments were performed in growth chamber in department of Botany in Jinnah University for Women in July 2005. Phaseolus mungo and Lens culinaris were selected for cultivation under PbCl2 treatment in pots of Hoagland solution.
For chlorophyll and other pigments, an extract of plants in 80% acetone were prepared. Spectral data for both spices were collected at close-ranges of UV/Visible Schimadzo spectrophotometer. The system detects research spectral data in the range of 200 to 800 nm. (Bohlar and Elisabeth, 1987).
Carbohydrates were analyzed in protein free filtrate extract of plants in water using anthrone reagent and complex absorbance was observed by spectrophotometer at 650 nm (Tandon, 1993).
After passing 2 days, the following agronomic parameters were determined a) Plant height b) Root Length c) Shoot fresh mass and shoot dry mass d) Leaf area e) Percentage of germination f) determination of pigments concentration (chlorophyll a and b and total chlorophyll) by spectrophotometer. Pigments in the leaves were extracted with 80% acetone and data were used to calculate the concentration of pigments.
RESULTS AND DISCUSSION
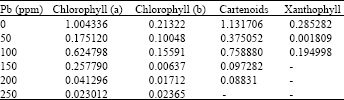
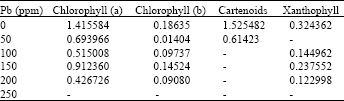
Results showed that the Pb affects the morphology of leaves (Fig. 1 and 2). Up to 50 ppm of Pb concentration, leaves showed normal growth and color while on high dose of Pb, leaves size were shortened with dark color. This indicated that process of photosynthesis was adversely affected by Pb toxicity. Plant exposed to Pb ions shows a decline in ultra structure, restrained synthesis of chlorophyll (Azmat et al., 2005a-c) plastoquinone and carotenoids (Truscott, 1990), obstructed electron transport as well as efficiency of CO2 as a result of reduced stomata density (Tripathy et al., 1983). Phaseolus mungo and Lens culinaris when grown in aquatic medium (Pb based Haogland solution) showed distinct changes in wavelength of chlorophyll a and b. Table 1 and 2 shows the change in maximum optical density with the change in concentration of Pb, which can be attributed with accumulation of Pb in leaves due to which decreased in concentration of chlorophyll (Table 3 and 4) were recorded (Azmat et al., 2005a, b).
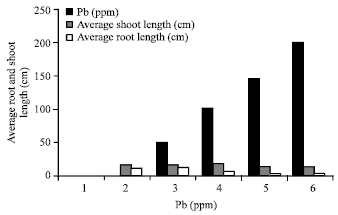 | |
| Fig. 1: | Effect of Pb on root and shoot length of plant of Phaseolus mungo |
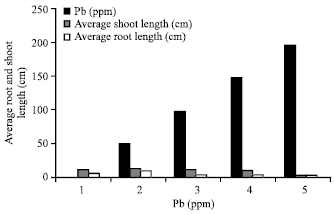 | |
| Fig. 2: | Effect of Pb on root and shoot length of plant of Lens culinaris |
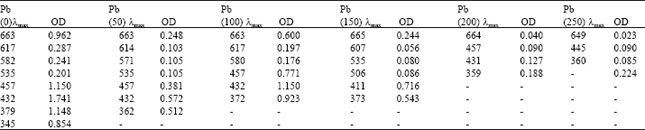
| Table 1: | Effect of Pb (ppm) on pigments absorbance and wavelength in Phaseolus mungo |
 | |
| Table 2: | Effect of Pb (ppm) on pigments absorbance and wavelength in Lens culinaris |
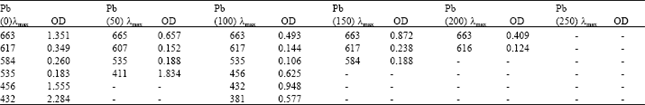 | |
| Key: λ = Wavelength, OD = Optical density, λ = 663 = Chlorophyll, 617 = Xanthophyll, 583 - 587 = Isocarotene, 530 - 430 = Caroteoids | |
| Table 3: | Concentration (mg g-1) of pigments of Phaseolus mungo under Pb stress |
 | |
| Table 4: | Concentration (mg g-1) of pigments of Lens culinaris under Pb stress |
 | |
| Table 5: | Effect of concentration of Pb on carbohydrate contents of Phaseolus mungo and Lens culinaris |
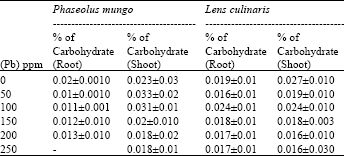 | |
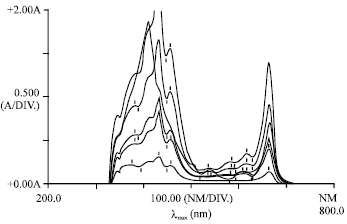
Figure 3 and 4 shows decrease in concentration of chloroplast pigments in the absorption spectrum of both Phaseolus mungo and Lens culinaris with change in wavelength. It may be due to the accumulation of Pb in prophyrin ring of chlorophyll due to which photosynthesis was not effective because light energy absorbed by accessory pigment is transferred to chlorophyll then it activates photosynthesis.
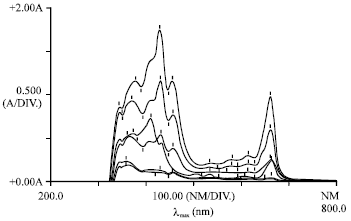 | |
| Fig. 3: | Spectral change of chlorophyll and other pigments of Phaseolus mungo with respect to change in concentration of Pb |
 | |
| Fig. 4: | Spectral change of chlorophyll and other pigments of Lens culinaris with respect to change in concentration of Pb |
 | |
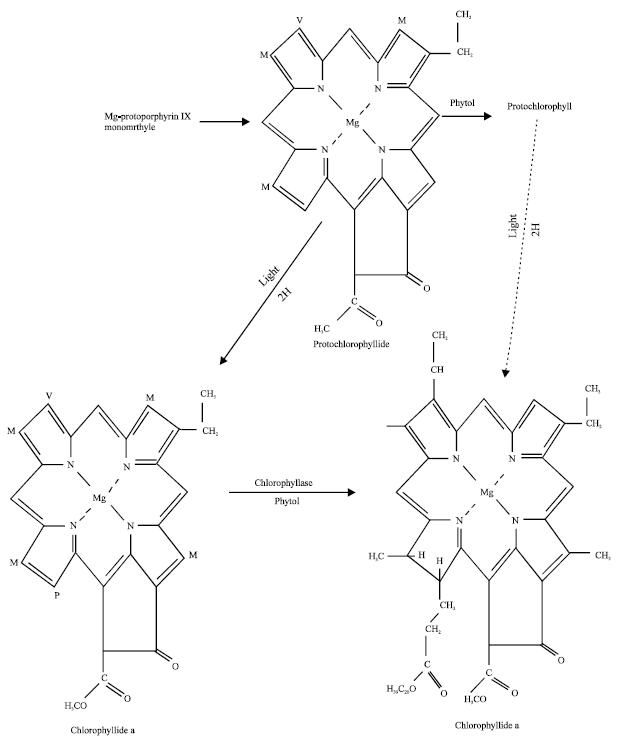 | |
| Fig. 5: | Biosynthesis of chlorophyll in presence of Pb shows the replacement of Mg ion |
The above investigation reveals that rate of photosynthesis were decline in red region of spectrum (Ambrosio et al., 1992) that is in the red absorption band of chlorophyll a. It may reduced the quantum yield of oxygen which is released after the quanta of light absorbed and this reduction in quantum yield of oxygen may effect the environment. Decrease in photosynthesis also reduced the absorption of CO2 and retardation of water molecule which causes the decrease in carbohydrate concentration in both species (Table 5). The dark green color of leaves may relate with the increase in the chlorophyll content at 150 ppm of Pb but reduced leaves size at higher concentration with compact root structure indicated that concentration of Mg and Fe in chlorophyll were reduced due to which inhibition of process of photosynthesis and leaf cells of such plants exhibited a reduction in grana stacks together with a reduction in the amount of stomata in relation to the Lamellar system as well as absence of starch grains. Table 5 shows the low carbohydrate concentration in presence of Pb which also support the above findings. Pb inhibits Chlorophyll b synthesis causing impaired uptake of essential elements such as Mg and Fe (Kowalewska et al., 1987). An enhancement of chlorophyllase activity of chlorophyll b is reported to be more affected than chlorophyll a by dose of Pb.
A strong relationship (Gitelson et al., 2003, Merzlyak et al., 2003) existed between Pb application and a decrease in photosynthesis of the whole plant and is believed to result from stomata closure rather than a direct effect of Pb on the process of photosynthesis. According to Kosobrukhov et al. (2004), the photosynthetic activity of plant is governed by many factors including stomatal cell size, number of stomata conductant, leaf area etc. While studying the effects of Pb on the development of thylakoid (Sarvari et al., 2002.) of cucumber and poplar plants. Savari et al. (2002) observed increased chlorophyll content either in PS OO core or LHC II at low concentrations of Pb treatment, whereas a strong decrease in chlorophyll level of seedlings was seen at the 50 ppm Pb treatment level. The concentration of Pb inside the leaf might have been high enough to directly inhibit chlorophyll synthesis (Sengar and Panday, 1996).
Investigation shows that spectral properties of photosynthetic pigment (Rai et al., 1999) alter in all treated plants where total chlorophyll contents diminished and efficiency of photosynthetic apparatus may be distorted by changes in pigment contents. Table 3 and 4 shows that maximum pigments were observed in control plant, which gets reduced at 250 ppm of Pb concentration. This indicated the visual non-specific symptoms of Pb toxicity as root growth inhibition, reduced leaf size and chlorosis.
Mechanism: A mechanism (Nechushtai et al., 1996; Devlin and Witham, 2001) has been proposed based on above findings which show the replacement of Mg from the porphyrin ring of chlorophyll which may be due to the higher entropy of activation of Pb as compared to Mg ion Fig. 5. Pb accumulation (Samardakiewicz and Wozny, 2000) in the roots follow the same pattern of absorption as of essential mineral ion in the roots. Photosynthetic unit is a group of collaborating molecules of pigments, which is essential for photochemical phase. Mechanism showed that when a quantum of light is absorbed and then it migrates through these collaborating molecules to the reaction center or the trapping center, with release of an electron (Nechushtai et al., 1996). This quantum of light may be used for chemical work that is, the generation of assimilatory power, ATP and NADPH but reduced leaves sized due to Pb shows decrease in surface area for photosynthesis and inhibition of chloroplast activity which also reduced the quantum yield of oxygen by following important steps in leaves.
Absorption spectrum of treated plants shows the change in spectral properties of pigments as compared to the control plants where maximum absorption was reported (0.962 for Phaseolus mungo and 1.352 for Lens culinaris) at λmax 663. This change in spectrum revealed the possibility of Pb substitution of Mg in chlorophyll leading to longer shift of wavelength (Gitelson et al., 2003; Merzlyak et al., 2003). Absorption spectrum of chloroplast pigment shows that Pb may interact indirectly with carotenoids pigments (Frank and Cogdell, 1996) affecting carbohydrate environment of photosynthetic antenna. It may relate with the effect of heavy metals on the photosynthetic apparatus with special attention paid to chlorophyll which are much more complex than just simple paid to substitution by other metals for central Mg ion in the porphyrin ring. The probable mechanism shows that if optical density is higher at 150 ppm of Pb, it indicated the maximum accumulation of metal in chlorophyll ring due to which maximum absorption of light took place but further enhancement of Pb shows the decrease in concentration of chlorophyll because leaves size (Williams et al., 1989) were reduced and no growth of seedling were observed.
CONCLUSION
Pb has considerable attention as a pollutant heavy metal pollutant due to the growing anthropogenic pressure on the environmental. Pb contaminated soils show a sharp decline line in crop productivity mainly through roots.
REFERENCES
- Ambrosio, D., K. Szabo and H.K.L. Ichtenthaler, 1992. Increase of the chlorophyll fluorescence ratio F690/F735 during the autumnal chlorophyll breakdown. Rad. Environ. Biophys., 31: 51-62.
CrossRef - Azmat, R., Zill-e-Huma, A. Hayat, T. Khanum and R. Talat, 2005. The inhibition of bean plant metabolism by cadmium metal I effect of Cd metal on physiological process of bean plant and Rhizobium species. Pak. J. Biol. Sci., 8: 401-404.
CrossRefDirect Link - Azmat, R., Y. Akhtar, R. Talat and F. Uddin, 2005. The inhibition of bean plant metabolism by CD metal and atrazine: I. The effect of atrazine with CD metal on growth, photosynthesis, nutritional level and rhizosphere of soil. Biotechnology, 4: 238-242.
CrossRefDirect Link - Baryla, A., P. Carrier, F. Frank, C. Coulmb, C. Sahut and M. Havaux, 2001. Leaf chlorosis in oilseed rape plants (Brassice napus) grown on cadmium-polluted soil: Causes and consequences for photosynthesis and growth. Planta, 212: 696-709.
Direct Link - Bohlar, N. and H.G. Elisabeth, 1987. Determination of total chlorophyll distribution pattern in leaves. Photosyn. Res., 12: 13-23.
CrossRef - Ewais, E.A., 1997. Effect of cadmium nickel and lead on growth chlorophyll content and proteins of weeds. Biol. Plant, 39: 403-410.
CrossRef - Frank, H.A. and R.J. Cogdell, 1996. Carotenoids in photosynthesis. Photochem. Photobiol., 63: 257-264.
CrossRefDirect Link - Gitelson, A., A. Zur, Y.O.B. Chivkunova and M.N. Merzlyak, 2002. Assessing carotenoid in plant leaves with reflectance spectroscopy. Photochem. Photobiol., 75: 272-281.
Direct Link - Hartmut, K.L., R. Hak and U. Rinderle, 1990. The chlorophyll fluorescence ratio F690/F730 in leaves of different chlorophyll content. Photosyn. Res., 25: 295-598.
CrossRef - Kosobrukhov, A., I. Knyazeva and V. Mudrik, 2004. Plantago major plants responses to increase content of lead in soil: Growth and photosynthesis. Plant Growth Regul., 42: 145-151.
Direct Link - Gitelson, A.A., Y. Gritz and M.N. Merzlyak, 2003. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol., 160: 271-282.
CrossRefDirect Link - Merzlyak, M.N., A.A. Gitelson, O.B. Chivkunova, A.E. Solovchenko and S.I. Pogosyan, 2003. Application of reflectance spectroscopy for analysis of higher plant pigments. Russian J. Plant Physiol., 50: 704-710.
CrossRefDirect Link - Sengar, R.S. and M. Pandey, 1996. Inhibition of chlorophyll biosynthesis by lead in greening Pisum sativum leaf segments. Biol. Plant, 38: 459-462.
CrossRef - Williams, K., F.B. Christopher and H.A. Moony, 1989. Relationships among leaf construction cost leaf longevity and light environment of the genus piper. Am. Natr., 133: 198-211.
CrossRef