
Wessam Mohamad Abdel-Wahab
Department of Zoology, Faculty of Science, University of Alexandria, Egypt
Journal of Pharmacology and Toxicology
Year: 2015 | Volume: 10 | Issue: 2 | Page No.: 60-70







ABSTRACT
Cyclosporine A (CsA) is a powerful immunosuppressant used for the management of transplant rejection as well as the treatment of several autoimmune diseases. Despite its effectiveness, nephrotoxicity still a main complication of CsA treatment. The present study investigated the therapeutic efficacy of thymoquinone (TQ) and/or selenium (Se) against CsA nephrotoxicity in rats. CsA (25 mg kg–1, po) was administered to rats for 21 days while TQ (10 mg kg–1, po) and/or Se (0.2 mg kg–1, po) were given for 7 days before CsA and continued through the 21 days of CsA administration. CsA nephrotoxicity was evidenced by marked elevation in the level of urea, creatinine and uric acid in serum. The levels of total protein and albumin were decreased. Significant increase in renal lipid peroxidation along with decreased levels of both enzymatic and non-enzymatic antioxidants were also major features for CsA nephrotoxicity. Supplementation with TQ or Se to CsA-intoxicated animals improved the functioning of the kidney and restored its oxidative/antioxidative homeostasis. Furthermore, a combination therapy of both was more effective in mitigating the above mentioned disturbances than each monotherapy. In conclusion, joint supplementation with TQ and Se may be a new approach for management of renal complications associated with clinical use of CsA.
PDF Abstract XML References Citation
Received: April 29, 2015;
Accepted: July 24, 2015;
Published: October 12, 2015
How to cite this article
Wessam Mohamad Abdel-Wahab, 2015. Therapeutic Efficacy of Thymoquinone and Selenium Against Cyclosporine A Nephrotoxicity in Rats. Journal of Pharmacology and Toxicology, 10: 60-70.
DOI: 10.3923/jpt.2015.60.70
URL: https://scialert.net/abstract/?doi=jpt.2015.60.70
DOI: 10.3923/jpt.2015.60.70
URL: https://scialert.net/abstract/?doi=jpt.2015.60.70
INTRODUCTION
Nephrotoxic drugs have been implicated in up to 25% of cases of renal failure. Cyclosporine A (CsA), a fungus-derived polypeptide, is a pivotal immunosuppressive agent that has revolutionized the management of allograft transplantation and autoimmune disorders (Ponticelli, 2005). However, the full clinical use of CsA has not been completely realized due to its frequent toxicity in particular for the kidney (Burdmann et al., 3003). Nephrotoxic effects of CsA include renal afferent vasoconstriction and reduction in glomerular filtration rate, increased renal vascular resistance and tubular atrophy mainly in the proximal tubules (Jackson et al., 1987; Kaye et al., 1993; Justo et al., 2003). Reactive oxygen species (ROS) are highly active and have the ability to react with many intracellular molecules including lipids, proteins and nucleic acids. The generation of ROS has been implicated in a variety of renal diseases (Lopez-Novoa et al., 2011). Excessive production of free radicals was suggested to be one of the main mechanisms involved in the pathogenesis of CsA nephrotoxicity (De Hornedo et al., 2007). Cyclosporine A has been shown to increase the generation of ROS and to enhance membrane lipid peroxidation (LPO) which results in the release of vasoconstrictors such as endothelin and thromboxane, finally leading to impairment in the kidney functioning (Parra et al., 1998; Hagar, 2004). Cyclosporine A also impairs the antioxidant defence system in the renal cortex (Mohamadin et al., 2005). Therefore, the therapeutic course of CsA together with a powerful antioxidant therapy may be an appropriate approach to eliminate or at least reduce the toxic side effects of CsA in the kidney.
Thymoquinone (TQ) has been identified as the main bioactive constituent in the volatile oil of Nigella sativa seeds. It possesses strong antioxidant properties and has been shown to increase the level of antioxidant enzymes such as glutathione peroxidase, glutathione reductase and catalase (Badary et al., 2003). It also acts as a cleaner by neutralizing oxygen radical anions. Thymoquinone protected against oxidative damage induced by a variety of free radical-generating agents including doxorubicin-induced cardiotoxicity and carbon tetrachloride-evoked hepatotoxicity (Nagi et al., 1999; Nagi and Mansour, 2000). The protective effect of TQ against nephrotoxicity and renal injury has also been reported (Basarslan et al., 2012).
Micronutrients are required for the human body in a very small quantity. They serve as prosthetic group in active sites or as co-enzymes for indispensable metalloenzymes. Among antioxidant micronutrients, Selenium (Se) is a fundamental trace element that plays an important role in number of physiological processes, including elimination of ROS and modulation of redox-sensitive enzyme cascades (Jihen et al., 2009). It has the ability to protect the structure and function of proteins, DNA and chromosomes against oxidative injury (Reddy et al., 2009). Evidences suggested that many selenoproteins, which contain Se in the form of selenocysteine, have important functions associated with antioxidant activity (De Freitas and Rocha, 2011). Low Se level was reported to be a frequent observation in patients with chronic kidney disease. Furthermore, Se supplementation in cases of kidney failure was found to reduce the products of oxidative stress (Iglesias et al., 2013).
Amelioration of renal damage induced by CsA will be beneficial regarding the safety of this drug. Recently, a new strategy in modern therapy has emerged which is the use of combination therapy instead of monotherapy. Therefore, the current study was undertaken to evaluate the therapeutic efficacy of TQ and Se either individually or collectively in alleviating CsA-induced nephrotoxicity and oxidative stress in rats.
MATERIALS AND METHODS
Drugs and chemicals: Cyclosporine A (Sandimmune, 100 mg) was obtained from Novartis Pharmaceuticals. Selenium (in the form of sodium selenite, Na2SeO3) and thymoquinone (2-isopropyl-5-methyl-1, 4-benzoquinone) were obtained from Sigma-Aldrich Co., USA. All other chemicals and reagents used in the current study were of analytical grade. Kits are commercially available and were obtained from Biodiagnostic Co. (Egypt) and Randox Laboratories (UK).
Animals and experimental design: A total of forty eight adult male Sprague-Dawley rats (initially weighing 210-230 g) were used in the current study. They were obtained from the Medical Research Institute, University of Alexandria (Alexandria, Egypt). They were housed in stainless steel cages (4-5 rats/cage) under standard laboratory conditions (temperature 24±3°C, humidity 40-50%, 12 h dark/light cycle). During the experiment, rats were fed standard rat food and given water ad libitum. All animal procedures were conducted in conformation with the Ethical Guidelines for the Use and Care of laboratory animals. An adaptation period of 2 weeks was allowed before onset of the experiment then rats were randomly divided into eight groups (six rats in each). The first group served as vehicle control and received olive oil; the second group (CsA group) received cyclosporine A (25 mg kg–1 day–1 dissolved in olive oil, post-orally) for 21 days; the third group (TQ group) received thymoquinone (10 mg kg–1 day-1 dissolved in olive oil, post-orally); the fourth group (Se group) received selenium (0.2 mg kg–1 day–1 dissolved in distilled water, post-orally); the fifth group (TQ+Se group) received thymoquinone and selenium (10 and 0.2 mg kg–1 day–1, respectively); the sixth group (CsA+TQ group) received cyclosporine A and thymoquinone (25 and 10 mg kg–1 day–1, respectively); the seventh group (CsA+Se group) received cyclosporine A and selenium (25 and 0.2 mg kg–1 day–1, respectively); the eighth group (CsA+TQ+Se group) received cyclosporine A, thymoquinone and selenium (25, 10 and 0.2 mg kg–1 day–1, respectively). Thymoquinone and/or Se were given for 28 days (starting 7 days before CsA administration and continued concurrently with CsA for 21 days).
Serum collection and preparation of kidney homogenate: On completion of the experimental duration (28 days), animals were killed after 24 h of last doses by cervical decapitation under light ether anaesthesia. Blood samples were left to clot at room temperature and centrifuged at 3000 rpm for 20 min. Sera were then separated and stored at -20°C for biochemical analysis. The abdominal cavities of the anesthetized rats were immediately opened and the kidneys were excised, trimmed of fats, washed with normal saline to remove blood and blotted between filter papers. For preparation of homogenate, 1 g of renal tissue was homogenized in 10 mL phosphate-buffered saline (PBS, pH 7.0 containing 1 mM EDTA) and centrifuged at 5000 rpm for 30 min. The collected supernatant was then kept at -20°C for further analysis.
Biochemical assays
Assessment of serum specific markers related to renal function: The level of creatinine, urea and uric acid were assayed spectrophotometrically according to the standard procedures using commercially available diagnostic kits.
Determination of serum level of total protein and albumin: The level of total protein was measured as described by Lowry et al. (1951) using bovine serum albumin as standard. Level of albumin was determined depending on its quantitative binding to bromocresol green to form a colored complex that absorbs at 578 nm (Doumas et al., 1971).
Determination of lipid peroxidation (LPO) level in renal tissue: The LPO level in kidney homogenate was measured as thiobarbituric acid reactive species (TBARS) depending on the reaction of TBARS with thiobarbituric acid (TBA) to form a colored complex that absorbs at 532 nm (Ohkawa et al., 1979). The specific activity is expressed as nmol mg–1 protein.
Determination of renal total antioxidant capacity (TAC): The TAC was measured in serum using the commercial kits according to the modified Miller and Rice-Evans method (Miller and Rice-Evans, 1996). This assay depends on the ability of antioxidants in the sample to inhibit the oxidation of ABTS (2, 2’-azino-bis (3-ethylbenzthiazoline-6-sulfonic acid)) to ABTS+ by metmyoglobin. The amount of ABTS+ produced can be measured spectrophotometrically at 405 nm. Troloxä serves as a standard or control antioxidant. Results are expressed as mmol equivalent Troloxä L–1.
Measuring reduced glutathione (GSH) content in renal tissue: Renal GSH content was determined as described by Ellman (1959). GSH reacts with Ellman’s reagent (5, 5-dithio-2-nitrobenzoic acid) in phosphate buffer (pH 8.0) and the absorbance was measured at 412 nm. Results are expressed as μmol mg–1 protein.
Determination of enzymatic antioxidant activity in renal tissue: The activity of glutathione peroxidase (GPx) was determined as described by Paglia and Valentine (1967). GPx catalyzes the oxidation of GSH to form oxidized glutathione which in then reduced by glutathione reductase and NADPH forming NADP+ and recycling GSH. The decrease in the absorbance (A340) is directly proportional to the GPx activity. Results were expressed in U mg–1 protein. The activity of glutathione s-transferase (GST) was determined by monitoring the conjugation of 1-chloro-2, 4-dinitrobenzene (CDNB) with GSH, absorbance was measured at 340 nm. The rate of increase is directly proportional to the GST activity in the sample (Habig et al., 1974). Results are expressed as μmol mg–1 protein. Catalase (CAT) activity was measured according to the method described by Aebi (1984). Hydrolysis of hydrogen peroxide was assayed spectrophotometrically at 240 nm in 50 mM phosphate buffer (pH 7). The specific activity of catalase was expressed as nmol mg–1 protein. The superoxide dismutase (SOD) activity in renal tissue was analyzed using nitroblue tetrezolium (NBT) as a substrate and phenazine methosulphate. The color intensity of the chromogen produced was measured at 560 nm. Results are expressed as U mg–1 protein (Kakkar et al., 1984).
Statistical analysis: Data are presented as mean±SE for each experimental group (n = 6). Statistical significance was analyzed using SPSS programme (Statistical Package for Social Science, version 16). For comparison, One-way Analysis of Variance (ANOVA) test was carried out followed by Tukey’s Range test for post-comparison. Statistical significance was acceptable at a level of p≤0.05.
RESULTS
Effect on kidney function biomarkers: As depicted in Table 1, significant increases (p≤0.05) in the level of serum biomarkers related to kidney function were recorded in CsA intoxicated rats. The level of urea, creatinine and uric acid were found to be increased by 193.9, 262.2 and 118.9% respectively when compared with the control group. Supplementation with TQ at a dose of 10 mg kg–1 significantly (p≤0.05) reduced the levels of these markers by 43.9, 39.8 and 38.7%, respectively when compared with the CsA group. Similarly, Se supplementation at a dose of 0.2 mg kg–1 significantly (p≤0.05) reduced these biomarkers by 38.1, 33.5 and 33.4%, respectively compared to the CsA group.
| Table 1: | Effect of CsA, TQ, Se and their different combinations on the level of urea, creatinine and uric acid in the control and different experimental groups |
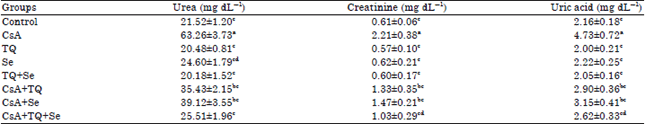 | |
| Each value represents the Mean±SE for each experimental group (n = 6), p≤0.05 for ANOVA. Mean values within the same column not sharing a common superscript letter (a, b, c and d) are significantly different. CsA: Cyclosporine A, TQ: Thymoquinone, Se: Sodium selenite | |
| Table 2: | Changes in the level of total proteins and albumin in the serum of the control and the different experimental groups |
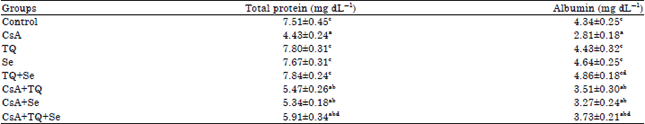 | |
| Each value represents the Mean±SE for each experimental group (n = 6), p≤0.05 for ANOVA. Mean values within the same column not sharing a common superscript letter (a, b, c and d) are significantly different. CsA: Cyclosporine A, TQ: Thymoquinone, Se: Sodium selenite | |
Furthermore, a combination therapy of both TQ and Se significantly (p≤0.05) reduced the levels of urea, creatinine and uric acid (by 59.6, 53.4 and 44.6%, respectively) more effectively than each monotherapy alone. No significant changes in kidney markers were observed in rats administered TQ and/or Se alone compared with the normal control indicating their safety at the selected doses (Table 1).
Effect on the level of total protein and albumin in serum: Table 2 shows the effect of CsA on serum total protein and albumin in the control and different experimental groups. Administration of CsA (25 mg kg–1) for 21 successive days induced a significant (p≤0.05) decrease by 40% and 35.3% in the level of total protein and albumin respectively. Pretreatment of CsA-intoxicated rats with TQ and/or Se for 7 days before CsA then concurrently during the course of CsA improved to some extent the level of total protein and albumin in serum although their values still show statistical significant difference when compared with the control group value (Table 2).
Effect on LPO, TAC and GSH: The data presented in Table 3 shows significant changes in the level of LPO, TAC and GSH content during treatment of rats with CsA, TQ and Se alone or in combinations. The results showed a significant increase (p≤0.05) by 131.4% in the level of LPO (measured as TBARS) in kidney of rats treated with CsA when compared with the control. Pretreatment with TQ or Se or a combination of both was very effective in reducing the elevated LPO level in CsA-intoxicated rats (40.7, 28 and 50%, respectively). Exposure to CsA caused significant decrease (p≤0.05) in serum total antioxidant capacity (-81%) and GSH content (-51%) in kidney tissue of rats. Monotherapy with TQ or Se alone showed recovery of the renal total antioxidant capacity and GSH content in rats exposed to CsA. Furthermore, a combined therapy of both TQ and Se was more powerful in restoring the renal total antioxidant capacity and GSH content when compared with each monotherapy alone. No statistically significant difference (p≤0.05) in TBARS, TAC and GSH in groups treated only with TQ and/or Se when compared with the control group (Table 3).
Effect on GPx, GST, SOD and CAT activities in renal tissue: According to Table 4, the administration of CsA (25 mg kg–1) for 21 successive days suppressed the antioxidative status of the kidney. It significantly (p≤0.05) decreased the activity of the enzymatic antioxidants GPx (-61.4%), GST (-43.4%), CAT (-52.1%) and SOD (-40.6%). Rats treated with CsA and supplemented with either TQ or Se alone showed significant increases in the activity of GPx, GST, SOD and CAT when compared with the CsA group. Furthermore, it was noticed that combined treatment with TQ and Se (group 8) provoked maximum protection and to a large extent improved the antioxidative status of the kidney as compared with individual treatment groups (groups 6 and 7).
| Table 3: | Effect of CsA, TQ, Se and their different combinations on the level of TBARS, TAC and GSH in the control and the different experimental groups |
 | |
| Each value represents the Mean±SE for each experimental group (n = 6), p≤0.05 for ANOVA. Mean values within the same column not sharing a common superscript letter (a, b, c and d) are significantly different. CsA: Cyclosporine A, TQ: Thymoquinone, Se: Sodium selenite, TBARS: Thiobarbituric acid reactive species, TAC: Total antioxidant capacity, GSH: Reduced glutathione | |
| Table 4: | Changes in the activity of GPx, GST, CAT and SOD in renal tissue of the control and the different experimental groups |
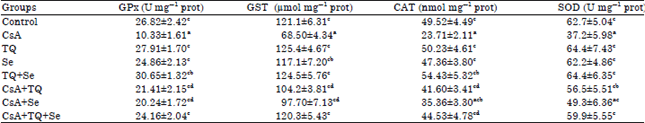 | |
| Each value represents the Mean±SE for each experimental group (n = 6), p≤0.05 for ANOVA. Mean values within the same column not sharing a common superscript letter (a, b, c and d) are significantly different. CsA: Cyclosporine A, TQ: Thymoquinone, Se: Sodium selenite, GPx: Glutathione peroxidase, GST: Glutathione S-transferase, CAT: Catalase, SOD: Superoxide dismutase | |
It is worth to note that the administration of TQ and/or Se to normal rats for 28 successive days showed no significant changes in the activity of these antioxidant enzymes (Table 4).
DISCUSSION
Over the past decade, CsA has been increasingly applied with considerable clinical benefit in management of organ transplantation and treatment of autoimmune diseases. However, its clinical uses are often accompanied by serious side effects, mostly nephrotoxicity. The precise mechanisms of CsA-induced nephrotoxicity are not completely known. Cumulative data pointed out to the involvement of excessive production of ROS in the pathogenesis of CsA nephrotoxicity (Parra et al., 1998; Hagar, 2004). This assumption was supported by the fact that using free radical scavengers provided a marked functional and structural protection against CsA-induced renal damage (Shin et al., 2002; Hagar et al., 2006). The present study provided further evidence for the involvement of oxidative stress in CsA nephrotoxicity. It also demonstrated the effectiveness of either TQ or Se in alleviating CsA-induced renal dysfunction and oxidative stress especially when administered in combination which points out to their synergistic protective effect.
Serum creatinine level is important for diagnosing first phases of renal failure while urea and uric acid levels are significant for the detection of late renal tissue damage. Uric acid and creatinine can be used as a rough index for the glomerular filtration rate. The data obtained in the current study indicated a significant increase in the level of creatinine, urea and uric acid in the serum of rats treated with CsA as compared with control group. These results point out to serious nephrotoxicity and renal functional impairment following CsA administration. This observation runs in consistency with those reported in earlier studies (Tirkey et al., 2005; Khan et al., 2006). ROS may be involved in altered glomerular filtration rate. CsA was found to increase the production of oxygen radicals which can alter the renal function through modulating the renal blood flow and the glomerular filtration rate (Capasso et al., 2008). Also, CsA causes perturbation in the renal vasoconstriction-vasodilation balance by increasing vasoconstrictor and reducing vasodilator leading to renal vasoconstriction and decreased renal blood flow (Parra et al., 1998). Monotherapy of TQ or Se in the present study antagonized the renal dysfunction induced by CsA as indicated by the decline in the level of urea, creatinine and uric acid when compared with the CsA group. Furthermore, they provided a more powerful protective effect when administered in combination compared with monotherapies of TQ or Se. These results demonstrated a clear nephroprotective action for each monotherapy and pointed out to their synergistic effect. Previous studies demonstrated similar protective effect for TQ (Sayed-Ahmed and Nagi, 2007) and Se (Randjelovic et al., 2012) against nephrotoxicity induced by other nephrotoxic agents which support the results of the current study. This effect may be related to the free radical scavenging properties of TQ and Se against ROS involved in vasoconstriction and the impairment in glomerular filtration rate as mentioned above.
Results from the present study revealed a significant decrease in the level of total protein and albumin in serum after 21 days of CsA administration when compared with control group which is in agreement with Jeon and Kim (2011). The observed decrease in protein content may be attributed to the reduced protein synthesis and/or increased proteolytic activity. CsA is hepatotoxic and its hepatotoxicity includes inhibition of hepatic protein synthesis, probably at the translation level (Buss et al., 1988). Free radicals are considered to be a major source for DNA damage. They can attack DNA molecule causing strand break and base alteration (Trivedi et al., 2008). Therefore, the reduction in protein content recorded herein may be an indirect effect of CsA through excessive production of free radicals which in turn cause DNA and RNA damage. In this study, CsA-induced alterations in protein content were ameliorated with the administration of Se and TQ either individually and collectively although their levels did not show full recovery when compared with the control group. The ability of Se and TQ to improve the protein content could be ascribed to their ability to suppress CsA-induced oxidative damage in the liver with subsequent recovery of the hepatic synthetic function.
Oxidative stress has been proposed to contribute to nephrotoxicity. Excessive production of ROS is the central key in the mechanisms involved in the pathogenesis of renal failure (Capasso et al., 2008). Data obtained in the present study showed a remarkable increase in the level of LPO (measured as TBARS) in rats treated with CsA which further supports the link between oxidative stress and CsA nephrotoxicity. Several lines of evidence suggested a similar correlation both in vivo and in vitro (Zhong et al., 1998; Chen et al., 2002; Anjaneyulu et al., 2003). CsA or its metabolites have been shown to enhance the production of O2, H2O2 and OH. radicals which are expected to accelerate LPO of biomembranes (Parra et al., 1998). The pathological consequence of this process is reflected in the impairment of membrane integrity and membrane-associated functions. Increased LPO can be counteracted by the use of antioxidants. Therefore, the use of antioxidants that scavenge or interfere with the production of ROS may be a successful approach to alleviate CsA nephrotoxicity. In the present study, a significant decline in the level of TBARS in CsA-treated rats was achieved with the administration of TQ or Se. In addition, supplementation with both agents in combination more effectively attenuated LPO in the kidney tissue. The protective effect of TQ and/or Se against CsA-induced oxidative stress in this study could be a direct effect by scavenging free radicals and inhibiting LPO or indirect through the enhancement of the antioxidant defense system in renal tissue. The notable suppressive effect of TQ against LPO may be related to direct inhibition of free radicals generation. Selenium was found to be able to disrupt the propagation of the free radical reactions and LPO in kidney tissue (Ognjanovic et al., 2008). Furthermore, it is incorporated into selenoenzymes which play an important role against free radicals (Chen and Berry, 2003).
Cells are provided with an efficient endogenous antioxidants, either non-enzymatic (as GSH) or enzymatic (as GPx, GST, CAT and SOD), to combat free radicals. Decrease in renal antioxidant capacity due to excessive ROS production makes it more susceptible to oxidative damage. In the present study, a marked decline in serum total antioxidant capacity, renal GSH content and renal GPx, GST, CAT and SOD activities were noted in rats treated with CsA. The suppression of the renal antioxidant defense system following CsA administration observed herein supports previous studies (Mohamadin et al., 2005; Josephine et al., 2006). These results points out to the consumption of the endogenous antioxidants during counteracting the enhanced free radical production (as evidenced by increased TBARS). Furthermore, the reduction of GSH content and the activities of the antioxidant enzymes could be attributed to the direct effect of CsA on the radical quenching enzymes and/or due to depletion of the enzyme substrates themselves. Studies suggested that treatment with CsA depletes GSH content in kidney tissue which seems to be a prime factor in LPO (Badary et al., 2005). Depletion of renal GSH could account for the inhibition of GPx and GST activities and consequently the CAT activity as CAT could compensate for the depletion of GPx by scavenging peroxide radicals (Kayanoki et al., 1996). Modulating the antioxidant defense machinery by agents has become a part of many therapeutic strategies. In the current study, supplementation of CsA-intoxicated rats with TQ or Se restored the renal antioxidative homeostasis. As well, a combination of both was found to be more efficient in improving the renal antioxidant status which may be closely related to their ability to enhance the intracellular defense system. The ability of TQ to preserve the antioxidant defense system may be partially due to preventing the depletion of intracellular GSH and/or stimulating the recovery of the antioxidant enzymes in kidney tissues (Sayed-Ahmed and Nagi, 2007). Furthermore, TQ was found to elevate the activity of these antioxidant enzymes through up-regulation of the GST, GPx and CAT genes (Ismail et al., 2010). Selenium helps to maintain the glutathione antioxidant system since it is incorporated into selenoproteins, which are important to antioxidant enzymes. Ognjanovic et al. (2008) and Randjelovic et al. (2012) reported that Se protected against renal toxicity through the restoration of antioxidant status of the kidney.
CONCLUSION
Cyclosporine A impaired the proper functioning of the kidney and induced disturbance in the renal oxidant/antioxidant homeostasis. Pretreatment with TQ or Se significantly improved the kidney functioning and restored its cellular antioxidant defense system. Furthermore, in combination, TQ and Se appear to be more effective than monotherapies which points out to their synergistic protective effect against CsA-induced nephrotoxicity and oxidative stress. Their beneficial effects as antioxidants imply the involvement of free radicals in CsA-induced nephrotoxicity and indicate their ability to decrease lipid peroxidation and potentiate the antioxidant defense system in a rat model.
REFERENCES
- Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Anjaneyulu, M., N. Tirkey and K. Chopra, 2003. Attenuation of cyclosporine-induced renal dysfunction by catechin: possible antioxidant mechanism. Renal Failure, 25: 691-707.
CrossRefDirect Link - Badary, O.A., R.A. Taha, A.M.G. El-Din and M.H. Abdel-Wahab, 2003. Thymoquinone is a potent superoxide anion scavenger. Drug. Chem. Toxicol., 26: 87-98.
CrossRefPubMedDirect Link - Badary, O.A., S. Abdel-Maksoud, W.A. Ahmed and G.H. Owieda, 2005. Naringenin attenuates cisplatin nephrotoxicity in rats. Life Sci., 76: 2125-2135.
CrossRefDirect Link - Basarslan, F., N. Yilmaz, S. Ates, T. Ozgur and M. Tutanc et al., 2012. Protective effects of thymoquinone on vancomycin-induced nephrotoxicity in rats. Hum. Exp. Toxicol., 31: 726-733.
CrossRefDirect Link - Burdmann, E.A., T.F. Andoh, L. Yu and W.M. Bennet, 2003. Cyclosporine nephrotoxicity. Semin. Nephrol., 23: 465-476.
CrossRefDirect Link - Buss, W.C., J. Stepanek and W.M. Bennett, 1988. Proposed mechanism of cyclosporine toxicity: Inhibition of protein synthesis. Transpl. Proc., 20: 863-867.
PubMed - Capasso, G., C.I. Di Gennaro, F.D. Ragione, C. Manna and R. Ciarcia et al., 2008. In vivo effect of the natural antioxidant hydroxytyrosol on cyclosporine nephrotoxicity in rats. Nephrol. Dial. Transplant, 23: 1186-1195.
CrossRefPubMedDirect Link - Chen, H.W., C.T. Chien, S.L. Yu, Y.T. Lee and W.J. Chen, 2002. Cyclosporine A regulate oxidative stress-induced apoptosis in cardiomyocytes: Mechanisms via ROS generation, iNOS and Hsp70. Br. J. Pharmacol., 137: 771-781.
CrossRefDirect Link - Chen, J. and M.J. Berry, 2003. Selenium and selenoproteins in the brain and brain diseases. J. Neurochem., 86: 1-12.
CrossRefPubMedDirect Link - De Freitas, A.S. and J.B.T. Rocha, 2011. Diphenyl diselenide and analogs are substrates of cerebral rat thioredoxin reductase: A pathway for their neuroprotective effects. Neurosci. Lett., 503: 1-5.
CrossRefDirect Link - De Hornedo, J.P., G. de Arriba, M.C. Fernandez, S. Benito and T.P. Cid, 2007. Cyclosporine A causes oxidative stress and mitochondrial dysfunction in tubular renal cells. Nefrologia, 27: 565-573.
Direct Link - Doumas, B.T., W.A. Watson and H.G. Biggs, 1971. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta, 31: 87-96.
CrossRefPubMedDirect Link - Ellman, G.L., 1959. Tissue sulfhydryl groups. Arch. Biochem. Biophys., 82: 70-77.
CrossRefPubMedDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Hagar, H.H., 2004. The protective effect of taurine against cyclosporine A-induced oxidative stress and hepatotoxicity in rats. Toxicol. Lett., 151: 335-343.
CrossRefPubMedDirect Link - Hagar, H.H., E. El Etter and M. Arafa, 2006. Taurine attenuates hypertension and renal dysfunction induced by cyclosporine A in rats. Clin. Exp. Pharmacol. Physiol., 33: 189-196.
CrossRefPubMedDirect Link - Iglesias, P., R. Selgas, S. Romero and J.J. Diez, 2013. Selenium and kidney disease. J. Nephrol., 26: 266-272.
Direct Link - Ismail, M., G. Al-Naqeep and K.W. Chan, 2010. Nigella sativa thymoquinone-rich fraction greatly improves plasma antioxidant capacity and expression of antioxidant genes in hypercholesterolemic rats. Free Radical Biol. Med., 48: 664-672.
CrossRefDirect Link - Jackson, N.M., C.H. Hsu, G.E. Visscher, M.A. Venkatachalam and H.D. Humes, 1987. Alterations in renal structure and function in a rat model of cyclosporine nephrotoxicity. J. Pharmacol. Exp. Ther., 242: 749-756.
Direct Link - Jeon, Y.J. and Y.S. Kim, 2011. Cyclosporine A inhibits albumin synthesis in Huh7 cells. Korean J. Internal Med., 26: 314-319.
CrossRefDirect Link - Jihen, E.H., M. Imed, H. Fatima and K. Abdelhamid, 2009. Protective effects of selenium (Se) and zinc (Zn) on cadmium (Cd) toxicity in the liver of the rat: Effects on the oxidative stress. Ecotoxicol. Environ. Saf., 72: 1559-1564.
CrossRefDirect Link - Josephine, A., C.K. Veena, G. Amudha, S.P. Preetha and P. Varalakshmi, 2006. Evaluating the effect of sulphated polysaccharides on cyclosporine a induced oxidative renal injury. Mol. Cell. Biochem., 287: 101-108.
CrossRefDirect Link - Justo, P., C. Lorz, A. Sanz, J. Egido and A. Ortiz, 2003. Intracellular mechanisms of cyclosporine A-induced tubular cell apoptosis. J. Am. Soc. Nephrol., 14: 3072-3080.
CrossRefDirect Link - Kakkar, P., B. Das and P.N. Viswanathan, 1984. A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem. Biophys., 21: 130-132.
PubMedDirect Link - Kayanoki, Y., J. Fujii, K.N. Islam, K. Suzuki, S. Kawata, Y. Matsuzawa and N. Taniguchi, 1996. The protective role of glutathione peroxidase in apoptosis induced by reactive oxygen species. J. Biochem., 119: 817-822.
Direct Link - Kaye, D., J. Thompson, G. Jennings and M. Esler, 1993. Cyclosporine therapy after cardiac transplantation causes hypertension and renal vasoconstriction without sympathetic activation. Circulation, 88: 1101-1109.
CrossRefDirect Link - Khan, M., J.C. Shobha, I.K. Mohan, M.U.R. Naidu, A. Prayag and V.K. Kutala, 2006. Spirulina attenuates cyclosporine-induced nephrotoxicity in rats. J. Applied Toxicol., 26: 444-451.
CrossRefDirect Link - Lopez-Novoa, J.M., Y. Quiros, L. Vicente, A.I. Morales and F.J. Lopez-Hernandez, 2011. New insights into the mechanism of aminoglycoside nephrotoxicity: An integrative point of view. Kidney Int., 79: 33-45.
CrossRefPubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mohamadin, A.M., H.A. El-Beshbishy and M.A. El-Mahdy, 2005. Green tea extract attenuates cyclosporine A-induced oxidative stress in rats. Pharmacol. Res., 51: 51-57.
CrossRefPubMedDirect Link - Nagi, M.N., K. Alam, O.A. Badary, O.A. Al-Shabanah, H.A. Al-Sawaf and A.M. Al-Bekairi, 1999. Thymoquinone protects against carbon tetrachloride hepatotoxicity in mice via an antioxidant mechanism. Biochem. Mol. Biol. Int., 47: 153-159.
PubMed - Nagi, M.N. and M.A. Mansour, 2000. Protective effect of thymoquinone against doxorubicin-induced cardiotoxicity in rats: A possible mechanism of protection. Pharmacol. Res., 41: 283-289.
CrossRefDirect Link - Ognjanovic, B.I., S.D. Markovic, S.Z. Pavlovic, R.V. Zikic, A.S. Stajn and Z.S. Saicic, 2008. Effect of chronic cadmium exposure on antioxidant defense system in some tissues of rats: Protective effect of selenium. Physiol. Res., 57: 403-411.
PubMedDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Paglia, D.E. and W.N. Valentine, 1967. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med., 70: 158-169.
CrossRefPubMedDirect Link - Parra, T., G. de Arriba, J.R. Conejo, M. Cantero and I. Arribas et al., 1998. Cyclosporine increases local glomerular synthesis of reactive oxygen species in rats: Effect of vitamin E on cyclosporine nephrotoxicity. Transplantation, 66: 1325-1329.
PubMedDirect Link - Ponticelli, C., 2005. Cyclosporine: From renal transplantation to autoimmune diseases. Ann. New York Acad. Sci., 1051: 551-558.
CrossRefDirect Link - Randjelovic, P., S. Veljkovic, N. Stojiljkovic, L. Velickovic, D. Sokolovic, M. Stoiljkovic and I. Ilic, 2012. Protective effect of selenium on gentamicin-induced oxidative stress and nephrotoxicity in rats. Drug Chem. Toxicol., 35: 141-148.
CrossRefDirect Link - Reddy, K.P., G. Sailaja and C. Krishnaiah, 2009. Protective effects of selenium on fluoride induced alterations in certain enzymes in brain of mice. J. Environ. Biol., 30: 859-864.
PubMedDirect Link - Sayed-Ahmed, M.M. and M.N. Nagi, 2007. Thymoquinone supplementation prevents the development of gentamicin-induced acute renal toxicity in rats. Clin. Exp. Pharmacol. Physiol., 34: 399-405.
CrossRefPubMedDirect Link - Shin, Y.H., S.H. Lee and K.C. Mun, 2002. Effect of melatonin on the antioxidant enzymes in the kidneys of cyclosporine-treated rats. Transpl. Proc., 34: 2650-2651.
CrossRefPubMedDirect Link - Tirkey, N., G. Kaur, G. Vij and K. Chopra, 2005. Curcumin, a diferuloylmethane, attenuates cyclosporine-induced renal dysfunction and oxidative stress in rat kidneys. BMC Pharmacol., Vol. 5.
CrossRef - Trivedi, M.H., R.J. Verma and N.J. Chinoy, 2008. Amelioration by black tea of sodium fluoride-induced effects on DNA, RNA and protein contents of liver and kidney and on serum transaminase activities in Swiss albino mice. Fluoride, 41: 61-66.
Direct Link - Zhong, Z., G.E. Arteel, H.D. Connor, M. Yin and M.V. Frankenberg et al., 1998. Cyclosporine A increases hypoxia and free radical production in rat kidneys: Prevention by dietary glycine. Am. J. Physiol.-Renal Physiol., 275: F595-F604.
Direct Link