
Houehanou M.A.G. Gbaguidi
Laboratoire d`Ecologie et de Management des Ecosyst�mes Aquatiques (LEMEA), Departement de Zoologie, Faculte des Sciences et Techniques, Universite d`Abomey-Calavi, B.P. 526 Cotonou, Benin
Alphonse Adite
Laboratoire d`Ecologie et de Management des Ecosyst�mes Aquatiques (LEMEA), Departement de Zoologie, Faculte des Sciences et Techniques, Universite d`Abomey-Calavi, B.P. 526 Cotonou, Benin
Youssouf Abou
Laboratoire d`Ecologie et de Management des Ecosyst�mes Aquatiques (LEMEA), Departement de Zoologie, Faculte des Sciences et Techniques, Universite d`Abomey-Calavi, B.P. 526 Cotonou, Benin
Journal of Fisheries and Aquatic Science
Year: 2017 | Volume: 12 | Issue: 1 | Page No.: 42-53







ABSTRACT
Background and Objective: The trophic ecology of the silver catfish, Chrysichthys nigrodigitatus (Lacepède, 1803) was examined to evaluate resource exploitation and utilization and the establishment of this highly priced species, introduced in the man-made lake (pond) of Ahozon (South-Benin). Methodology: Chrysichthys nigrodigitatus individuals were sampled monthly in both open water and aquatic vegetation habitats from August, 2014 to October, 2015 and stomach contents were evaluated on each specimen collected. Results: The eco-morphological patterns depicted along with dietary analysis showed that C. nigrodigitatus is an opportunistic benthic feeder foraging mainly on aquatic insects (59.9%), substrate particles (11.76%), detritus (9.18%), seeds (5.51%) and algae (4.97%). High niche breadths ranging between 0.55-8.35 were recorded and niche overlaps (0.39<Øjk<0.98) between size classes indicated ontogenetic diet shifts. The trophic plasticity behavior of this claroteid and the broad range of preys ingested, led to active spawning and high colonization that indicated the successful establishment of C. nigrodigitatus in lake Ahozon. Conclusion: The success of fisheries and its sustainable exploitation require a holistic approach of lake management including the prevention of domestic waste dumping, the protection of spawning and foraging grounds, the planned harvest of the fish stock, the protection of lake’s margin to avoid erosion and the periodic ecological monitoring of lake Ahozon.
PDF Abstract XML References Citation
Received: November 11, 2016;
Accepted: December 12, 2016;
Published: December 15, 2016
How to cite this article
Houehanou M.A.G. Gbaguidi, Alphonse Adite and Youssouf Abou, 2017. Trophic Ecology and Establishment of the Silver Catfish, Chrysichthys nigrodigitatus (Pisces: Siluriformes: Claroteidae) Introduced in an Artificial Pond of Benin, West Africa. Journal of Fisheries and Aquatic Science, 12: 42-53.
DOI: 10.3923/jfas.2017.42.53
URL: https://scialert.net/abstract/?doi=jfas.2017.42.53
DOI: 10.3923/jfas.2017.42.53
URL: https://scialert.net/abstract/?doi=jfas.2017.42.53
INTRODUCTION
The silver catfish, Chrysichthys nigrodigitatus (Lacépède, 1803) is among the dominant African commercial fishes of high economic value and widely serves as food for human consumption in West Africa1,2. Chrysichthys nigrodigitatus is a prominent member of the claroteids and occurred abundantly in some Benin inland waters (Lake Nokoué, lake Ahémé, Costal lagoons, Porto-Novo lagoon etc.) where this species made about 7-26% of the total annual fish catches3,4.
This catfish belongs to the genus Chrysichthys, family Claroteidae, Siluriformes order and Ostariophysi super order. Synonimies are Pimelodus nigrodigitaus, Arius acutivelus, Chrysichthys furcatus, Chrysichthys buettikoferi and Chrysichthys cameronensis5. In general, the species showed grew silvery color with a white belly and a black adipose6. Chrysichthys nigrodigitatus exhibited a pointed snout slightly longer than or equal to the width of the mouth and the pre-maxyllary tooth plate width made 20-30% of the head length5. In lake Nokoué and Porto-Novo lagoon of Southern Benin, the sizes at first sexual maturity of C. nigrodigitatus were 30.3 cm TL and 29.3 cm TL for males and females, respectively7. In the tropical estuary of Nigeria, Asuquo et al.8 reported the species as foraging on a variety of benthic food resources.
Though widespread in the natural inland waters where the species grows and spawns actively, it is quite difficult to reproduce C. nigrodigitatus in fish pond in order to supply aquaculture center with fingerlings (seeds). Therefore, the numerous man-made lakes, established throughout the country are alternative environment mediums that may contribute to aquaculture development and particularly, to serve as spawning grounds for some delicate species such as C. nigrodigitatus. In December, 2011, about 265 live individuals (15 genitors and 250 figerlings) of C. nigrodigitatus has been introduced by the fishermen in the sand-dragged man-made lake of Ahozon (South-Benin) where the species, currently, made about 11.22% of the fish assemblages and hence constituted in this artificial lake, the second dominant species after the cichlid Sarotherodon galilaeus9.
Notwithstanding the economic/commercial importance of this catfish and its relative high abundance in lake Ahozon, knowledge on the bio-ecology and particularly on the feeding ecology of C. nigrodigitatus in this man-made lake is not documented. In particular, nothing is known about the feeding habits, resource exploitation and utilization and foraging patterns of this claroteid in the sand-dragged artificial lake of Ahozon. Fisheries ecologists have been conducted studies on feeding ecology to evaluate the dietary composition and the food habits of a fish species in order to provide useful information for fisheries management, aquaculture valorization and habitat protection. As reported by Gbaguidi et al.9, the success of fisheries management, species conservation and aquaculture development requires knowledge on trophic ecology of the target species. Evaluating the prominence and establishment of a species in a man-made lake requires extensive qualitative and quantitative data on the exploitation and utilization of the food resources considered as energy sources for species survival, growth and active spawning10.
The current study investigated the feeding ecology of C. nigrodigitatus in order to document resource exploitations and species establishment in the sand-dragged man-made lake of Ahozon. The output will contribute to habitat protection, species conservation and fisheries/aquaculture valorizations of C. nigrodigitatus in the numerous man-made lakes of Benin.
MATERIALS AND METHODS
Study site: The study site is the artificial lake of Ahozon (Ouidah city, South-Benin) located between 06°22’52"N and 002°10’34"E and covering about 0.1658 km2 (Fig. 1a). Lake Ahozon is an abandoned and unmanaged water body originated from the accumulation of running waters in a hole created by sand-dragging activities9. The study area comprises a vast wetland at the costal area that extended from Cotonou city (South-Benin) to Ouidah, some sandy soils and red ferric soils9. The region experiences a sub-equatorial climate with two wet seasons (April-July, mid-September-October) with a peak usually recorded in June and two dry seasons, a major one from December-March and the minor one from mid-August to mid-September11. Yearly rainfall averaged 1307.3 mm and ambient temperatures ranged between 25 and 33.6°C. Monthly evaporation varied between 59.2 and 145 mm11.
With regard to water quality, lake Ahozon exhibited dissolved oxygen concentrations ranging between 0.73 and 11.8 mg L–1 (mean: 5.43 mg L–1), percentages of dissolved oxygen saturation between 10.5 and 208.8% (mean: 82.9%) and conductivity averaging 240 μ cm–1 (range: 50-560 μ cm–1). Average pH was 7.51 (range: 6.7-9.7) and water temperatures varied between 28.2 and 38.7°C (mean: 33.25°C). Also, depths ranged between 16.2-240 cm (mean: 80.19 cm) and transparencies averaged 36.63 cm (range: 16.2-60.5 cm)9.
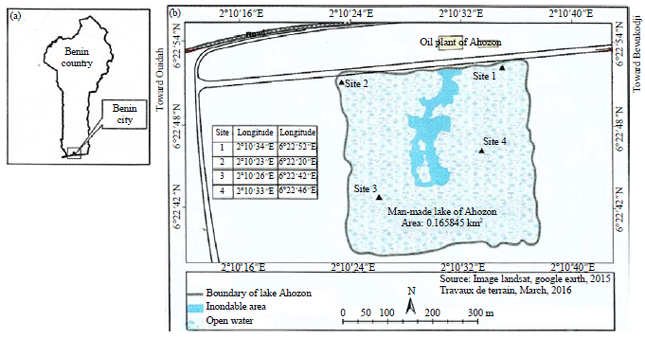 | |
| Fig. 1(a-b): | Map showing (a) Ouidah city, the study region and (b) Lake Ahozon along with the sampling sites |
The fish community of lake Ahozon comprised the cichlids, Sarotherodon galilaeus, Oreochromis niloticus and Tilapia guineensis, the African bonytongue, Heterotis niloticus (Osteoglossidae), the African catfish, Clarias gariepinus (Clariidae) and the claroteid, Chrysichthys nigrodigitatus9. Benthic macro invertebrates recorded were chironomid larvae and a gasteropod mollusk, Melanoïde tubercularis. Zooplanktons genera recorded in lake Ahozon were Trichocerca, Keratella, Brachionus and copepods. Aquatic vegetation mainly comprised Cyperus crassipes, Cyperus rotundus, Fuirena umbellata and Ropogon gayanus, Ludwigia perennis, Emilia praetermissa, Eleocharis complanata, Enydra fluctuans, Mariscus ligularis and dominant phytoplankton included Navicula, Peridinium, Scenedemus, Pinnularia, Spirogyra, Cosmarium, Melosira, Synechocystis, Microcystis, Oscillatoria, Euglena, Phacus, Surirella and Lychmophora. The water of lake Ahozon was occasionally utilized for subsistence agriculture. Likewise, subsistence multi species fisheries sporadically occurred in lake Ahozon and were practiced by a couple of migrant fishermen12.
Sampling sites: To document the feeding ecology of C. nigrodigitatus in the man-made lake of Ahozon, four sampling locations were considered, two sites in the "Open water" habitat and two sites in the "Aquatic vegetation" habitat (Fig. 1b). The "Open water" habitat showed a relatively high depth and high water velocity, but exempt of vegetation whereas the "Aquatic vegetation", the lake’s edge, was shallow and characterized by relatively dense vegetation and low water velocities.
Fish collection: Chrysichthys nigrodigitatus individuals were collected twice a month during 15 consecutive months (August, 2014-October, 2015). Samplings were performed both in the "Aquatic vegetation" habitat and in the "Open water" habitat using cast net (9.80 m-diameter, 4.90 m height, 40 mm mesh), seine (4.15 m length×1.77 m width, 3 mm mesh), hooks (90 m length) and experimental gill nets (40×1.05 m, 40 mm mesh)11. Seining was done in the marginal aquatic vegetation by setting the seine stationary and kicking the vegetation to drive the fish in to the net12,13. Cast nets, gill nets and hooks were used in the "Open water". After collection, fish samples were preserved in 10% formalin and then transported to the "Laboratoire d’ Ecologie et de Management des Ecosystèmes Aquatiques". In the laboratory, the fish samples were removed from the formalin and preserved in 70% ethanol to facilitate manipulations14.
Dietary analysis: The ethanol preserved-fishes were identified using the inland water fish guide of Van Thielen et al.6 and the identification keys of Leveque et al.5. Chrysichthys nigrodigitatus individuals were sorted by habitat and by sampling period15. Each individual was then weighed to the nearest 0.1 g with an electronic balance (Philips) and measured for Total Length (TL) and Standard Length (SL) to the nearest 0.1 mm with an ichtyo meter.
Chrysichthys nigrodigitatus individuals were then dissected and the digestive tract was removed and its length was measured as the distance from the distal end to the anus16. The alimentary canal was then opened and all food resources were removed and spread on a glass slide for examination first under a binocular (model: Pierron) to identify large food items. Water was added to facilitate separation and identification of small food items. To identify algae, a few drops of water containing fine prey items from stomach contents were examined under a photonic microscope10,12. Prey items were identified to the lowest possible taxonomic level using the reference of Needham and Needham17 for algae and macro invertebrates. The identified food items were then separated and blotted on a paper towel to remove excess moisture. The volume of each food category from an individual stomach was estimated by water displacement using an appropriately sized-graduated cylinder12. The prey items belonging to a given category were gathered into a single sample for volumetric estimates. For small prey items (<0.002 mL), volume estimations were made by spreading the food resource on a glass slides, then estimating visually the approximate volume of the material by comparison with a 0.01 mL drop of water delivered with a pipet onto a clean glass slide. For example, a food item having a volume approximately one quarter of the volume of the 0.01 mL water droplet was recorded as 0.0025 mL12.
Data analysis: For each of the 1020 stomachs of C. nigrodigitatus dissected, the estimated volume of each food resource was recorded on Excel software spreadsheet containing all type of prey items identified. The proportional volumetric consumption of each prey was then computed as follows10:
where, Pi is the volumetric proportion of food item i in the diet, Vi is the total volume of the food item i in n stomachs, Vt is the total volume of food ingested by n stomachs, n is the total number of stomachs examined (1020 stomachs). Seasonal (wet, flood, dry) variations in volumetric proportions were depicted with analysis of variances (ANOVA) using SPSS18. Also, to explore ontogenetic diet shifts, the volumetric proportions of each food resource ingested were computed for different size classes of C. nigrodigitatus. Niche breadth was calculated following Simpson’s formula10:
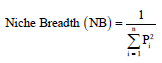 |
where, Pi is the proportion of food item i in the diet and n is the total number of food items in the diet, NB ranges from 1, when only one prey item is used to n, when all preys are ingested in equal proportions. The matrices of computed niche breadth were submitted to one-way ANOVA using SPSS software computer program18.
The coefficient of emptiness is the ratio of number of empty stomachs to total number of stomachs examined and was expressed as follows:
where, Ce is the coefficient of emptiness, Ne is the number of empty stomachs, Nt is the total number of stomachs examined. The frequency of occurrence (FO) of each prey items was computed using the following formula of Bahou et al. 19:
where, Ji is the number of stomachs containing prey item i, Nt is the total number of non-empty stomachs (Nt = 658). Also, diet similarities and ontogenetic diet shift were explored using Pianka’s niche overlap index (Øjk) computed as follows10:
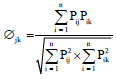 |
where, Øjk is the dietary overlap between species j and species k, Pij is the proportion of resource i used by species j, Pik is the proportion of resource i used by species k and n is the number of resource categories utilized.
To evaluate the eco-morphological patterns of C. nigrodigitatus, the linear regressions of "Gut Length (GL)×body weight (W)" and "Gut Length (GL)×Standard Length (SL)" were examined. Likewise, the ratio of SL-GL (GL/SL) was computed as a measure of relative gut length and compared to published reference ratios20,21. These eco-morphological data were used to document the food habit of C. nigrodigitatus.
RESULTS
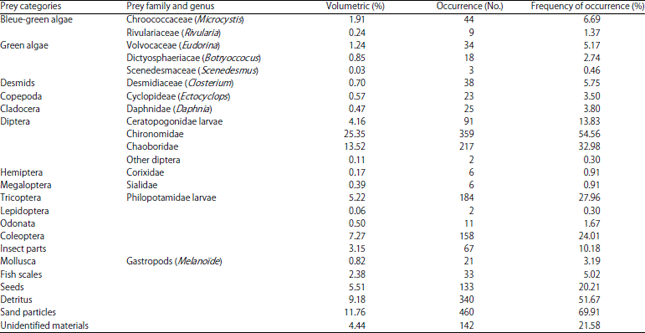
Prey ingested and niche breadth: In the artificial lake of Ahozon, C. nigrodigitatus consumed about 25 benthic and pelagic food resources dominated by aquatic insects (59.9%), substrate particles (11.76%), detritus (9.18%), seeds (5.51%) and algae (4.97%) (Table 1). Aquatic insects were dominated by diptera larvae and nymphs (43.14%) comprised of Chironomidae (25.35%), Chaoboridae (13.52%) and Ceratopogonidae (4.16%). Other major aquatic insects ingested were Coleoptera (7.27%), Tricoptera (5.22%) with Megaloptera (Sialidae), Hemiptera (Corixidae), Odonata and Lepidoptera, the minor aquatic insects consumed by C. nigrodigitatus.
Aggregated algae ingested by C. nigrodigitatus comprised six families and six genera including bleue-green algae such as Microcystis (Chroococcaceae), Rivularia (Rivulariaceae), green algae such as Eudorina (Volvocaceae), Botryoccocus (Dictyosphaeriacae), Scenedesmus (Scenedesmaceae) and a desmid, Closterium (Desmidiaceae). Minor preys (4.24%) included fish scales (2.38%), a gastropod mollusk, Melanoïde tubercularis (0.82%) and some microcrustacea such as Daphnia (0.47%), a cladoceran and Ectocyclops (0.57%) a copepod (Table 1).
In lake Ahozon, niche breadth averaged 5.53±2.78 and ranged between 0.55 for SL <80 mm and 8.35 for SL reaching 120 mm. These relatively high niche breadths recorded indicated that C. nigrodigitatus consumed a broad spectrum of food resources in the man-made lake of Ahozon. Ontogenetically, lower size (SL <80 mm) tended to show reduced niche breadths compared to sub-adults and adults (SL >80 mm) which exhibited a relatively high values between 3.64 and 8.35. In general, the niche breadth did not show any significant (p>0.05) seasonal variations and mean values during wet, flood and dry periods were NB = 7.82, NB = 7.97 and NB = 7.42, respectively.
Empty stomachs: On 1020 individuals of C. nigrodigitatus dissected, 362 stomachs corresponding to 35.49% were empty (Table 2). In general, the coefficient of emptiness (Ce) varied seasonally and ontogenetically and ranged between 0 and 58.82%. However, one-way analysis on the percentage of empty stomachs in lake Ahozon did not show significant (p>0.05) variations with the development stages (juvenile, sub-adult, adult) and season (wet, flood, dry). Indeed, the computed F-value, along with degrees of freedom and p-value was F2,8 = 1.682, p = 0.263 for life stage and F2,8 = 0.391, p = 0.692 for season, thus indicating that seasons and life stages did not affect the incidences of empty stomachs. Nevertheless, empty stomachs in adults were higher and averaged 43.07±12.11, whereas lower percentages of empty stomachs were recorded among juveniles with a mean of 20.64±19.79 (Table 2). Seasonally, higher empty stomachs were recorded during the dry periods and averaged 39.95±20.48, whereas, those of wet season were the lowest and averaged 32.29±9.74.
| Table 1: | Volumetric percentages and frequency of occurrence (%) of prey items ingested by Chrysichthys nigrodigitatus from lake Ahozon |
 | |
| Table 2: | Seasonal variations of emptiness coefficients (Ce) of Chrysichthys nigrodigitatus from lake Ahozon |
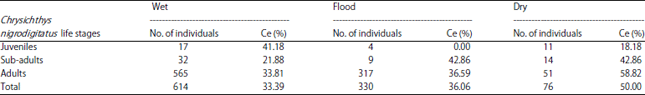 | |
| Total number of empty stomachs: 362, Wet: 205, Flood: 119, Dry: 38, Population coefficient of emptiness (%): 35.49% | |
| Table 3: | Matrix of niche overlaps (Øjk) of Chrysichthys nigrodigitatus by size classes from lake Ahozon |
 | |
Frequency of occurrence: Overall, the percentage occurrence recorded for the food resources consumed by C. nigrodigitatus in lake Ahozon ranged between 0.46 and 69.91% (Table 1). The lowest frequencies of occurrence were recorded among algae such as Scenedesmus (Scenedesmaceae) found in 3 (0.46%) stomachs, Rivularia (Rivulariaceae) occurring in 9 (1.37%) stomachs and among miscellaneous aquatic insects such Lepidoptera occurring in 2 (0.30%) stomachs, Corixidae (Hemiptera) found in 6 (0.91%) stomachs and Sialidae (Megaloptera) recorded in 6 (0.91%) stomachs. The highest occurrences were recorded for substrate particules occurring in 460 (69.91%) stomachs, detritus found in 340 (51.67%) stomachs and among aquatic insects such as Chironomidae (Diptera) larvae and nymphs occurring in 359 (54.56%) stomachs, Chaoboridae (Diptera) larvae and nymphs found in 217 (32.98%) stomachs and Philopotamidae (Trichoptera) larvae recorded in 184 (27.96%) stomachs of C. nigrodigitatus.
Seasonal variations of diet: In lake Ahozon, C. nigrodigitatus showed insignificant (p>0.05) seasonal dietary variations for sand particles, seeds and aquatic insects (Fig. 2). Indeed, the computed F and p-values from one-way analysis of variance were F2,10 = 2.674, p = 0.129 for substrate particles, F2,10 = 3.088, p = 0.101 for seeds and F2,10 = 2.686, p = 0.128 for aquatic insects. Nevertheless, the proportional consumption of sand particles were greater during the flood season with a volumetric proportion averaging 14.62±0.48, those of seeds were greater during the dry period and averaged 24.68±22.99% and those of aquatic insects were greater during the wet period with a mean of 66.97±1.02%. Inversely, C. nigrodigitatus exhibited significant (p<0.05) seasonal dietary variations for detritus and algae. The calculated F and p-values from one-way ANOVA were F2,10 = 6.323, p = 0.023 for detritus with the highest mean value (13.87±3.43%) recorded during the wet period and F2,10 = 4.390, p = 0.05 for algae with the highest mean value (9.31±3.87%) recorded during the flood period.
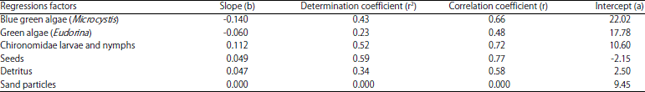
Diet similarities and ontogenetic diet shifts: Niche overlaps (Øjk) between different size classes of C. nigrodigitatus were relatively high and ranged between Øjk = 0.39 and Øjk = 0.98 (Mean: 0.72±0.23). The lowest niche overlap (Øjk = 0.39) was recorded for the similarity between the subpopulation of size category "SL <80 mm" and that of size category "SL: 120-160 mm". The highest niche overlap (Øjk = 0.98) was recorded for the similarity between the subpopulation of size category "SL: 80-120 mm" and that of size category "SL: 120-160 mm" (Table 3). In particular, the juvenile subpopulation (SL <80 mm) exhibited relatively lower diet similarities (0.39<Øjk<0.56) with subadult and adult subpopulations. Indeed, algae and microcrustacae dominated the diet of C. nigrodigitatus juvenile (SL <80 mm) whereas, aquatic insects, seeds, detritus and substrate particules relatively dominated the diet of subadults and adults (SL >80 mm). Furthermore, though not significant, the linear regression between the volumetric proportions (%) of blue green algae, green algae and Standard Length (SL) gave some negative slopes, b = -0.140 and b = -0.060, respectively, indicating that the proportional consumption of these algae decreased with SL. In contrast, the proportional consumption of seed, aquatic insects and detritus increased with Standard Length (SL) and gave reduced positive slopes b = 0.049, b = 0.112 and b = 0.047, respectively. This size-related foraging trend of C. nigrodigitatus in lake Ahozon is an indication of ontogenetic diet shifts (Table 4).
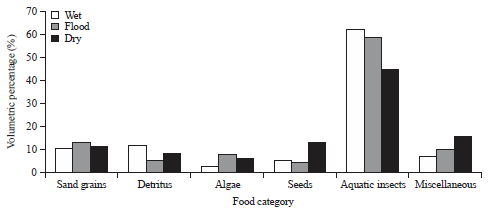 | |
| Fig. 2: | Seasonal variations of foods resources consumed by Chrysichthys nigrodigitatus from the man-made of lake Ahozon |
| Table 4: | Volumetric percentages of food resources consumed by size classes (mm SL) of Chrysichthys nigrodigitatus from lake of Ahozon |
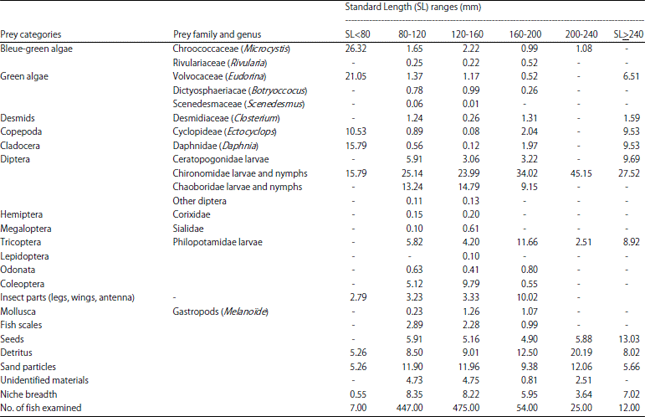 | |
| SL: Standard length | |
Food resources and morphological relationships: The proportional consumption of dominant food resources (algae, aquatic insects, seeds, detritus, sand particles) were plotted against Standard Length (SL) and Gut Length (GL) to explore the eco-morphological correlates of the trophic structure of C. nigrodigitatus. The output from linear regression relationships indicated that the volumetric proportion of blue green algae was negatively correlated with SL (b = -0.140, r = 0.66) and the volumetric proportion of green algae was negatively correlated with SL (bSL = -0.060, rSL= 0.48) and GL (bGL = -0.210, rGL = 0.84) (Table 5). Inversely, the volumetric proportion of aquatic insects, seeds and detritus were positively correlated with SL and GL indicating that the proportional consumptions of these three prey items increased with SL and GL (Table 6).
| Table 5: | Linear regression between the volumetric percentages of prey items ingested and Standard Length (SL) of Chrysichthys nigrodigitatus from lake Ahozon |
 | |
| Table 6: | Linear regression between the volumetric percentages of prey items ingested and Gut Length (GL) of Chrysichthys nigrodigitatus from lake Ahozon |
 | |
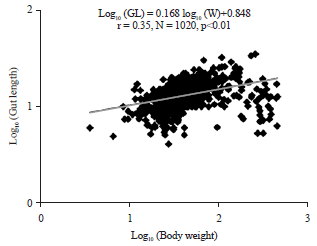 | |
| Fig. 3: | Relationship between log (body weight (W)) and log (Gut Length (GL)) of Chrysichthys nigrodigitatus from lake Ahozon |
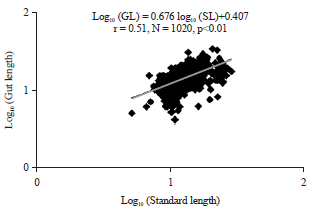 | |
| Fig. 4: | Relationship between log (Gut Length (GL)) and log (Standard Length (SL)) of Chrysichthys nigrodigitatus from lake Ahozon |
Also, the eco-morphological patterns of C. nigrodigitatus were evaluated through the linear regressions of "Gut Length (GL)×body weight (W)" and "Gut Length (GL)×Standard Length (SL)". Likewise, the ratio of SL-GL (GL/SL) was calculated as a measure of relative gut length and compared to published reference ratios20,21. Both equations showed positive slopes, b = 0.168 and b = 0.676 and were as follows:
| • | Log10 (GL) = 0.168 log10 (W)+0.848, r = 0.35 (r2 = 0.13), N = 1020, p<0.01 (Fig. 3) |
| • | Log10 (GL) = 0.676 log10 (SL)+0.407, r = 0.51 (r2 = 0.26), N = 1020, p<0.01 (Fig. 4) |
In addition, the ratio of GL/SL ranged between 0.22 (23.8 cm SL) and 2.19 (13.7 cm SL) with a mean of 1.10± 0.27.
DISCUSSION
Knowledge on trophic ecology is of great interest for habitat protection, species conservation and establishment, fisheries management and overall ecosystem valorization10,22. In the man-made lake of Ahozon, C. nigrodigitatus foraged on about 25 food resources dominated by aquatic insects (59.9%), substrate particles (11.76%), detritus (9.18%), seeds (5.51%) and algae (4.97%) (Table 1). These results indicated that, in this special habitat, aquatic insects were the major food items consumed by this introduced catfish species. In particular, diptera larvae and nymphs (43.14%) mostly composed of three items, Chironomidae (25.35%), Chaoboridae (13.52%) and Ceratopogonidae (4.16%), dominated aquatic insects. This high consumption of aquatic insects by C. nigrodigitatus was probably due not only to its food preference, but also to the high availability of insects in this artificial water body. Indeed, in lake Ahozon, the presence of marginal wetlands associated with aquatic vegetation could have boosted the proliferation of aquatic insects that were available for C. nigrodigitatus. In this study, there was no record of domestic waste dumping or organic pollution that could cause chironomid proliferations. Therefore, only the colonization and the decomposition of marginal vegetation may have increased the abundance of chironomids.
In the Cross River Estuary of Nigeria, like in lake Ahozon, aquatic insects (19.6%) were one of the major preys consumed by C. nigrodigitatus. However, fish dominated the diet of the Cross River Estuary with 21.6% whereas, fish parts remained trivial (2.38%) in lake Ahozon21. In contrast with our findings, no aquatic insects have been recorded in the diet of C. nigrodigitatus from the Epe Lagoon of Nigeria and the percentage of algae consumed by C. nigrodigitatus from this lagoon reached 78.49%, against 4.97% ingested in the artificial lake of Ahozon23. As reported by Fagade and Olaniyan24, bivalves (84%) and gastropods (14%) were the main prey items as being present in the diet of C. nigrodigitatus from Lagos lagoon of Nigeria.
These differences recorded in the diet composition could be partially attributed to the period of sampling between study regions. For example, the diet investigation in the Epe Lagoon of Nigeria extended from January-July (2008), probably corresponding to two seasons (dry, wet) whereas lake Ahozon samplings encompassed 15 months, hence, including the three seasons (wet, flood, dry). Another major cause of the differences in the diet composition between lake Ahozon population and that of the three water bodies (Lagoon of lagos, Cross River Estuary, Epe Lagoon) of Nigeria is that these later are natural, ancient and brackish aquatic biota with high biological diversity (fish, crustacean, mollusks, crabs, algae etc.) more available to C. nigrodigitatus. Inversely, lake Ahozon is a young and isolated artificial water body established about 11 years (2005) ago and hence, exhibiting relatively low food resources, except aquatic insects which may have been more available due to its high rate of colonization that has been especially enhanced by the presence of marginal aquatic vegetation. For example, bottom sampling during the study showed only one mollusk, Melanoïde tubercularis, a gastropod and no crabs and shrimps have been recorded in any of both habitats ("Open water", "Aquatic vegetation"). Nevertheless, like lake Ahozon population, detritus were consistently found in the diet of these three lakes (Lagoon of lagos, Cross River Estuary, Epe Lagoon) from Nigeria.
These feeding patterns depicted indicated that C. nigrodigitatus exhibited a relatively high trophic flexibility/plasticity that enables the species to switch from one diet to another according to habitat and food availability23,25. This trophic plasticity behavior of C. nigrodigitatus was shown by the relative high niche breadth ranging between NB = 0.55 and NB = 8.35 and averaging 5.53±2.78, much higher than those reported by Offem et al.22 in the Cross River of Nigeria where niche breadths varied between 0.80 and 0.92. This trophic plasticity is an advantage and explains why C. nigrodigitatus occurs and invades most inland waters such as rivers, estuaries, lagoons and freshwater lakes where this claroteid appears to be an important commercial and economic fisheries component.
Also, the frequencies of occurrence of the preys consumed by C. nigrodigitatus followed nearly the same patterns depicted with the volumetric percentage of the diet composition. Indeed of the 25 prey items ingested by C. nigrodigitatus, only five (aquatic insects, algae, sand particles, detritus, seeds) were dominant and consistently occurred individually in a relatively high number of stomachs. Among them, the aggregated aquatic insects (Diptera, Coleoptera, Tricoptera, Megaloptera, Hemiptera, Odonata and Lepidoptera) were the most dominant food resources and occurred in 620 (94.22%) stomachs, followed by sand particles found in 460 (69.91%) stomachs, detritus occurring in 340 (51.67%) stomachs, seeds occurring in 133 (20.21%) stomachs and aggregated algae (Microcystis, Rivularia, Eudorina, Botryoccocus, Scenedesmus, Closterium) found in 130 (19.76%) stomachs.
Overall, insignificant (p>0.05) seasonal and ontogenetic variations of empty stomachs was recorded in lake Ahozon. The high percentage (35.49%) recorded during the study period may be attributed to feeding cycle and fishing gears (hooks, gillnets) used to sample C. nigrodigitatus. Indeed, once set, hooks and gillnets spent about 12 h before follow-up and fish harvests. Consequently, digestion and feces evacuation continue to increase the number of empty stomachs. In the Ebonyi River in Southeastern Nigeria, Iyabo26 reported a percentage of empty stomachs between 8.3 and 21.4%, lower than our findings.
In lake Ahozon, proportional consumptions of sand particles, seeds and aquatic insects consumed by C. nigrodigitatus showed insignificant (p>0.05) seasonal variations. These trends may be attributed to the high availability of these food resources in the water body regardless of seasons so that C. nigrodigitatus may have unlimited access and consumed them ad libitum according to their needs and food requirements10. Inversely, like the coexisting tilapine cichlid S. galilaeus, the proportional consumptions of detritus showed significant (p<0.05) seasonal variations and higher consumption were recorded during the wet and dry periods. Indeed, during the flood season, detritus could be less available to fishes because flushed by water currents that caused unstable lake bottom and high disturbances.
In this study, the relatively high niche overlaps recorded between different size classes and varying from Øjk = 0.39 and Øjk = 0.98, suggested that C. nigrodigitatus showed high diet similarities between different life stage categories. Similar trends were recorded for the coexisting sympatric species, S. galilaeus exhibiting high diet overlaps (Øjk) ranging between Øjk = 0.77 and Øjk = 0.97). Nevertheless, the lower diet similarities (Øjk range: 0.39-0.56) recorded between juvenile subpopulation (SL <80 mm) and subadults/adults (SL >80 mm) class subpopulation indicated an ontogenetic diet shift. Indeed, because C. nigrodigitatus juveniles possess less developed digestif tracts, plancktonic live preys (algae, microcrustacae) dominated their diet whereas, macroscopic preys such as aquatic insects, seeds, detritus and substrate particules dominated the diet of sub-adults and adults.
Morphological data, such as Standard Length (SL), Weight (W), Gut Length (GL) and the proportional consumption of the food resources have been widely used by fisheries biologists and ecologists to approach both the eco-morphogical trends of the preys consumed and the food habits of a fish species10,20,21. In lake Ahozon, the linear regression between SL of C. nigrodigitatus and the volumetric proportion of blue green algae gave a negative slope (b = -0.140, r = 0.66), indicating that the proportional consumption of blue green algae decreased with SL. Similar eco-morphological trends were recorded with the volumetric proportion of green algae crossed with SL and GL that generated some slopes bSL = -0.060 and bGL = -0.210, indicating that the consumption of green algae decreased with SL and GL. In contrast, the consumption of aquatic insects, seeds and detritus were positively correlated with SL (b range: 0.047-0.112) and GL (b range: 0.046-0.217) (Table 6), indicating that the proportional consumptions of these three prey items increased with SL and GL. According to Gbaguidi et al.9, juvenile fish with less-developed digestive tract forage mostly on live food (algae, microcrustacea etc.). As the fish grows, the digestive system becomes well-developed and digestive enzyme secretions are initiated. Therefore, in contrast with juveniles, sub-adults and adults take advantage to other delicate food resources such as detritus, seeds and aquatic insects to satisfy their nutritional needs27.
With regard to C. nigrodigitatus food habit, in the current feeding ecology study, the ratio of SL-GL (GL/SL) averaged 1.10±0.27 and ranged between 0.22 and 2.19. These ratios are similar to those reported by Paugy21 for many African invertebrate feeders such as the claroteids Chrysichthys auratus (GL/SL = 1.98) and Auchenoglanis occidentalis (GL/SL = 0.88), the Amphilidae Amphilius atesuensis (GL/SL = 0.51), the mormyrid Brienomyrus brachyistus (GL/SL = 0.45), the cichlids Haplochromis adolphifrederici (GL/SL = 1.68) and Haplochromis gracilor (GL/SL = 1.92) and the Mockochidae Synodontis ocelifer (GL/SL = 2.18), Synodontis schall (GL/SL = 1.96) etc. These morphological features indicated that C. nigrodigitatus is an opportunistic invertebrate feeder that is also confirmed by the current dietary analysis. Indeed, aggregated invertebrates, mainly aquatic insects, dominated the stomach content of C. nigrodigitatus with a proportional consumption of 60.72% (Table 1).
In addition, C. nigrodigitatus exhibited an omnivorous food habits indicated by a consistent proportional consumption of five prey items, algae, seeds, detritus, sand particles and aquatic insects. As reported by Paugy21, most omnivores have a GL/SL ratio between 0 and 3 that also confirm our findings (GL/SL = 0.22-2.19) in the current study. Furthermore, the linear regression model between log (W) and log (GL) gave a positive slope b = 0.118, close to Kramer19 omnivore slope range (0.4-0.68) supporting that C. nigrodigitatus may be an omnivore. In the Cross River estuary of Nigeria, Offem et al.22 reported fish and shrimps as being the most abundant preys in the diet of this claroteid. As results, C. nigrodigitatus is a benthic and pelagic omnivore that intensively forages on invertebrates and detritus and at a relatively low degree, on algae and fishes.
The current study well-documented the feeding ecology of C. nigrodigitatus and showed that this introduced claroteid species is well-established in the man-made lake of Ahozon. Indeed, in lake Ahozon, C. nigrodigitatus intensively exploited and consumed a combination of twenty five food resources dominated by aquatic insects, detritus, sand particles, seeds and algae along with some minor preys such as copepod, cladoceran, gastropod mollusk and fish scales.
The high niche breadth recorded indicated the ability of C. nigrodigitatus to exploit and to consume a broad spectrum of available food resources to satisfy its energy and protein requirements needed to grow and to spawn28. In addition, the high degree of trophic plasticity depicted in the current study and also reported by Offem et al.22, Lawal et al.23 and Iyabo26, led to an opportunistic food habit and allowed C. nigrodigitatus to take advantage to the available prey items in the water body. As results, the combinaison of these positive dietary traits led to the perfect establishment of C. nigrodigitatus in lake Ahozon, that is indicated by the active breeding, the high condition indices and the relatively high colonization of this claroteid in lake Ahozon9,29-31. The establishment of C. nigrodigitatus coupled with that of S. galilaeus and reported by Gbaguidi et al.9 is a good indicator for the fisheries success in the man-made lake of Ahozon. The enhancement of the fisheries/aquaculture development and the sustainability of the ichthyofaunal exploitation require the prevention of domestic wastes dumping, the protection of spawning and foraging grounds, the planned harvest of the fish stock, the protection of the lake margin to avoid erosion and the periodic ecological monitoring of lake Ahozon9.
CONCLUSION
This study documented the feeding ecology of C. nigrodigitatus (Pisces: Siluriformes: Claroteidae) and its establishment in the artificial lake of Ahozon. The outputs consistently indicated that C. nigrodigitatus is an opportunistic benthic feeder that incorporated in its diet, a broad range of prey items dominated by aquatic insects, detritus, sand particles, seeds, algae and minor food resources such as copepod, cladocera and mollusks. The trophic plasticity behavior displayed by the species and the wide spectrum of preys ingested, led to active spawning, high recruitment and colonization that indicated the successful establishment of C. nigrodigitatus in the artificial lake of Ahozon. The sustainability of the fisheries/aquaculture success and fish exploitation in lake Ahozon requires a holistic approach of lake management.
ACKNOWLEDGMENTS
The "Laboratoire d’Ecologie et de Management des Ecosystèmes Aquatiques (LEMEA), Département de Zoologie, Faculté des Sciences et techniques, Université d’Abomey-Calavi" provided logistic and financial assistance. We are grateful to the numerous reviewers for their thorough peer-review of this manuscript. We sincerely thank Mr. Doukpo Célestin, the owner of lake Ahozon to allow us to conduct this investigation on this man-made water body. We are also grateful to Mr. Houessinon Geoffroy and Djihouessi Bernold for their assistance in fish sampling and laboratory works.
REFERENCES
- Adite, A. and R. van Thielen, 1995. Ecology and fish catches in natural lakes of Benin, West Africa. Environ. Biol. Fish., 43: 381-391.
CrossRefDirect Link - Laleye, P., J.C. Phillipart and P. Poncin, 1995. Biologie de la reproduction de deux especes de Chrysichthys (Siliuriforme, Bagridae) du Lac Nokoue et de la lagune de Porto-Novo au Benin. Revue Zoologie Africaine, 109: 213-224.
Direct Link - Asuquo, P.E., U.I. Enin and B.E. Job, 2013. Ontogenetic Variation in the Diet of Chrysichthys nigrodigitatus (Lacepede, 1803) in a tropical estuarine ecosystem in Nigeria. J. Fish. Aquatic Sci., 8: 172-177.
CrossRefDirect Link - Gbaguidi, H.M.A.G., A. Adite and E. Sossoukpe, 2016. Ecology and fish biodiversity of man-made lakes of southern benin (West Africa): Implications for species conservation and fisheries management. J. Environ. Protect., 7: 874-894.
CrossRefDirect Link - Adite, A., K.O. Winemiller and E.D. Fiogbe, 2005. Ontogenetic, seasonal, and spatial variation in the diet of Heterotis niloticus (Osteoglossiformes: Osteoglossidae) in the Sô River and Lake Hlan, Benin, West Africa. Environ. Biol. Fishes, 73: 367-378.
CrossRefDirect Link - Winemiller, K.O., 1990. Spatial and temporal variation in tropical fish trophic networks. Ecol. Monogr., 60: 331-367.
CrossRefDirect Link - Adite, A., K.O. Winemiller and E.D. Flogbe, 2006. Population structure and reproduction of the African bonytongue Heterotis niloticus in the So River-floodplain system (West Africa): Implications for management. Ecol. Freshwater Fish, 15: 30-39.
CrossRefDirect Link - Winemiller, K.O. and M.A. Leslie, 1992. Fish assemblages across a complex, tropical freshwater/marine ecotone. Environ. Biol. Fish., 34: 29-50.
CrossRefDirect Link - Winemiller, K.O., 1989. Ontogenetic diet shifts and resource partitioning among piscivorous fishes in the Venezuelan ilanos. Environ. Biol. Fish., 26: 177-199.
CrossRefDirect Link - Bahou, L., T. Kone, V. N'Douba, K.J. N'Guessan, E.P. Kouamelan and G.B. Gouli, 2007. Food composition and feeding habits of little tunny (Euthynnus alletteratus). Proceedings of the International Council for the Exploration of the Sea Annual Science Conference, September 17-21, 2007, Helsinki, Finland, pp: 1044-1052.
- Kramer, D.L. and M.J. Bryant, 1995. Intestine length in the fishes of a tropical stream: 1. Ontogenetic allometry. Environ. Biol. Fish., 42: 115-127.
CrossRefDirect Link - Paugy, D., 1994. [The ecology of fishes in a temporary tropical stream (Baoule, a tributary of the Upper Senegal R. in Mali): Environmental and diet adaptation]. Revue d'Hydrobiologie Tropicale, 27: 157-172, (In French).
Direct Link - Offem, B.O., Y. Akegbejo-Samsons and I.T. Omoniyi, 2008. Diet, size and reproductive biology of the silver catfish, Chrysichthys nigrodigitatus (Siluformes: Bagridae) in the Cross River, Nigeria. Rev. Biol. Trop., 56: 1785-1799.
CrossRefPubMedDirect Link - Lawal, M.O., O.J. Sangoleye and B.M. Seriki, 2010. Morphometry and diet of Chrysichthys nigrodigitatus (Lacepede) in Epe Lagoon, Nigeria. Afr. J. Biotechnol., 9: 7955-7960.
Direct Link - Facade, S.O. and C.I.O. Olaniyan, 1973. The food and feeding interrelationship of the fishes in the Lagos Lagoon. J. Fish Biol., 5: 205-225.
CrossRefDirect Link - Adite, A., A. Gbankoto, I.I. Toko and E.D. Fiogbe, 2013. Diet breadth variation and trophic plasticity behavior of the African bonytongue Heterotis niloticus (Cuvier, 1829) in the Sô River-Lake Hlan aquatic system (Benin, West Africa): Implications for species conservation and aquaculture development. Nat. Sci., 5: 1219-1229.
CrossRefDirect Link - Iyabo, U.B., 2014. Diet composition, feeding habits and condition factor of Chrysichthys nigrodigitatus in Ebonyi river (A tropical flood river system), Southeastern Nigeria. Continental J. Agric. Sci., 8: 29-37.
Direct Link - Alonso, M.K., E.A. Crespo, N.A. Garcia, S.N. Pedraza, P.A. Mariotti and N.J. Mora, 2002. Fishery and ontogenetic driven changes in the diet of the spiny dogfish, Squalus acanthias, in Patagonian waters, Argentina. Environ. Biol. Fish., 63: 193-202.
CrossRefDirect Link - Duque, A.B. and K.O. Winemiller, 2003. Dietary segregation among large catfishes of the Apure and Arauca Rivers, Venezuela. J. Fish Biol., 63: 410-427.
CrossRefDirect Link - Bowen, S.H. and B.R. Allanson, 1982. Behavioral and trophic plasticity of juvenile Tilapia mossambica in utilization of the unstable littoral habitat. Environ. Biol. Fish., 7: 354-362.
CrossRefDirect Link - Garcia-Berthou, E., 1999. Food of introduced mosquitofish: Ontogenetic diet shift and prey selection. J. Fish Biol., 55: 135-147.
CrossRefDirect Link