
Grasian Immanuel
Centre for Marine Science and Technology, Manonmaniam Sundaranar University, Rajakkamangalam-629502, Kanyakumari District, Tamilnadu, India
LiveDNA: 91.486
Journal of Fisheries and Aquatic Science
Year: 2016 | Volume: 11 | Issue: 4 | Page No.: 323-330







ABSTRACT
Objective: The present investigation was carried out to study the encapsulation of Artemia nauplii with probionts to promote the survival rate of Artemia as well to enhance the bacteriostatic effect of Artemia against shrimp pathogens. Methodology: The probionts, such as Lactobacillus acidophilus, L. sporogenes and yeast Saccharomyces cerevisiae were individually cultured, the extracellular metabolites of these probionts were extracted and the antagonistic effect of the metabolites was tested against the shrimp pathogens Vibrio cholerae and V. parahaemolyticus. The metabolites of L. acidophilus and L. sporogenes were showed a growth inhibition against V. parahaemolyticus and V. cholerae. The second instar stage of Artemia franciscana nauplii were selected for probiont encapsulation study. Results: The nauplii were encapsulated with selected probionts individually. The control nauplii were enriched with microalgae Nanochloropsis. During probionts enrichment, the survival was monitored for 24 h. The control nauplii showed less survival (89.33-90.60%) when compared with probionts (96.0-99.33%) encapsulated Artemia nauplii. After 24 h enrichment, the individual probiotic load of Artemia nauplii was determined. Then these control and probionts encapsulated nauplii were individually challenged with the pathogens V. cholerae and V. parahaemolyticus. Conclusion: The survival of Artemia nauplii was recorded till their death. During challenge test against Vibrio pathogens, A. franciscana nauplii encapsulated with control-Nanochloropsis showed higher mortality within limited time interval when compared with Artemia nauplii encapsulated with probionts.
PDF Abstract XML References Citation
Received: April 06, 2016;
Accepted: May 17, 2016;
Published: June 15, 2016
How to cite this article
Grasian Immanuel, 2016. Bioencapsulation of Brine Shrimp Artemia Nauplii with Probionts and Their Resistance Against Vibrio Pathogens. Journal of Fisheries and Aquatic Science, 11: 323-330.
DOI: 10.3923/jfas.2016.323.330
URL: https://scialert.net/abstract/?doi=jfas.2016.323.330
DOI: 10.3923/jfas.2016.323.330
URL: https://scialert.net/abstract/?doi=jfas.2016.323.330
INTRODUCTION
Disease outbreaks are being increasingly recognized as a significant constraint on aquaculture production and trade affecting the economic development of the sector in many countries. For instance, disease is now considered to be the limiting factor in the shrimp culture sub sector1,2. So far conventional approaches, such as the use of disinfectants and antimicrobial drugs have had limited success in the prevention or cure of aquatic diseases2. The massive use of antimicrobials for disease control and growth promotion in animals increases the selective pressure exerted on the microbial world and encourages the natural emergence of bacterial resistance. Not only resistant bacteria can proliferate, after an antibiotic has killed off the other bacteria, but also they can transfer their resistance genes to other bacteria that have never been exposed to the antibiotics3.
Several alternative strategies for the use of antimicrobials in disease control have been proposed and have already been applied successfully in aquaculture, such as the use of vaccines4, use of immunostimulants for the enhancement of the non-specific defense mechanisms of the host and the use of probiotic bacteria5. Fuller6 defined probiotics as "A live microbial feed supplement, which beneficially affects the host animal for improving its intestinal microbial balance". Probiotics for aquaculture are generally selected based on their ability to produce antimicrobial metabolites, however attachment to intestinal mucus is important in order to colonize with in the gut of its host.
A wide variety of commercial probiotics supplemented products are available for use in both humans and animal production systems. The majority of these consist of either single Bifidobacteria sp., Lactobacillus acidophilus as well as a variety of fungi. Health claims for these products include alterations to the ecology of the gut microflora, resulting in the suppression of pathogens. Lactobacillus acidophilus is generally beneficial and produces antimicrobial substances, such as acidolin, acidophilin, lactocidin and bacteriocin. The ability of lactobacilli to produce toxic metabolites, such as lactic acid, hydrogen peroxide and bacteriocins have been suggested as being responsible for the ability to inhibit other bacteria7.
Several well documented studies on the use of probiotics as biological control agents in the developing bivalve mollusks, crustaceans and fish were published. Studies by Gatesoupe8-10 have been made on the efficacy of Artemia nauplii in bioencapsulating bacteria and this indicates that it strongly depends on the type of bacteria used, time of exposure and status (live or dead) of the bacteria. Bioencapsulated lactic acid bacteria have been successfully introduced into turbot larvae with significant improvements in survival11. Bacteria with various characteristics have been incorporated into Artemia nauplii to orally challenge turbot larvae with a pathogenic Vibrio anguillarum strains12,13. This route has also been used to vaccinate sea bass fry14, juvenile carp15 and fish fry16.
The probiotic applications of Aeromonas media A199 was found to prevent death of the oyster Crassostrea gigas larvae when they were challenged in vivo with the pathogen Vibrio tubiashii, although A. media A199 was not able to persist more than 4 days on the host17. The administration of the probiotic strain to the larvae caused a spectacular decrease of the pathogen densities in the larvae compared to those in the larvae treated with V. tubiashii only.
Rengpipat et al.18 reported the use of a Bacillus strain S11 as a probiotic administered to the black tiger shrimp Penaeus monodon larvae fed with the Bacillus fortified Artemia had significantly shorter development times and lower disease problems than larvae reared without the Bacillus sp. (S11). After feeding for 100 days, P. monodon post larvae were challenged with the pathogenic Vibrio harveyii (D331) by immersion. Ten days later all the groups treated with Bacillus (S11) had 100% survival, whereas the control group had only 26% survival. Literature related to the above are still needed in the aquaculture point of view, especially hatchery production of quality and disease free/tolerant finfish and shell fish larvae concern. Hence, the present study was undertaken to bioencapsulate Artemia nauplii by the known probionts and also to investigate the resistant effect of probionts enriched Artemia nauplii against pathogenic Vibrio sp. by means of a challenge test.
MATERIALS AND METHODS
Isolation of probionts from food samples
Lactobacillus strains: The milk and cheese samples were aseptically collected and streaked individually on MRS agar plates having the pH range from 6-6.6. After incubation at 37°C for 24-48 h, the streaked plates were observed for different colonies. These colonies were selected and were again restreaked individually on MRS plates. The plates were incubated at 37°C for 48 h to obtain large cell colonies. Then the colonies were isolated and stored in slants and further physiological and biochemical tests were carried out to confirm the strains based on Bergy’s manual of determinative bacteriology19. Based on the results, two strains, such as L. acidophilus and L. sporongenes were identified.
Yeast strains: A loop full of grape juice was taken and streaked on Potato Dextrose Agar (PDA) plates at pH 3.5. This low pH (3.5) could prevent the growth of other microorganisms including bacteria. After 24-48 h of incubation at 35°C, the streaked plates were observed for budding colonies. Well isolated individual budding cells were selected and again restreaked on the PDA plates and were incubated at 35°C for 24-48 h for budding growth. This was carried to obtain pure colonies of yeast. The obtained colonies were then streaked in PDA slants in the test tubes and were used for further strain identification. Based on the results, only one yeast strain Saccharomyces cerevisiae was identified.
Isolation of Vibrio pathogens from infected shrimp: The diseased shrimp Penaeus indicus (White shrimp) were collected from a local shrimp farm at Rajakkamangalam, Tamilnadu, India. Tissue samples were aseptically taken from infected areas and mashed with mortar and pestle. These samples were streaked on the plates containing TCBS agar. The plates were incubated at 32°C for 24 h. Then well isolated colonies were selected and restreaked on the same agar plates and were incubated at 32°C for 24 h. Based on the colour and morphology, different colonies were isolated and streaked in TCBS slants for further species level identification19, accordingly two Vibrio strains like V. cholarae and V. parahaemolyticus were identified.
Antagonistic study: Probiotic organisms usually possess antagonistic activity against the pathogens in the intestinal region. The isolated probionts were checked for possessing antagonistic activity. For this, the following methods were performed.
Preparation of extracellular metabolite: The stock Lactobacillus (L. acidophilus and L. sporogenes) cultures were inoculated in 100 mL MRS broth20 individually and the yeast (S. cerevisiae) culture was inoculated in YPD broth. These cultures were incubated at 37°C for 48 h in a shaker at 250 rpm. After 48 h, the cultures were taken and centrifuged at 10000 rpm for 15 min. The culture supernatant contains the extra cellular metabolites produced by the organisms were individually filtered through membrane filter (0.45 μm pore size). Then the filtrates were extracted with ethyl acetate at pH 7 and concentrated in a vacuum rotary evaporator.
Antibacterial activity testing: At first, sterile Muller Hinton agar plates were prepared, sterile cotton swabs were dipped in the individual cell suspension of the pathogens (V. cholerae and V. parahaemolyticus) and were evenly swabbed in separate plates. The surface was allowed to dry for 5 min. In each plate, four wells were cut at a size of 5 mm diameter. Then the wells were marked and filled (20 μL) with, respective concentrated extract (metabolite). Simultaneously, the fourth well was treated as control and it was filled with 20 μL sterile water. Then the plates were incubated at 32°C for 24-48 h. After incubation, the zone of inhibition was measured (mm level). Triplicate plates were maintained for each organism.
Encapsulation of brine shrimp Artemia nauplii using probionts
Artemia cyst hatching procedure: Cysts of brine shrimp Artemia franciscana (Great Salt lake strain) were purchased from San Francisco Bay Brand (San Francisco, California, USA) and allowed to hatch under optimum hatching conditions (35 ppt salinity, 27±1°C temperature and 1000 lux light) in the laboratory. After 15-20 h of incubation, nauplii were hatched out, these nauplii were transferred separately into a 50 L capacity fibre glass tank for 24 h to attain II instar stage.
Preparation of probiotic enrichment diets: Pure cultures of the bacteria L. acidophilus and L. sporogenes were individually inoculated in MRS broth and incubated at 37°C for 24-48 h under continuous agitation. Similarly, the yeast (S. cerevisiae) culture was inoculated in Potato Dextrose (PD) broth and cultured at room temperature (25-28°C) under continuous agitation. The concentration of the bacterial and yeast cultures were determined by counting total CFU on MRS and PD agar plates, respectively, after incubation.
Artemia nauplii enrichment procedure: The II instar stage of Artemia nauplii were separated from the hatching container using 120 μm sieve and transferred to 5 L capacity glass container with fresh sea water (35 ppt). To determine experimental treatment concentration, the Artemia nauplii were fed with enrichment media, such as yeast (SC), L. sporogenes (LS) and L. acidophilus (LA) with different concentrations like 102, 103 and 104 CFU mL–1. The control Artemia nauplii were fed with microalgae-Nanochloropsis with the same concentrations21. The Artemia nauplii were stocked at the rate of 20 mL–1 in total volume of 200 mL sterilized seawater in 250 mL capacity glass containers and aerated continuously. The enrichment media were delivered in two doses at 12 h intervals. The enrichment duration was 24 h. After 24 h, the survival percentage of Artemia nauplii was assessed in each concentration and treatments. Simultaneously, triplicates were maintained for all concentrations and treatments.
Assessment of bacterial load in enriched Artemia nauplii: After 24 h of enrichment, 1 mL well mixed samples (enriched nauplii, with counts varying from 13-20 mL–1) from 103 concentration alone was taken in sterilized vials of 5 mL capacity from each enrichment medium for bacteriological samplings. During bacteriological sampling, nauplii were rinsed with sterile saline water (18 ppt) and known volume of nauplii was homogenized in 5 mL sterile saline water. Serial dilutions (10–1 to 10–6) of the homogenate in sterilized saline water were made and 1 mL of diluted samples from the last three dilutions, such as 10–4, 10–5 and 10–6 were poured on plates containing individual media namely Sabouraud Dextrose Agar (SDA) plates for enumerating yeasts, Seawater agar (SWA) plates for enumerating total aerobic flora, Man Rogosa Sharp (MRS) agar for enumerating Lactobacillus sp. and thiosulphate citrate bile salt sucrose (TCBS) plates for enumerating Vibrios. SDA and MRS plates were incubated at 37°C for 24-48 h, whereas SWA and TCBS plates were incubated at room temperature (25-28°C) for 16-20 h. Triplicates were maintained for each dilution and each medium CFUs were counted after incubation for each replication21.
Pathogenic challenge test: After enrichment, control and treatment nauplii were challenged with already isolated shrimp pathogens, such as V. parahaemolyticus and V. cholerae. The challenge was made at a rate of 5 enriched Artemia nauplii mL–1 in a total of 200 mL water taken in glass bowls in triplicate, without aeration. Vibrio parahaemolyticus and V. cholerae were grown in nutrient broth with NaCl enrichment (3%) and upto 108 CFU mL–1 was obtained within 24 h. The concentration of pathogens dispensed was 5.25 CFU mL–1 (V. parahaemolyticus) and 5.75 CFU mL–1 (V. cholerae) in each challenge bowl. The survival was monitored and counted by sampling the nauplii every 24 h interval till all nauplii had died. The Cumulative Mortality Index (CMI) was calculated by summing the mortality counts noted at each time interval.
where D is the number of dead individuals at the time, ×1,×2,×3, …,×n. The higher the CMI value, the lower the pathogenic resistance22,23.
Statistical analysis: The results obtained in the present study were expressed as Mean±SD and were analyzed with Regression analysis and also one-way ANOVA test with post hoc multiple comparison of SNK test at a significant level of 5% using a computer software STATSTICA 06 (Statosoft, Bedford, UK).
RESULTS
Antagonistic effect of metabolites of probionts against Vibrio sp.: Metabolites of L. acidophilus and L. sporogenes showed a maximum inhibitory effect against both Vibrio spp. with the zone size between 10.33-12.0 mm. But S. cerevisiae showed a maximum zone of inhibition against V. cholerae (9.33 mm) and minimum (7.80 mm) against V. parahaemolyticus (Table 1).
Survival of Artemia nauplii after enrichment studies: After 24 h of enrichment, the survival of A. franciscana nauplii was noted, it showed that the control nauplii enriched with algae Nanochloropsis displayed the survival of 90.60±0.57% at 104 concentration. However in the experimental groups, the survival was significantly (p<0.05) increased with maximum of 99.33±0.57% recorded in nauplii enriched with the probiont S. cerevisiae at 104 concentration, followed by L. acidophilus and L. sporogenes with 99.00±1.00 and 98.66±1.15%, respectively at the same concentration (Fig. 1).
Assessment of bacterial load in enriched Artemia nauplii: The mean aerobic flora counted on SWA plates of control group was high (36.33±2.08×10–4 CFU mL–1), whereas it was significantly (p<0.05) reduced, i.e., 75% less in S. cerevisiae medium enriched Artemia nauplii (9.33±1.15×10–4 CFU mL–1). At the same time, it was 66% (12.33±1.52×10–1 CFU mL–1) and 44% (20.33±2.08×10–1 CFU mL) reduction in L. sporogenes and L. acidophilus, respectively.
| Table 1: | Antagonistic effect of extracellular metabolites produced by probionts against Vibrio pathogens using agar well diffusion method |
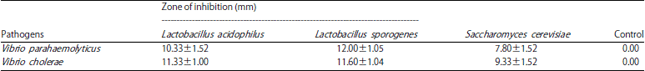 | |
| Each value is the Mean±SD of triplicate analysis | |
| Table 2: | Mean bacterial count of Aerobic flora (SWA), Vibrios (TCBS), yeast (SDA) and Lactobacillus (MRS) in A. franciscana nauplii enriched with 10-3 concentration of probionts SC, LS, LA and control |
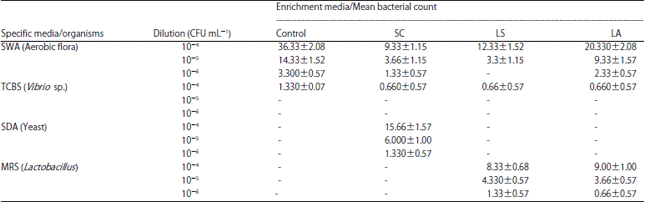 | |
| Each value is the Mean±SD of triplicate analysis | |
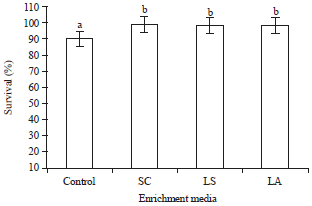 | |
| Fig. 1: | Survival (%) of A. franciscana nauplii after 24 h enrichment with control (Nanochloropsis), SC, LS and LA media at 104 concentration. Each value is the Mean±SD of triplicate analysis. Bars showing same alphabets are statistically non-significant (one-way ANOVA, p>0.05 and subsequent post hoc multiple comparison with SNK test), SC: S. cerevisiae, LS: L. sporogenes and LA: L. acidophilus |
The same trend was obtained in 10–5 and 10–6 dilution except no colonies was observed at Artemia enriched with L. sporogenes medium at 10–6 dilution (Table 2).
The mean Vibrio sp., load in the A. franciscana nauplii enriched with different probiotic sources was uniform with 0.66±0.57×10–4 CFU mL–1, whereas it was 1.33±0.07×10–4 CFU mL–1 in the nauplii enriched with control medium (Nanochloropsis). In the other concentrations (10–5 and 10–6), no Vibrio colonies identified in all the enrichment media. Yeast colonies were identified only in the Artemia nauplii enriched with yeast S. cerevisiae medium with the maximum of 15.66±1.57×10–4 CFU mL–1. Similarly maximum number of Lactobacillus colonies were only identified in the A. franciscana nauplii enriched with L. sporogenes (8.33±0.68×10–4 CFU mL–1) and L. acidophilus (9.0±1.00×10–4 CFU mL–1).
Pathogenic challenge studies in enriched Artemia nauplii: The A. franciscana nauplii were succumbed to death from 24 h onwards. Control nauplii had 28±2.0 and 32±1.0% mortality when challenged with the pathogens V. parahaemolyticus and V. cholerae, respectively. At the same time, the mortality percentage of yeast enriched Artemia nauplii was reduced to 20±1.0% and 26.0±1.0% in V. parahaemolyticus and V. cholerae treatment, respectively. In the case of L. sporogenes enriched Artemia nauplii, the reduction in mortality percentage was little higher when compared with control group i.e., 43% (16±2.0%) and 44% (18±3.0%) in respect of V. parahaemolyticus and V. cholerae treatment. Lactobacillus acidophilus showed better result on the reduction in mortality percentage of 57% (12±2.0%) and 50% (16±1.0%) in Artemia nauplii challenged against V. parahaemolyticus and V. cholerae, respectively (Table 3).
The mortality percentage of all the groups of Artemia nauplii challenged with V. parahaemolyticus and V. cholerae was constantly increasing at every 24 h interval. Finally, at 120th h, 100% mortality was occurred in all the tested groups except those nauplii enriched with L. acidophilus challenged with V. parahaemolyticus. In this group, 100% mortality was occurred only during 144th h.
The data observed for increase in the mortality percentage of Artemia nauplii with advancement of time were positively regressed (p<0.05) in all the tested groups challenged with both pathogens.
| Table 3: | Time and percentage mortality of Artemia franciscana nauplii enriched with control, SC, LS and LA after challenged with pathogen Vibrio sp. |
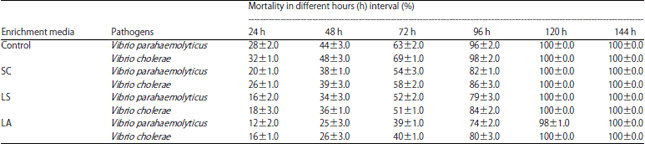 | |
| Each value is the Mean±SD of triplicate analysis | |
| Table 4: | Cumulative Mortality Index (CMI%) value of Artemia franciscana enriched with Control, SC, LS and LA exposed to pathogens V. parahaemolyticus and V. cholerae |
 | |
| Each value is the Mean±SD of triplicate analysis. Within each column, means with different superscript letters are statistically significant (One-way ANOVA, p<0.05 and subsequent post hoc multiple comparison with SNK test) | |
It was observed that there was a significant correlation between percentage mortality and time in all the tested groups (for V. parahaemolyticus: R2 = 0.9379-0.9747, for V. cholerae: R2 = 0.9220-0.9692).
Cumulative Mortality Index (CMI) calculated for A. franciscana nauplii in all the groups are provided in Table 4. The CMI for the control group of Artemia nauplii challenged with V. parahaemolyticus was 42936±528, which was reduced to 5.75, 7.43 and 12.52% in other groups, such as Artemia nauplii enriched with S. cerevisiae, L. sporogenes and L. acidophilus, respectively. Comparison of reduction in mortality percentage of control group with experimental groups was significantly varied (F = 4.832-23.717, p<0.01 to p<0.0001).
The CMI for the control group of A. franciscana nauplii challenged with V. cholerae was 43848, which was reduced to 5.74, 8.10 and 11.98% in Artemia nauplii enriched with S. cerevisiae, L. sporogenes and L. acidophilus, respectively. Comparison of reduction in mortality percentage of control group with experimental groups revealed that the variation was statistically more significant (F = 7.2287-10.9175, p<0.001).
DISCUSSION
The practice of using commercial antibiotics in the culture system of several fin fishes and shell fishes has led to the resistance development towards the bacterial strains. In hatcheries, because of this practices, several antibiotics, which were previously effective against certain pathogenic bacteria will become less or non-effective to the bacteria in due course. This may sometime requires high doses of antibiotics24. An alternative source for this is using antimicrobial characteristic probiotics instead of synthetic antibiotic drugs.
The beneficial effect of L. acidophilus, L. sporogenes and S. cerevisiae in human and farm animals has been reported in several studies25,26. Further more, these probionts have been preventing attachment of harmful bacteria and preventing infection27. The probionts , such as S. cerevisiae, L. acidophilus, L. sporogenes can be easily isolated from different types of food source. In the present study also these three probionts were isolated from food products such as milk, cheese and grapes.
The metabolites produced by these organisms are mainly acting as antibiotic26. In the present study, the effect of extracellular metabolites produced by the above probionts were tested against shrimp pathogens Vibrio species (V. cholerae and V. parahaemolyticus). Among these probionts, L. acidophilus and L. sporogenes are more active against V. parahaemolyticus and V. cholerae, whereas the inhibition level was comparatively low in the metabolite produced by yeast species S. cerevisiae. Hamilton-Miller et al.28 studied the preparation mode of probiotic supplemented diet with 13 different probionts. They concluded that only single strain supplemented diet could give better results rather multi strains of probionts.
Artemia enrichment or bioencapsulation is widely applied in marine fish and crustacean hatcheries all over the world for enhancing the nutritional value of Artemia with essential fatty acids and other nutrients29 and also for drug delivery30. Artemia nauplii and rotifers are also being used for the delivery of probionts to fish larvae and shrimp larvae10. In the present study, II instar stage of Artemia franciscana nauplii were enriched with algae (Nanochloropsis) as a control and probionts (Lactobacillus bacteria and yeast) as experimental in three different concentrations (102, 103 and 104 CFU mL–1) for 24 h. Then the survival rate was assessed and it showed that the Artemia nauplii enriched with control diet exhibited only 89% survival, whereas the probionts enriched Artemia nauplii exhibited 94-99% survival in all the groups. Earlier studies by Patra and Mohamed21 have reported that Artemia enriched with yeast (S. boulardii) alone showed a survival of 85% in all treatments.
Bacterial colonization of the nauplii could occur externally, via attachment to the body surfaces or internally by ingestion13. After the nauplii were removed from the bacterial suspension, the bacterial content decreased rapidly. This decrease might be due to the removal of the external bacteria after the nauplii were washed and placed in sterile water. The bacteria still detected could be the one colonizing the interior or firmly attached to the external surfaces31. In the present study, after 24 h of enrichment, the control and experimental Artemia nauplii were tested for the presence of aerobic flora Vibrios, Lactobacilli sp. and Yeast. For that only 103 CFU mL–1 enrichment was taken. In A. franciscana nauplii, the mean aerobic flora counted on SWA plates of control was maximum of 36.33±2.08×10–4 CFU mL–1. Whereas, it was comparatively 75% less in S. cerevisiae medium enriched Artemia nauplii. At the same time, 66 and 44% reduction of this load was observed in L. sporogenes and L. acidophilus media, respectively.
High degree of bacterial variability has been reported earlier in crustacean digestive tracts32. In an experiment, where turbot larvae were fed with Lactic Acid Bacteria (LAB) enriched rotifers, counts of 104 to 105 CFU per larva were retrieved in the groups fed with the increasing levels of LAB10. But no LAB colonies were detected for control group without LAB. The results obtained in the present study also indicated that the probionts (SC, LS and LA) did not significantly change the comparison of Vibrio and other aerobic flora.
Rico-Mora and Voltolina33 challenged Artemia nauplii with V. alginolyticus and V. parahaemolyticus isolates and they obtained almost 100% mortality after 24 h for the 1st species and 48 h for the second species. In penaeid shrimp, differences in the pathogenicity of bacteria could depend on the species tested34 and on the strain characteristics35. In the present study, Artemia nauplii enriched with, respective probiotic and control media, after challenged with Vibrio species showed 28-32% mortality in the control group after 24 h. At the same time, the mortality percentage of yeast enriched Artemia was reduced from 29-19%, followed by L. sporogenes (46-44%) and L. acidophilus (57-50%). Similarly, the survival rate of turbot larvae fed with rotifers enriched with LAB and then challenged with Vibrio pathogen for 72 h was higher than treatment without LAB10.
CONCLUSION
Based on the results obtained in the present study it can be concluded that probionts, such as S. cerevisiae, L. sporogenes and L. acidophilus are necessary to overcome a Vibrio infection in Artemia nauplii.
ACKNOWLEDGMENT
The author wishes to thank University Grants Commission (UGC), Newdelhi for its financial support in the form of a Research grant (F.No: 43-81/2014 (SR).
REFERENCES
- Witte, W., I. Klare and G. Werner, 1999. Selective pressure by antibiotics as feed additives. Infection, 27: S35-S38.
CrossRefDirect Link - Garriques, D. and G. Arevalo, 1995. An Evaluation of the Production and Use of a Live Bacterial Isolate to Manipulate the Microbial Flora in the Commercial Production of Penaeus vannamei Postlarvae in Ecuador. In: Swimming Through Troubled Water, Browdy, C.L. and J.S. Hopkins (Eds.). World Aquaculture Society, USA., pp: 53-59.
- Fuller, R., 1989. Probiotics in man and animals. J. Applied Bacteriol., 66: 365-378.
CrossRefPubMedDirect Link - Juven, B.J., F. Schved and P. Lindner, 1992. Antagonistic compounds produced by a chicken intestinal strain of Lactobacillus acidophilus. J. Food Protect., 55: 157-161.
Direct Link - Gatesoupe, F.J., 1991. Managing the dietary value of Artemia for larval turbot, Scophthalmus maximus; the effect of enrichment and distribution techniques. Aquacult. Eng., 10: 111-119.
CrossRefDirect Link - Gatesoupe, F.J., 1993. Bacillus sp. Spores as Food Additive for the Rotifer Brachionus plicatilis: Improvement of their Bacterial Environment and their Dietary Value for Larval Turbot, Scopthalamus maximus L. In: Fish Nutrition in Practice, Kaushik, S. (Ed.). Institut National de la Recherche Agronomique, Paris, pp: 561-568.
- Gatesoupe, F.J., 1994. Lactic acid bacteria increase the resistance of turbot larvae, Scophthalmus maximus, against pathogenic Vibrio sp. Aquat. Living Resour., 7: 277-282.
CrossRefDirect Link - Chair, M., M. Dehasque, S. van Poucke, H. Nelis, P. Sorgeloos and A.P. de Leenheer, 1994. An oral challenge for turbot larvae with Vibrio anguillarum. Aquacult. Int., 2: 270-272.
CrossRefDirect Link - Grisez, L., M. Chair, P. Sorgeloos and F. Ollevier, 1996. Mode of infection and spread of Vibrio anguillarum in turbot Scophthalmus maximus larvae after oral challenge through live feed. Dis. Aquat. Organisms, 26: 181-187.
CrossRefDirect Link - Chair, M., R.S.J. Gapasin, M. Dehasque and P. Sorgeloos, 1994. Vaccination of European sea bass fry through bioencapsulation of Artemia nauplii. Aquacult. Int., 2: 254-261.
Direct Link - Joosten, P.H.M., M. Aviles-Trigueros, P. Sorgeloos and J.H.W.M. Rombout, 1995. Oral vaccination of juvenile carp (Cyprinus carpio) and gilthead seabream (Sparus aurata) with bioencapsulated Vibrio anguillarum bacterin. Fish Shellfish Immunol., 5: 289-299.
CrossRefDirect Link - Campbell, R., A. Adams, M.F. Tatner, M. Chair and P. Sorgeloos, 1993. Uptake of Vibrio anguillarum vaccine by Artemia salina as a potential oral delivery system to fish fry. Fish Shellfish Immunol., 3: 451-459.
CrossRefDirect Link - Gibson, L.F., J. Woodworth and A.M. George, 1998. Probiotic activity of Aeromonas media on the Pacific oyster, Crassostrea gigas, when challenged with Vibrio tubiashii. Aquaculture, 169: 111-120.
CrossRefDirect Link - Rengpipat, S., W. Phianphak, S. Piyatiratitivorakul and P. Menasveta, 1998. Effects of a probiotic bacterium on black tiger shrimp Penaeus monodon survival and growth. Aquaculture, 167: 301-313.
CrossRefDirect Link - De Man, J.C., M. Rogosa and M.E. Sharpe, 1960. A medium for the cultivation of Lactobacilli. J. Applied Bacteriol., 23: 130-135.
CrossRefDirect Link - Patra, S.K. and K.S. Mohamed, 2003. Enrichment of Artemia nauplii with the probiotic yeast Saccharomyces boulardii and its resistance against a pathogenic Vibrio. Aquacult. Int., 11: 505-514.
CrossRefDirect Link - Immanuel, G., T. Citarasu, V. Sivaram, M.M. Babu and A. Palavesam, 2007. Delivery of HUFA, probionts and biomedicine through bioencapsulated Artemia as a means to enhance the growth and survival and reduce the pathogenesity in shrimp Penaeus monodon postlarvae. Aquacult. Int., 15: 137-152.
CrossRefDirect Link - Sivagnanavelmurugan, M., G.K. Ramnath, B.J. Thaddaeus, A. Palavesam and G. Immanuel, 2015. Effect of Sargassum wightii fucoidan on growth and disease resistance to Vibrio parahaemolyticus in Penaeus monodon post-larvae. Aquacult. Nutr., 21: 960-969.
CrossRefDirect Link - Buts, J.P., G. Corthier and M. Delmee, 1993. Saccharomyces boulardii for Clostridium difficile-associated enteropathies in infants. J. Pediatr. Gastroenterol. Nutr., 16: 419-425.
PubMedDirect Link - Kleeman, E.G. and T.R. Klaenhammer, 1982. Adherence of Lactobacillus species to human fetal intestinal cells. J. Dairy Sci., 65: 2063-2069.
CrossRefDirect Link - Hamilton-Miller, J.M., S. Shah and C.T. Smith, 1996. Probiotic remedies are not what they seem. Br. Med. J., 312: 55-56.
PubMedDirect Link - Aguilar-Aguila, A., A.T. Mansir and A.R. Manriquez, 1994. Using brine shrimp as a drug carrier for therapeutic applications in aquaculture. Aquacult. Eng., 13: 301-309.
CrossRefDirect Link - Gomez-Gil, B., M.A. Herrera-Vega, F.A. Abreu-Grobois and A. Roque, 1998. Bioencapsulation of two different Vibrio species in nauplii of the brine shrimp (Artemia franciscana). Applied Environ. Microbiol., 64: 2318-2322.
Direct Link - Dempsey, A.C., C.L. Kitting and R.A. Rosson, 1989. Bacterial variability among individual penaeid shrimp digestive tracts. Crustaceana, 56: 267-278.
CrossRefDirect Link - Rico-Mora, R. and D. Voltolina, 1995. Effects of bacterial isolates from Skeletonema costatum cultures on the survival of Artemia franciscana nauplii. J. Invertebr. Pathol., 66: 203-204.
CrossRefDirect Link - Vera, P., J.I. Navas and M.C. Quintero, 1992. Experimental study of the virulence of three species of Vibrio bacteria in Penaeus japonicus (Bate 1881) juveniles. Aquaculture, 107: 119-123.
CrossRefDirect Link - De la Pena, L.D., T. Tamaki, K. Momoyama, T. Nakai and K. Muroga, 1993. Characteristics of the causative bacterium of vibriosis in the kuruma prawn, Penaeus japonicus. Aquaculture, 115: 1-12.
CrossRefDirect Link