
N.M. Azuadi
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, Serdang Selangor, Malaysia
S.S. Siraj
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, Serdang Selangor, Malaysia
S.K. Daud
Department of Biology, Faculty of Science, Universiti Putra Malaysia, Serdang Selangor, Malaysia
A. Christianus
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, Serdang Selangor, Malaysia
S.A. Harmin
Centre of Land and Aquatic Biotechnology, Faculty of Science and Biotechnology, Universiti Industri Selangor, 45600 Bestari Jaya, Selangor, Malaysia
S. Sungan
Department of Agriculture, Indigenous Fisheries Research and Production Centre, Tarat, Sarawak, Malaysia
R. Britin
Department of Agriculture, Indigenous Fisheries Research and Production Centre, Tarat, Sarawak, Malaysia
Journal of Fisheries and Aquatic Science
Year: 2011 | Volume: 6 | Issue: 7 | Page No.: 740-750







ABSTRACT
Natural spawning ground of most Malaysian streams are undergoing extreme changes as a result of human activities, pollution and climate change (extreme temperature fluctuation) which lead to dramatic decrease in mahseer (Tor tambroides, Cyprinidae) populations. An experiment was conducted to evaluate the effectiveness of salmon gonadotropin releasing hormone analogue (sGnRHa) alone or in combination with dopamine antagonist domperidone (DOM) on breeding performances of female breeders. Sixty females T. tambroides ranging in weight 2.01 to 3.96 kg while thirty males with average weight 1.38 to 1.72 kg were used in this study. The females were given pretreatment with Ovaplant (23.4-44.9 μg kg-1) for 6 weeks prior to induction. Females were tested with various treatments such as Ovatide, sGnRHa, sGnRHa+DOM and DOM. SGnRH analogue alone does not contain dopamine antagonist to block dopaminergic inhibition action. Females were given single intramuscular injection of Ovatide (0.5 mL kg-1 BW), sGnRHa (10 μg kg-1 BW), sGnRHa+DOM (10 μg kg-1 BW+5 mg kg-1 BW) or DOM (5 mg kg-1 BW). The control group was administered with 0.9% NaCl. The fish did not spawn spontaneously but through hand stripping to release eggs. Results showed that Ovatide significantly produced highest ovulation rate, total stripped eggs and stripped fecundity. Interestingly, we observed that broodstock groups injected with sGnRHa+DOM produced highest fertilization, hatching and survival rates compared to other treatments. Good egg and larval quality were recorded in the group treated with sGnRHa+DOM. No females ovulated in the group receiving DOM alone and saline solution. The combination treatment was found to be more potent in inducing ovulation as compared to sGnRHa alone or DOM alone. Our results suggest that dopaminergic inhibition is a major barrier in inducing spontaneous spawning in captive mahseer and we successful produced progeny generation of T. tambroides larvae.
PDF Abstract XML References Citation
How to cite this article
N.M. Azuadi, S.S. Siraj, S.K. Daud, A. Christianus, S.A. Harmin, S. Sungan and R. Britin, 2011. Enhancing Ovulation of Malaysian Mahseer (Tor tambroides) in Captivity by Removal of Dopaminergic Inhibition. Journal of Fisheries and Aquatic Science, 6: 740-750.
DOI: 10.3923/jfas.2011.740.750
URL: https://scialert.net/abstract/?doi=jfas.2011.740.750
DOI: 10.3923/jfas.2011.740.750
URL: https://scialert.net/abstract/?doi=jfas.2011.740.750
INTRODUCTION
Tor tambroides (Bleeker, 1854) is a cyprinid commonly known as mahseer and locally referred to as kelah, empurau or pelian. It inhabits rivers and lakes, ascending to the rapid river as well rocky bottom to breed. Some of the Tor species such as T. khudree are described by Ogale (2002) as a sport fish that provides unbeatable recreation to fishing lovers around the world, better than salmon. In recent years due to human interference, kelah stocks have been threatened with multi-sided dangers such as overfishing, industrial pollution, deforestration which distrupt natural spawning ground of riverine fish populations. Deterioration of environmental conditions and demand of this species as food and game fish resulting in over exploitation of natural stocks lead to noticeable decline of mahseers in the wild (Ogale, 2002).
During the last 20 years, one of the major developments in fish culture is captive breeding. There has been a rising application of modern and efficient techniques to induce final oocyte maturation, ovulation, spawning and spermiation. These methods engage the use of synthetic agonists of gonadotropin-releasing hormone. Gonadotropin-releasing hormone (GnRH) treatment is gaining reception throughout the world, surpass the hypophysation techniques such as pituitary treatment that has been used for decennary (Yaron, 1995) in induced spawning of fish. Earlier studies on induced ovulation and spawning in Clarias species by administration of Luteinizing Hormone Releasing Hormone (LHRH) analogue alone or together with a dopamine antagonist (Ngamvongchon et al., 1988; Manickam and Joy, 1989; Tan-Fermin and Emata, 1993), sGnRH analogue(s) exhibited maximum activity when evaluating the structure-activity relationship of GnRH peptides.
Inducing final oocyte maturation and ovulation by using a type of GnRH analogues proved to be effective; however, fish biological potencies differ greatly. Salmon GnRH (sGnRH) analogue and mammalian GnRH analogue (mGnRH) or LHRHa are the most common GnRH analogues used in fish spawning induction. SGnRH was found to be more effective than mGnRH in rousing serum gonadotropin (GtH) levels and ovulation in goldfish Carassius auratus (Peter et al., 1985) common carp Cyprinus carpio and Chinese loach Paramisgurnus dabryanus (Lin et al., 1988, 1991) and African catfish Clarias gariepinus (Szabo et al., 2007). Dual control (positive and negative) are demostrated in many teleost fish. Gonadotropin (GtH) discharge is stimulated by gonadotropin releasing hormones (GnRH) and is repressed by dopamine (DA) (Chang and Peter, 1983; De Leeuw et al., 1985). The evidence concerning the dopaminergic inhibition of GtH release frequently comes from study reported in goldfish Carassius auratus and other cyprinid species (Peter et al., 1986) and in African catfish Clarias gariepinus (Van Oordt and Goos, 1987). In goldfish, dopamine can eradicate the pituitary's reaction to Luteinizing Hormone-releasing Hormone Analogue (LHRHa) or to lesion of dopaminergic tracts in the pituitary gland and preoptic nucleus (Chang and Peter, 1983). In the other hand, dopamine antagonists such as pimozide (PIM) and domperidone (DOM) can boost plasma GtH levels or, if administered in conjunction with LHRHa and sGnRHa, can markedly potentiate the response to LHRHa and sGnRHa.
Successful induced ovulation and multiple breeding in captivity by artificial propagation have been achieved in Tor tambroides (Ingram et al., 2005). Ovaprim that consist sGnRHa and domperidone is the most successful hormone used to induce ovulation in mahseer (Ingram et al., 2005). Ovulation induction using 0.5 mL Ovaprim in one dose proved to have highest fertilization rate compared to frog pituitary extract (FPE) and Clarias pituitary extract (CPE) (Adebayo and Popoola, 2008). Nowadays, larval production cost always concerned by hatcheries. They are looking for ways to reduce induction work cost while getting higher larval production. Adebayo and Popoola (2008) reported that production cost in African catfish was highest using CPE followed by Ovaprim and FPE. Popularity of newly formulated hormone such as Ovatide arises because of the affordable price and effectiveness as inducing agents compare to other hormones. The active ingredients in Ovatide and Ovaprim are almost the same but Ovatide has low cost and low viscosity compared to Ovaprim. The effectiveness of Ovatide is already tested in spotted snakehead Channa punctatus (Marimuthu et al., 2009), walking catfish Clarias batrachus (Sahoo et al., 2005) and Indian major carp Catla catla. Higher spawning fecundity and fertilization rate were reported in Indian major carps Cirrhina mrigala and Labeo rohita when spawning was induced with Ovatide compared to Ovaprim.
Domperidone is commonly use in combination with other hormones to induce spawning in many species such as nase Chondrostoma nasus, bighead carp, grey mullet, silver perch, African catfish, rohu, common carp and many other teleost fish. In some fishes there is a strong inhibition of basal and GnRH stimulated discharge of LH by Dopamine (DA) (Van der Kraak et al., 1988). Consequently administration of DA antagonists (e.g: Domperidone (DOM), reserpine, pimozide or metoclopramide) prior to the treatment with GnRHa (Ovaplant) removes the inhibition on gonadotrophs and enhanced the stimulatory effect for GnRHa on LH release. Domperidone has equal or somewhat greater potency than Pimozide in potentiating the action of releasing hormone agonist in loach; however, DOM had markedly greater efficiency in common carp (Lin et al., 1988).
Nutrition and food for breeders is a major concern in breeding activity. High protein requirement is important for development of eggs and production of good egg quality. Homemade food main ingredients using natural source such as soy bean, angkabang fruit, beans, ficus and palm oil fruit as main ingredients. In this study high protein fruit (angkabang) was used to feed broodstocks before induction work. High amounts of protein, carbohydrate and micro and macro elements in the breeder diet showed improvement in reproductive performance compare to other treatment diets (Dada and Ogunduyile, 2011).
This study aimed to compare effectiveness of dopamine antagonist on breeding performance in filial 1 generation of Tor tambroides females in captive condition.
MATERIALS AND METHODS
Experimental fish: F1 generation of broodfish (Tor tambroides) used in this study was obtained from Tarat Indigenous Fisheries Production and Research Center (IFPRC) farm, Serian, Sarawak. These fish were reared for 4-5 years in concrete recirculating aquaculture system (RAS) tanks at the farm facility of Lu Thian Tack (LTT) Aquaculture Farm, Asajaya, Sarawak. These fishes were fed at 5% body weight twice daily. In the morning, the fish were fed with self made feed (angkabang fruit, emperang fish, squid oil, vitamin C, flour) containing 62.6% crude protein, 17.6% crude lipid, 1.8% crude fibre, 12.7% ash, 5.3% carbohydrate and 4770.5 kal g-1 energy. In the evening, the fish were fed with commercial tilapia pellet (Cargill, Malaysia) at different protein contents according to their age.
Broodstock selection: Matured F1 broodfish comprised of 60 females and 30 males were selected from 124 m3 concrete tanks and acclimatized in separate 20 m3 concrete RAS tanks and maintained at ambient temperature of 26.0-27.3°C. The females were selected on the basis of having round and soft abdomen with swollen, protruding and reddish vents. For identification purposes, all the breeders were implanted with Passive Implant Transponders (PIT Tags) (American Veterinary Identification Devices (AVID), Norco, CA, USA). The female and male breeders with weight ranging from 2.01-3.96 and 1.38-1.72 kg, respectively were selected for induced breeding.
Hormone preparation and artificial propagation: Ovaplant (Syndel Laboratories, Vancouver, Canada) (75 and 150 μg pellets, 26.04-50.00 μg kg-1) was implanted 6 weeks before ovulation induction trials. Salmon gonadotropin releasing hormone analogue (sGnRHa) and domperidone (Sigma, St. Louis, MO, USA) were used. The sGnRHa and DOM were dissolved in acidified 0.9% saline solution (Chang et al., 1984a,b) before used. The various treatments and dosages used were: Ovatide (0.5 mL kg-1), sGnRHa (10 μg kg-1), sGnRHa+DOM (10 μg kg-1+5 mg kg-1), DOM (0.5 mL kg-1) and 0.9% saline solution (0.5 mL kg-1). Ovatide (Hemmo Pharma, Mumbai, India) supplied in liquid form containing 20 μg of sGnRHa and 10 mg of domperidone in 1.0 mL. Ovatide was injected at 0.5 mL kg-1 BW as recommended by distributor and used as a positive control. Fish were anesthetized with MS222 before implantation or injection to avoid stress and for ease of handling. After these procedures the fish were hold in 0.2% acriflavine solution to treat injury and reduce infection. The length and weight of the fish were recorded before the hormone administration. Ovulation rate was evaluated in the morning 24 h post induction by gently stripping the female’s abdomen. The female breeders were anesthetized with MS222 (1:1000) and slight pressure was applied to the abdomen to determine whether eggs could be stripped from the fish. Eggs were stripped directly into a plastic jar. Volume of the eggs was recorded and 1 mL of the eggs was taken as subsample for fecundity estimation. Two matured males on the basis of having viscous and creamy milt were selected to fertilize eggs of a female. The same stripping procedure was applied on the male for milt. The milt was then stripped into the jar and mixed for 3-7 min to optimize fertilization capacity (dry method) as described by Joshi et al. (2002). Water was added slowly into the jar to initiate fertilization and these eggs were then washed with fresh water. The water hardened eggs were incubated in separated plastic mesh trays according to broodfish. A water level of 5 cm above the eggs with water discharge at 1-2 L min was maintained. The eggs were completely covered with dark plastic with small holes to avoid direct exposure to the light. Unfertilized eggs turned opaque and were removed twice a day (7:00 am.. and 6:00 pm.) to avoid any fungal infection.
Eggs and larval quality measurements: Small aliquots of 1 mL eggs (n = 3) were sampled and used to calculate total stripped eggs. As for egg quality evaluation, fertilization and hatching rates were estimated. Fertilization rate was determined by observing at least 50 eggs displaying 32-cell stage which was 24 h Post Fertilization (PF). Unfertilized eggs were removed to prevent fungal infection. At fourth days PF, the eggs started to hatch and all the eggs hatched at 6 days PF. Hatching rate was determined as time from fertilization until 50% of the eggs hatched. Eggs were sampled by placing 100-200 eggs on a petri disk then the numbers of hatched larvae were counted. During the larval rearing period, 40 larvae from each replicated tank per egg clutch were taken from the rearing tanks, anesthetized with MS222 (1:10,000) and standard length measurements will be recorded on day 1 to 7 Post Hatching (PH). Deformed larvae were recorded. These were done in triplicates and survival rate was calculated. Average of 40 eggs and larval were measured under stereomicroscope, SMZ-168 (2x magnification, 34.5 mm working distance) to determine egg diameter and larval length, respectively. Duration of embryogenesis was determined as the time between fertilization and hatching rate.
Water quality parameters: Water quality parameters such as water temperature, dissolved oxygen, pH, conductivity, turbidity, salt and total dissolve solid were monitored daily using multiparameter water quality apparatus, WQC 24-1-2.
Statistical analysis: Data were expressed as Mean±Standard error of the mean (SEM). Differences in the mean between treatments were tested using one-way analysis of variance (ANOVA), p<0.05, followed by Duncan multiple comparison test. Differences in ovulation success were tested using Chi-Square test for independence. These tests were carried out using SPSS 16.0 software.
RESULTS
Breeding performances: Daily water temperature, dissolved oxygen, pH, conductivity, turbidity, salt and total dissolved solid (26.1-27.4°C, 5.95-13.4 mg L-1, 7.43-8.0, 8.6-13.9% FS, 0% FS, 0% and 0-0.1 g L-1, respectively) were recorded. This condition is suitable for ovulation induction activity. Ovulation in female broostocks was detected 24 h post administration. The ovulatory performance, egg quality and larval production of T. tambroides at different treatments were summarized in Table 1. Ovatide treatment showed the most significant (p<0.05) ovulation rate (50% ovulated) among all the treatments. The numbers of ovulated fish in sGnRHa alone (17% ovulated) and sGnRHa+DOM (25% ovulated) were not different from each other but significantly higher than DOM alone and saline treatment. Unovulated fish were observed in the DOM and saline treatments. The total stripped eggs and fecundity was significantly highest (p<0.05) when females were injected with Ovatide. Injection with sGnRHa+DOM (10 μg kg-1+5 mg kg-1 BW) resulted in lowest stripped egg yield. Significantly highest (p<0.05) fertilization, hatching and survival rates were obtained in fish administered with sGnRHa+DOM compared to those with Ovatide and sGnRHa. Deterioration of egg quality was observed at sGnRHa treatment leading to low fertilization and hatching percentages. Normal larval production rate was significantly highest in the group treated with sGnRHa+DOM but the highest total larval production was highest from Ovatide treatment. Significantly highest (p<0.05) abnormally was observed the group treated with sGnRHa alone than Ovatide and sGnRHa+DOM treatment. Types of abnormalities are shown in Fig. 2.
Egg and larvae quality: The egg diameter ranged from 2.92 to 3.3, 2.74 to 3.12 and 2.92 to 3.43 mm in Ovatide, sGnRHa and sGnRHa+DOM treatment, respectively. The mean egg diameter was significantly higher (p<0.05) in group treated with sGnRHa+DOM than Ovatide and sGnRHa egg.
| Table 1: | Effectiveness of dopamine antagonist (DOM) on ovulation, total stripped eggs, fecundity, fertilization, hatching, survival and larval production of T. tambroides. The observation were based on n=3 of twelve replicates |
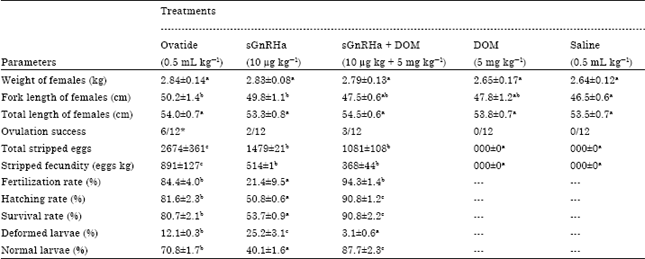 | |
| Means value with the different superscript was significantly different (p<0.05) evaluated using one-way ANOVA followed by Duncan’s post-hoc test, *Significantly greater than the other treatments (p<0.05, χ2test) | |
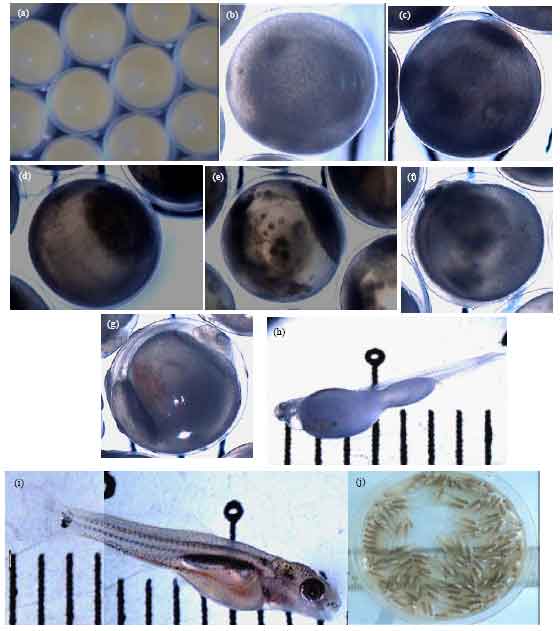 | |
| Fig. 1(a-j): | Embryonic and larval development in Tor tambroides, (a): Water hardened eggs, (b): Fertilized egg, (c): One cell stage, (d): 4 h PF egg (16 cell stage), (e): 9 h PF egg (morula stage) and (f). 32 h PF egg (eyed stage). (g): 70 h PF egg (ready to hatch). (h): Just hatch larvae. I: Seven day old larvae. J: Fourteen days old fry |
A similar trend was observed in diameter of the water hardened eggs. Duration for embryogenesis process significantly faster (p<0.05) in the group treated with sGnRHa+DOM (80 h 0 min±8 min) compare to the group treated with Ovatide and sGnRHa alone (81 24±7 and 83 h 24 min±26 min, respectively) Table 2. Development of egg from PF until 7 days old larva is shown in Fig. 1a-f. Eggs in the sGnRHa treated group were the last to hatch and produced higher number of deformed larvae. Larval length for the 1 day Post Hatch (PH) was significantly longest in the group treated with sGnRHa+DOM (8.6 to 9.5 mm).
| Table 2: | Eggs and larval qualities of Malaysian mahseer (Tor tambroides) injected with positive control of Ovatide or sGnRHa alone or DOM alone or combined sGnRHa + DOM or negative control of 0.9% saline solution |
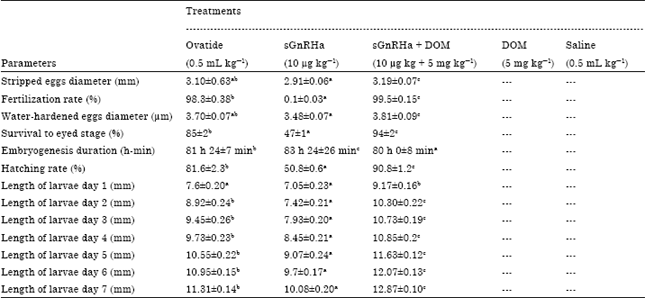 | |
| Means values with the different superscript were significantly different (p<0.05) evaluated using one-way ANOVA followed by Duncan’s post-hoc test | |
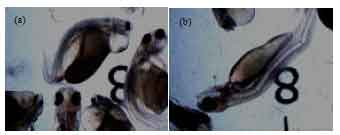 | |
| Fig. 2(a-b): | Deformed larvae collected from the group treated with sGnRHa alone |
Nonsignificant larval length was observed in group treated with Ovatide (7-8.3 mm) and sGnRHa alone (6.4-7.5 mm). At 2 day PH, larval length increased and showed significantly longest (p<0.05) in the group treated sGnRHa+DOM (9.8-11 mm) followed by Ovatide (8-9.5 mm) and sGnRHa alone (6.8-7.8 mm). This pattern was similar until day 7 PH. At 7 day PH, the increment of larval length continued and showed significantly longest in the group treated with sGnRHa + DOM (12.5-13.2 mm) followed by Ovatide (11-12 mm) and sGnRHa (9.5-10.8 mm). Increment of the larvae length is shown in the Fig. 1.
DISCUSSION
In this experiment, viable hatched larvae were produced from female broodstock weighing 2.78 kg. Ingram et al. (2007) reported that over 2.5 kg female broodfish were sexually matured and can produce hatchable eggs. While, in peninsular Malaysia, only female breeders of T. tambroides over 4.5 kg were found to be sexually matured (Tan, 1980). In Andang region of Sarawak, Sungan (1991) observed that females T. douronensis as small as 0.625 kg are able to release free-flowing eggs which are considered as matured. No females found ovulated in the control group probably because as observed in earlier finding that mahseer did not spawn in captivity naturally (Ingram et al., 2005). Similar result was noted by Taufek et al. (2009) when African catfish reared in captivity did not ovulate when induced with saline solution. The highest ovulation rate was observed in 0.5 mL kg-1 BW Ovatide treatment. Ovatide successful induced spawning in several fish species such as; stinging catfish, Heteropneustes fossilis (Marimuthu et al., 2000); spotted snakehead, Channa Punctatus (Marimuthu et al., 2009); walking catfish, Clarias batrachus (Sahoo et al., 2005) and several cyprinid species like silver carp, Hypophthalmichthys molitrix and common carp, Cyprinus carpio (Horvath et al., 1997).
DOM alone failed to induce ovulation in Malaysian mahseer females because it was not able trigger complete ovulation thus resulting in nonresponse to stripping. This might be due to insufficient release of gonadotropin which slows the eggs maturation process (Van der Kraak et al., 1983; Billard et al., 1984; Sahoo et al., 2005). DOM without sGnRHa cannot help in oocyte maturation and induce ovulation. Higher stripping pressure and egg plunging was observed when sGnRHa was administered resulting in blood tinged eggs. Blood on the stripped egg coagulated and clogged the micropile which lead to poor fertilization. Treatment with sGnRHa alone caused low egg production and eggs quality which affect larval production. Low egg quality may lead to higher larval deformities. Highest deformity in sGnRHa alone treatment may be attribute to the fertilization of unripe ova which leading to abnormal embryonic development. Combination of DOM and GnRHa accelerate oocyte development and increase plasma estradiol levels (Aizen et al., 2005). The highest production of larvae was obtained when treated with Ovatide but fertilization, hatching and larval survival rates were highest when treated with sGnRHa+DOM. This could be due to highest production of eggs in Ovatide treatment but lower fertilization, hatching and survival rates compare to sGnRHa+DOM treatment. Treatment with sGnRHa+DOM enhances fertilization, hatching and survival rates. In this group, eggs diameter and water hardened eggs were significantly higher compared to other groups. This situation lead to production of longer and healthy larvae compared to other treatments. Previous study showed that striped seabass injected with triiodothyronine (T3) prior to spawning, produced eggs and larvae, consequently increase in growth, swimbladder inflation and larval/fingerling survival.
Low effect of sGnRHa alone or DOM alone on oocyte maturation and ovulation probably because of inadequate and failure of GtH levels increment in circulation (Szabo et al., 2002). SGnRHa and DOM alone were not effective to trigger oocyte maturation and ovulation in females and need to combine. Combination of both also will stimulate adequate increase in GtH level in the circulation. Dopamine is released by the hypothalamus and acts as a GtH release inhibitor in many teleost and has been shown to directly inhibit basal and GnRH-stimulated GtH release (Peter et al., 1991). Combination of dopamine antagonist and sGnRHa was highly effective in increasing circulating GtH levels and inducing ovulation in goldfish and common carp (Szabo et al., 2002). Similar to other cyprinids, the results of this study showed that dopamine acts as an inhibitory factor on Gth releasing. DOM alone was unable to induce ovulation. Combination of sGnRHa and dopamine antagonist domperidone proves effective in inducing ovulation. In some fish, strong dopaminergic inhibition has been demonstrated, consequently dopamine receptor antagonists often combined with GnRHa (Szabo, 2003). Pimozide, domperidone and metoclopramide are the most commonly used dopamine antagonist which potentiating the effects of GnRHa (Yaron, 1995).
The present study demonstrated that Ovatide (0.5 mL kg-1) treatment was the most successful in inducing ovulation followed by sGnRHa+DOM (10 μg kg-1+5 mg kg-1) and sGnRHa (10 μg kg-1). Five mg kg-1 DOM alone and the control treatment showed no potential at all. Combination of sGnRHa with dopamine antagonist (DOM) showed more potent than sGnRHa alone or DOM alone in inducing ovulation. This study is very important in providing baseline information to help commercial hatcheries to produce optimum collection of good quality eggs leading to higher larval production.
ACKNOWLEDGMENTS
The authors thank Agri-Biotechnology Institute (ABI) for financial support and LTT Aquaculture Farm, Asajaya, Sarawak Malaysia in allowing use its facilities and the broodstocks. Thank also goes to Tarat Indigenous Fisheries Production and Research Center (IFPRC) farm, Serian, Sarawak for the full support and co-operation.
REFERENCES
- Adebayo, O.T. and O.M. Popoola, 2008. Comparative evaluation of efficacy and cost of synthetic and non-synthetic hormones for artificial breeding of African catfish (Clarias gariepinus Burchell, 1822). J. Fish. Aquat. Sci., 3: 66-71.
CrossRefDirect Link - Aizen, J., I. Meiri, I. Tzchori, L.S. Sivan and H. Rosenfeld, 2005. Enhancing spawning in the grey mullet (Mugil cephalus) by removal of dopaminergic inhibition. Gen. Comp. Endocrinol., 142: 212-221.
CrossRefDirect Link - Billard, R., K. Bieniarz, R.E. Peter, M. Sokolowka, C. Weil and L.W. Crim, 1984. Effects of LHRH and LHRH-A on plasma GtH levels and maturation/ovulation in the common carp, Cyprinus carpio, kept under various environmental conditions. Aquaculture, 41: 245-254.
CrossRef - Chang, J.P. and R.E. Peter, 1983. Effects of dopamine on gonadotropin release in female goldfish, Carassius auratus. Neuroendocrinology, 36: 351-357.
PubMedDirect Link - Chang, J.P., D.S. MacKenzie, D.R. Gould and R.E. Peter, 1984. Effects of dopamine and norepinephrine on in vitro spontaneous and gonadotropin-releasing hormone-induced gonadotropin release by dispersed cells or fragments of the goldfish pituitary. Life Sci., 35: 2027-2033.
CrossRefDirect Link - Chang, J.P., R.E. Peter and L.W. Crim, 1984. Effects of dopamine and apomorphine on gonadotropin release from the transplanted pars distalis in goldfish. Gen. Comp. Endocrinol., 55: 347-350.
CrossRefPubMedDirect Link - Dada, A.A. and F.D. Ogunduyile, 2011. Effects of velvet bean (Mucuna pruriens) on sperm quality of African catfish, Clarias gariepinus (Burchell, 1822) broodstock. J. Fish. Aquat. Sci., 6: 655-661.
CrossRefDirect Link - De Leeuw, R., J.W. Resink, E.J.M. Rooyakkers and H.J.T. Goos, 1985. Pimozide modulates the luteinizing-hormone-releasing hormone effect on gonadotropin release in the African catfish, Clarias lazera. Gen. Comp. Endocrinol., 58: 120-127.
CrossRefDirect Link - Ingram, B., S. Sungan, G. Gooley, S.Y. Sim, D. Tinggi and S.S. De Silva, 2005. Induced spawning, larval development and rearing of two indigenous Malaysian mahseer, Tor tambroides and T. douronensis. Aquacult. Res., 36: 983-995.
CrossRefDirect Link - Ingram, B.A., S. Sungan, D. Tinggi, Y.S. Sim and S.S. De Silva, 2007. Breeding performance of Malaysian mahseer, Tor tambroides and T. douronensis broodfish in captivity. Aquacult. Res., 38: 809-818.
CrossRefDirect Link - Joshi, P.L., T.B. Gurung, S.R. Basnyat and A.P. Nepal, 2002. Domestication of wild golden mahseer (Tor putitora) and hatchery operation. FAO Fish. Tech. Pap., 431: 173-178.
Direct Link - Lin, H.R., G. Van Der Kraak, X.J. Zhou, J.Y. Liang, R.E. Peter, J.E. Rivier and W.W. Vale, 1988. Effects of [D-Arg6, Trp7, Leu8, Pro9 NEt]-luteinizing hormone-releasing hormone (sGnRH-A) and [D-Ala6, Pro9NEt]-luteinizing hormone-releasing hormone (LHRH-A), in combination with pimozide or domperidone, on Gonadotropin release and ovulation in the Chinese loach and common carp. Gen. Comp. Endocrinol., 69: 31-40.
CrossRefDirect Link - Lin, H.R., X.J. Zhou, G. Van Der Kraak and R.E. Peter, 1991. Effects of gonadotropin-releasing hormone agonists and dopamine antagonists on gonadotropin secretion and ovulation in Chinese loach, Paramisgurnus dabryanus. Aquaculture, 95: 139-147.
CrossRefDirect Link - Manickam, P. and K.P. Joy, 1989. Induction of maturation and ovulation by pimozide-LHRH analogue treatment and resulting high quality egg production in the Asian catfish, Clarias batrachus (L.). Aquaculture, 83: 193-199.
CrossRefDirect Link - Marimuthu, K., M.A. Haniffa and M.A. Rahman, 2009. Spawning performance of native threatened spotted snakehead fish, Channa punctatus (Actinopterygii: Channidae: Perciformes), induced with ovatide. Acta Ichthyologica Piscatoria, 39: 1-5.
CrossRefDirect Link - Ngamvongchon, S., O. Pawaputanon, W. Leelapatra and W.E. Johnson, 1988. Effectiveness of an LHRH analogue for the induced spawning of carp and catfish in Northest Thailand. Aquaculture, 74: 35-40.
CrossRefDirect Link - Peter, R.E., J.P. Chang, C.S. Nahorniak, R.J. Omeljaniuk, M. Sokolowska, S.H. Shih and R. Billard, 1986. Interactions of catecholamines and GnRH in regulation of gonadotropin secretion in teleost fish. Recent Prog. Horm. Res., 42: 513-548.
PubMedDirect Link - Peter, R.E., C.S. Nahorniak, M. Sokolowska, J.P. Chang and J.E. Rivier et al., 1985. Structure-activity relationships of mammalian, chicken and salmon gonadotropin-releasing hormones in vivo in goldfish. Gen. Comp. Endocrinol., 58: 231-242.
CrossRefPubMedDirect Link - Sahoo, S.K., S.S. Giri and A.K. Sahu, 2005. Effect on breeding performance and egg quality of Clarias batrachus (Linn.) at various doses of ovatide during spawning induction. Asian Fish. Sci., 18: 77-83.
Direct Link - Szabo, T., C. Medgyasszay and L. Horvath, 2002. Ovulation induction in nase (Chondrostoma nasus, Cyprinidae) using pituitary extract or GnRH analogue combined with domperidone. Aquaculture, 203: 389-395.
CrossRefDirect Link - Szabo, T., 2003. Ovulation induction in northen pike Esox lucius L. using different GnRH analogues, ovaprim, dagin and carp pituitary. Aquacult. Res., 34: 479-486.
CrossRefDirect Link - Szabo, T., F. Radics, T. Barth and L. Horvath, 2007. In vivo activity of native GnRHs and their analogues on ovulation in the African catfish, Clarias gariepinus (Burchell). Aquacult. Res., 38: 140-146.
CrossRefDirect Link - Tan, E.S.P., 1980. Some aspects of the biology of Malaysian riverine cyprinids. Aquaculture, 20: 218-289.
CrossRefDirect Link - Tan-Fermin, J.D. and A.C. Emata, 1993. Induced spawning by LHRHa and pimozide in the Asian catfish Clarias macrocephalus (Gunther). J. Applied Ichthyol., 9: 89-96.
CrossRefDirect Link - Taufek, N., S.A. Harmin and A. Christianus, 2009. Effect of chicken gonadotropin-releasing hormone (cGnRH-II) on plasma steroid hormone, maturation and ovulation in African catfish, Clarias gariepinus (Burchell). Afr. J. Biotechnol., 8: 6700-6709.
Direct Link - Van der Kraak, G., H.R. Lin, E.M. Donaldson, H.M. Dye and G.A. Hunter, 1983. Effects of LH-RH and des-Gly10 [D-Ala6] LH-RH-ethylamide on plasma gonadotropin levels and oocyte maturation in adult female coho salmon (Oncorhynchus kisutch). Gen. Comp. Endocrinol., 49: 470-476.
PubMedDirect Link - Van Oordt, P.G.W.J. and H.J.T. Goos, 1987. The African catfish, Clarias gariepinus, a model for the study of reproductive endocrinology in teleosts. Aquaculture, 63: 15-26.
CrossRefDirect Link - Yaron, Z., 1995. Endocrine control of gametogenesis and spawning induction in the carp. Aquaculture, 129: 49-73.
CrossRef