
M. L. Pappas
Laboratory of Agricultural Entomology and Zoology, Department of Agricultural Development, Democritus University of Thrace, Pantazidou 193, 68 200, Orestiada, Greece
G. D. Broufas
Laboratory of Agricultural Entomology and Zoology, Department of Agricultural Development, Democritus University of Thrace, Pantazidou 193, 68 200, Orestiada, Greece
D. S. Koveos
Laboratory of Applied Zoology and Parasitology, Faculty of Agriculture, Aristotle University of Thessaloniki, 54 124, Thessaloniki, Greece
Journal of Entomology
Year: 2011 | Volume: 8 | Issue: 3 | Page No.: 301-326







ABSTRACT
The family Chrysopidae includes many species that could be considered important biological control agents. Chrysopid larvae and the adults of certain species are polyphagous predators and feed on several pests of economic importance. The use of chrysopids in biological control has been enabled by the development of efficient mass-rearing facilities for species bearing certain characteristics, such as Chrysoperla species. From the different approaches used in implementing biological control, augmentation release is the most commonly applied method in the case of chrysopids. To date, only some Chrysoperla sp. are commercially available for use in biological control. Food supplements, attractants and hibernation boxes are also available for enhancing conservation of chrysopids. Difficulties in the systematics of chrysopids have made the release of Chrysoperla sp. more problematic. The evaluation of the efficacy of chrysopid species that share common characteristics with Chrysoperla sp. as potential biological control agents, as well as further studying the performance of Chrysoperla species under field conditions could enhance their role in Integrated Pest management (IPM).
PDF Abstract XML References Citation
Received: August 09, 2010;
Accepted: August 21, 2010;
Published: November 08, 2010
How to cite this article
M. L. Pappas, G. D. Broufas and D. S. Koveos, 2011. Chrysopid Predators and their Role in Biological Control. Journal of Entomology, 8: 301-326.
DOI: 10.3923/je.2011.301.326
URL: https://scialert.net/abstract/?doi=je.2011.301.326
DOI: 10.3923/je.2011.301.326
URL: https://scialert.net/abstract/?doi=je.2011.301.326
INTRODUCTION
Several chrysopid species are included among the most important aphidophagous predators. Their efficacy in biological control of aphids, as well as other arthropod pests has been well recognized for more than 250 years (Senior and McEwen, 2001). Due to the larval polyphagous feeding habits, chrysopids are considered to be important natural enemies of several pests besides aphids, such as whiteflies, thrips, lepidopteran pests and mites (Principi and Canard, 1984). In the context of biological control, most attempts have evaluated the efficacy of Chrysoperla carnea sensu lato in augmentation releases in the field or in the greenhouses (Ridgway and McMuphy, 1984; Hagen et al., 1999; Nordlund et al., 2001).
An important issue in the successfull use of chrysopids in biological control is the problematic systematics that still persists in the identification of Chrysoperla species (Tauber et al., 2000). Failures in chrysopid releases may, at least to some extent be explained by importations of wrongly identified species (Senior and McEwen, 2001).
In cases when the use of commercialy available chrysopids such as Chrysoperla carnea (Stephens) or Chrysoperla rufilabris (Burmeister) is problematic, other species could be exploited.
Species of the genera Mallada Navás, Dichochrysa Yang and Ceraeochrysa Adams might be proven efficient candidates for use in biological control (López - Arroyo et al., 1999a, b; Tauber et al., 2000; Daane, 2001). Data on the predation efficacy of other chrysopid species besides Chrysoperla are limited and mainly referred to laboratory conditions.
Research on chrysopid species occurs worldwide and comprises recently an important section in applied entomological research. The International Association for Neuropterology (IAN, www.neuroptera.com) consists a scientific forum that provides the opportunity for communication between researchers interested in the study of Neuropteroidea in general. Many of the IAN members are actively involved in the study of the biology and use of chrysopids in biological control programs. Their work has been well documented and summarized in the Proceedings of International Symposia organized by the Association. Ten Symposia have been held since 1982 and the Proceedings of most of them have been published or are currently in press (Gepp et al., 1984, 1986; Mansell and Aspöck, 1990; Canard et al., 1992, 1996; Panelius, 1998; Sziraki, 2002; Pantaleoni et al., 2005 (2007).
Research on all aspects of the biology of chrysopids and their use in biological control has been comprehensively reviewed since 1984 in two books, the ‘Biology of Chrysopidae’ edited by Canard et al. (1984) and "Lacewings in the Crop Environment" by McEwen et al. (2001). In the present study, the integration of most important factors that could affect and shape the efficacy of chrysopid species in biological control, as well as considerations for further research have been reviewed.
FAMILY CHRYSOPIDAE: THE MOST IMPORTANT GENERA FOR USE IN BIOLOGICAL CONTROL
The family Chrysopidae contains about 1200 species that belong in 75 genera and 11 sub- genera. The larvae of all species, as well as the adults of certain species are predaceous and could be important biological control agents of several soft-bodied arthropods, such as aphids, coccids and mites. Due to their important role in biological control programs, the species of the family Chrysopidae are the most well studied among Neuroptera (Brooks and Barnard, 1990).
Chrysopid systematics have been problematic for a long time with a great number of species from different places of the world and different time periods to be wrongly referred to as species of the genus Chrysopa Leach (New, 2001). An important contribution to avert these conflicting and contradictory assumptions was the review of Brooks and Barnard (1990) that identified and presented 1200 species of the family with regard mainly to the morphology of male genitalia. These species have been categorized in three sub-families, namely Nothochrysinae, Apochrysinae and Chrysopinae based on adult morphology (Brooks and Barnard, 1990). The Chrysopinae includes almost 97% of well known chrysopid species and specifically all those species of great economic importance commonly attracted in houses by lights during night (Brooks and Barnard, 1990; De Freitas and Penny, 2001). The species that belong to the genera Dichochrysa, Chrysopa, Ceraeochrysa and Chrysoperla Steinmann are considered to be the most important and abundant biological control agents (Brooks and Barnard, 1990; Albuquerque et al., 2001; Daane, 2001).
Dichochrysa is to date the largest in species number genus of the family Chrysopidae with at least 130 described species that can be found worldwidely but not at the Neotropics (Brooks and Barnard, 1990). According to morphological, biological and phylogenetic studies, many more Dichochrysa species will be discovered in the future in other places of the world (Aspock et al., 1980; Brooks and Barnard, 1990; Dong et al., 2004). Many species formerly described in the genus Mallada by Brooks and Barnard (1990), are now considered to belong to Dichochrysa based on morphological features (Brooks, 1997), although differences in their biology remain to be defined (New, 2001). Furthermore, the genus Anisochrysa Nakahara is considered to be synonym to Mallada (Dong et al., 2004). According to Daane (2001), species that belong to Mallada genus will prove to be efficient biological control agents in the future. Dichochrysa species have been recorded in several field crops such as fruit and nut orchards, vegetables, ornamentals, as well as in forests (Greve, 1984; New, 1984; Semeria, 1984; Zeleny, 1984; Duelli, 2001; Szentkirályi, 2001a, b). Only the trash-carrier larvae are predaceous, whereas the adults feed mainly on pollen and honeydew (New, 2001).
Several species of the family Chrysopidae have been for a long time mistakenly considered to belong to the genus Chrysopa. According to Brooks and Barnard (1990), Chrysopa sensu stricto genus includes approximately 50 species and some sub - species. Both the adults and larvae are predaceous (New, 2001).
The genus Ceraeochrysa includes 46 species and is the dominant genus in the Neotropics (Brooks and Barnard, 1990; Tauber et al., 2000).Ceraeochrysa species are recorded from Canada to Argentina and mainly in the tropics (De Freitas and Penny, 2001). They are commonly found in orchards and several agroecosystems where they are important biological control agents (Lopez-Arroyo et al., 1999a, b; Tauber et al., 2000; Albuquerque et al., 2001).
The genus Chrysoperla includes approximately 36 species that can be found worldwidely (Brooks and Barnard, 1990). Their biology and ecology have been extensively studied and these are the species that are mainly used in biological control programs (Brooks, 1994; New, 2001). However, Chrysoperla systematics are currently problematic, especially for those species of the- carnea group (Brooks, 1994). The adults are not predaceous and feed on honeydew and pollen, whereas larvae are polyphagous predators (New, 2001).
GENERAL BIOLOGICAL ASPECTS OF CHRYSOPIDS
The biology of the most well studied chrysopid species has been reviewed by Canard and Principi (1984), Canard (2001) and Canard and Volkovich (2001).
Chrysopid eggs are oval, white, yellow or green, their size ranges from 0.7 to 2.3 mm and are usually laid on the edge of a silken stalk (with the exception of the species that belong to the genus Anomalochrysa McLachlan which lay unstalked eggs) (Gepp, 1984). The stalk is assumed to play an important defensive or nutritional role for the newly hatched larva and its length is a species specific characteristic that may vary from 2 to 26 mm and be influenced by the mother’s body size, as well as environmental conditions (Tauber et al., 2003). Recently, two silk genes expressed by Mallada signata (Schneider) females were identified and sequenced by Weisman et al. (2009).
Eggs are usually laid by the female singly or in clusters (Duelli, 1984; Gepp, 1984) on the underside or on the top of leaves or shoots where there is plenty of food facilitating its discovery by the young larvae (Duelli, 1984). Embryonic developmental time depends mainly on temperature. For example, C. carnea embryos develop in about 13 days at 15°C and 2.5 days at 35°C, whereas for Chrysoperla externa (Hagen) in 14 and 4 days, at 15.6 and 26.7°C, respectively (Canard and Principi, 1984; Albuquerque et al., 1994).
Chrysopid larvae (three larval instars) can be divided in two groups in respect to their morphology and behavior (Gepp, 1984; Diaz-Aranda et al., 2001; Tauber et al., 2003). In the first group, the debris-carrying larvae or trash-carriers (e.g., larvae of the genera Dichochrysa, Ceraeochrysa, Glenochrysa Esben-Petersen, Leucochrysa McLachlan, Italochrysa Principi), collect with their mouthparts several pieces of plant material, exuviae or dead prey items and place them on their dorsa, thus forming a small ‘packet’ of ‘trash’ that is supposed to protect them from ants and other natural enemies (Eisner et al., 1978). In the second group, the larvae do not carry trash on their dorsa and are naked (e.g., Chrysoperla larvae).
Larval development is mainly influenced by temperature (Canard and Principi, 1984). In most cases, developmental time is shorter in Chrysoperla sp. in relation to Dichochrysa sp. or Nothochrysa McLachlan sp. For example, the larval developmental time for C. externa ranged from 46.5 days at 15.6°C to 12.2 days at 26.7°C (Albuquerque et al., 1994), whereas for Dichochrysa prasina Burmeister ranged from 85 to 19.7 days at 15 and 30°C, respectively (Pappas et al., 2008a). All chrysopid larvae are polyphagous predators with a broad range of prey, such as aphids, coccids, cicadellids, whiteflies, thrips, psyllids, eggs and larvae of Lepidoptera, Coleoptera, Diptera or Neuroptera and eriophyid or tetranychid mites (Canard and Principi, 1984; Canard, 2001). Prey location is usually random but there are also cases when it has been mediated to a certain extent by the honeydew produced by the prey (Canard, 2001). There are also chrysopid species that are highly specific to a certain prey species, such as Chrysopa slossonae Banks larvae which are only associated with the wooly alder aphid, Priciphilus tesselatus (Fitsch) (Tauber and Tauber, 1987; Milbrath et al., 1993). Larval prey quantity and quality may have a great influence on preimaginal development, as well as on the reproductive potential of adults (Principi and Canard, 1984; Canard, 2001).
Upon the completion of larval development, the third instar fully grown larva spins a cocoon and remains inside it until adult emergence (Canard and Principi, 1984). Cocoons are usually placed on the plant, inside curled leaves, on the leaves or in the soil (Canard and Volkovich, 2001). The development of the cocoon usually lasts for one or two weeks and depends on several factors, mainly temperature and sex. For example, cocoon development in C. externa was completed in 7.1 days at 24°C, whereas for Chrysopa pallens (Rambur) in 12.7 days at 20°C (Grimal and Canard, 1990; Carvalho et al., 1998).
Most of chrysopid adults are not predaceous and feed on plant derived food, such as nectar and pollen, as well as with insect honeydew (e.g., aphid or coccid honeydew). Pollen grains have been found in relatively high quantities in the digestive trunk of C. carnea and D. prasina adults (Bozsik, 1992, 2000; Villenave et al., 2005). The adults of species of the genera Anomalochrysa, Atlantochrysa Hölzel and Chrysopa are considered to be predaceous (Brooks and Barnard, 1990). Besides several insects, such as aphids, coccids and mites, other food type like pollen grains, fungi spores and yeast have also been identified in the midgut of predaceous chrysopid adults (Principi and Canard, 1984; Bozsik, 1992; Canard, 2001).
To date, published research articles have mainly focused on the effects of several factors on certain biological aspects of chrysopids relative to their use in biological control. A great number of researchers have studied the effects of several abiotic factors, such as temperature (Honek and Kocourek, 1988; Tauber et al., 1987, 2006; Albuquerque et al., 1994; Volkovich and Arapov, 1996; López-Arroyo et al., 1999a; Nakahira et al., 2005; Mantoanelli et al., 2006; Silva et al., 2007; Pappas et al., 2008a, c), photoperiod (Tauber and Tauber, 1972; Hodek and Honek, 1976; Nechols et al., 1987; Canard, 1988, 1990, 1997, 2005; Grimal, 1988; Canard and Grimal, 1988; Chang et al., 1995; Volkovich and Arapov, 1996; Volkovich and Blumental, 1997; Fujiwara and Nomura, 1999; Macedo et al., 2003; Nakahira and Arakawa, 2005), relative humidity (Tauber and Tauber, 1983; Pappas et al., 2008b) or CO2 (Gao et al., 2010) on certain life-history traits and predation capacity of chrysopids.
In Integrated Pest Management (IPM) programs, biological pest control with chrysopids could be efficiently combined with the use of selective pesticides. Vogt et al. (2001) have reviewed the effects of different management practices, such as the use of pesticides and transgenic Bacillus thuringiensis (Bt) plants on C. carnea sensu lato. There are many studies focusing on the effects of pesticides commonly used in agriculture on certain life-history traits of chrysopid species (Liu and Chen, 2000; Qi et al., 2001; Carvalho et al., 2002, 2003; Chen and Liu, 2002; Medina et al., 2003; Bueno and Freitas, 2004; Godoy et al., 2004; Ferreira et al., 2006; Silva et al., 2005, 2006; Nadel et al., 2007; Rezaei et al., 2007; Giolo et al., 2009; Mandour, 2009; Schneider et al., 2009) and on pesticide resistance development (Pathan et al., 2008, 2010; Venkatesan et al., 2009), mostly suggesting the relatively broad tolerance of chrysopids to many pesticides. Furthermore, there are several research papers reporting the effects of Bt on the performance of chrysopid species (Hilbeck et al., 1998a, b, 1999; Meier and Hilbeck, 2001; Dutton et al., 2002; Romeis et al., 2004; Pilcher et al., 2005; Rodrigo-Simon et al., 2006; Mellet and Schoeman, 2007; Sharma et al., 2007; Sisterson et al., 2007; Guo et al., 2008; Lawo and Romeis, 2008, Li et al., 2008, 2010; Mason et al., 2008).
With the attempt to evaluate the quality of certain prey species, as well as the predation efficacy of chrysopids, there are a lot of published papers mostly focusing on the performance of larvae and less on adults (Zheng et al., 1993a, b; Legaspi et al., 1994; Giles et al., 2000; Chen and Liu, 2001; Limburg and Rosenheim, 2001; Patt et al., 2003; Miller et al., 2004; Pappas et al., 2007, 2008c, 2009; Cheng et al., 2009, 2010; Huang and Enkegaard, 2010). The efficiency of chrysopids has also been also evaluated in the context of IPM revealing the biotic potential of this group of aphidophagous predators in agricultural practice (Reddy, 2001; Corrales and Campos, 2004; Furlong et al. 2004; Santos et al., 2007; Mirmoayedi and Maniee, 2008, 2009; Turquet et al., 2009).
MASS-REARING CHRYSOPIDS FOR USE IN BIOLOGICAL CONTROL
To date, the mass-rearing of commercially available chrysopids is mainly based on eggs of lepidopteran species of the genera Sitotroga, Ephestia (Anagasta) and Corcyra that have been proved as nutritionally superior and of low cost food to produce, in comparison to other artificial diets tested (Tauber et al., 2000; Riddick, 2009). Until now, any attempts to develop a sufficient and low cost artificial diet that could replace the use of prey species eggs were unsuccessful (Ridgway et al., 1970; Cohen and Smith, 1998).
Although most research projects have been focused on the development of the best food for rearing larvae, mass-rearing of adults has not received much attention (Nordlund et al., 2001). Non predaceous chrysopid adults (e.g., Chrysoperla adults) are usually reared on a proteinaceous liquid diet consisting of a mixture of protein hydrolysate, sucrose and water (Hagen and Tassan, 1970). However, when considering predaceous adults (e.g., Chrysopa adults), their mass-rearing is difficult and very costly (Tauber et al., 2000).
The evaluation of artificial and factitious foods on the performance of several chrysopid species has been the subject of many published research articles. From the mass-rearing of chrysopids perspective, several methods for the production of larvae by offering an artificial diet (Cohen and Smith, 1998; Vasquez et al., 1998; Zaki and Gesraha, 2001; Sattar et al., 2007; Sattar and Abro, 2009) or a factitious food (Singh and Varma, 1989; Pappas et al., 2007, 2008a; Matos and Obrycki, 2006; Sattar and Abro, 2009) have been tested for different chrysopid species. Mass-rearing techniques have been reviewed in extent by Tulisalo (1984), Tauber et al. (2000) and Nordlund et al. (2001).
The automation of mass-rearing systems of Chrysoperla species is still in progress with the aim to save production costs and space (McEwen et al., 1999). Such systems include suitable techniques for maintaining chrysopid adults, automatically feeding larvae and collecting eggs for mass- rearing a large number of larvae (Tauber et al., 2000). The automation of one of more steps in the mass- rearing of Chrysoperla species could result in the reduction of production costs and the enhancement of their commercial availability (Nordlund and Correa, 1995; Nordlund et al., 2001).
The short term storage of eggs and diapausing adults of certain Chrysoperla species could be achieved at low temperatures (8 to 13°C) without reduction of their quality (Tauber et al., 1993, 1997; López-Arroyo et al., 2000). Afterwards, depending on market demands and appropriate adjustment in the mass - rearing system, the immediate, predictable and synchronous completion of post - diapause development and commencement of egg laying by females could be feasible.
Another important aspect in the commercialization of chrysopid species and other natural enemies is the quality control of the individuals to be released. A first important step in the mass - rearing is the correct identification of the species to be reared, as well as the verification of it at various time periods after setting up the rearing. Chrysoperla sp. colonies are known to deteriorate after rearing for a long period of time under controlled laboratory conditions (Jones et al., 1978; Chang et al., 1996). Regular quality control inspections of the stock colony in terms of survival and reproduction performance combined with proper handling of chrysopids life - cycle (i.e., maintenance in diapause or at low temperatures) seems to be crucial for the successful marketing of chrysopids. A few studies have noted the need for setting up certain standards in the commercialization of chrysopids (Gardner and Giles, 1996; Daane and Yokota, 1997; O’Neil et al., 1998; Silvers et al., 2002).
CHRYSOPID SPECIES THAT HAVE BEEN USED IN BIOLOGICAL CONTROL
The efficiency of chrysopids as biological control agents against aphids was studied in 1742 for the first time (Senior and McEwen, 2001). Since then, the vast majority of published research articles concerning the evaluation of certain chrysopid species for use in biological control programs have been mainly focused on Chrysoperla species. These species are considered to be important biological control agents in certain agroecosystems worldwidely and they are the most commonly released commercially available chrysopids (Tauber et al., 2000). Besides Chrysoperla sp., there is also some scattered information concerning other important chrysopid species tested for use in biological control belonging to other genera, as well, such as Chrysopa, Mallada and Ceraeochrysa (Table 1).
| Table 1: | Some chrysopid species which have been studied for use in biological control under field or laboratory conditions* |
 | |
| *Reported by Ridgway and McMuphy (1984), Hagen et al. (1999), Albuquerque et al. (2001), Daane and Hagen (2001), Maisonneuve (2001), Senior and McEwen (2001) | |
| Table 2: | Pest species which have been successfully controlled by Chrysoperla species |
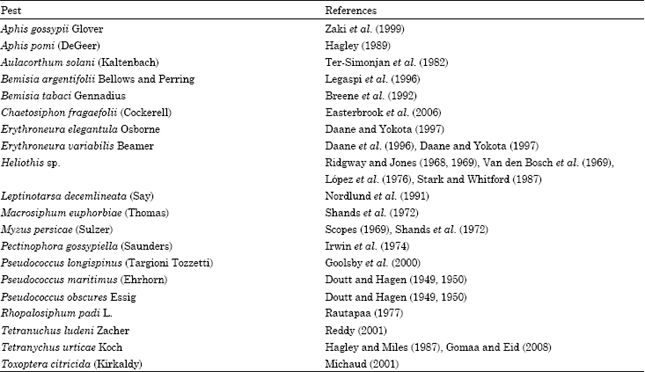 | |
Chrysoperla species have been released to control aphids in pepper, eggplant pea, potato and cotton fields. They have also been used to control Leptinotarsa decemlineata (Say) in eggplants, Panonychus ulmi (Koch) in apple orchards and Heliothis virescens (Fabricius) in cotton (Nordlund et al., 2001). In the greenhouse, C. carnea, Chrysopa septempunctata Wesmael, Chrysopa formosa Brauer and C. perla have been successfully used for aphid control in several crops, such as pepper, cucumber, eggplant, lettuce (Tulisalo, 1984). Chrysopids released in North America for biological control include mainly C. carnea to control several pests, such as mealybugs, aphids cicadellids and aphids, C. rufilabris to control chrysomelids, whiteflies, aphids and cicadellids, Chrysoperla externa (Hagen) against noctuids (Daane and Hagen, 2001) and Chrysoperla plorabunda (Fitsch) to control aphids (Michaud, 2001 and references therein). Some examples of the earliest till the latest attempts to use Chrysoperla species in biological control both in field crops as well as in greenhouses are shown in Table 2.
In the Neotropics, according to Albuquerque et al. (2001), C. externa and Ceraeochrysa species (e.g., Ceraeochrysa cubana (Hagen)) are considered to be of primary importance for the implementation of biological control programs.
In the Australian region among the very few indigenous lacewing species, M. signata is the most promising candidate for use in biological control (New, 2002), whereas in China Chrysoperla sinica (Tjeder) has already been used in augmentative release biological control programs with success (Senior and McEwen, 2001).
CHRYSOPERLA CARNEA SIBLING SPECIES PROBLEM: IMPLICATIONS FOR BIOLOGICAL CONTROL
Many species of the genus Chrysoperla, such as C. carnea, C. rufilabris and C. externa are important biological control agents (Albuquerque et al., 1994; Legaspi et al., 1994). Among the above mentioned species, C. carnea is the most studied species that has also been extensively used in releases in the context of biological control. According to recent studies, C. carnea comprises a complex of species that is generally referred to as Chrysoperla carnea Stephens sensu lato (Thierry et al., 1992; Henry et al., 2001). C. carnea species is considered to be a mega - species referred to as - carnea - group, which is one of the four species groups of the genus Chrysoperla. Although there are morphological differences between the species of the other three groups (comans -, nyerima - and pudica - group), the species belonging to the carnea - group are morphologically uniform (Brooks, 1994).
According to several studies, C. carnea consists of many reproductively isolated species that have no morphological differences and produce courtship songs of low frequency by vibrating their abdomen on the substrate when they are ready to mate. The songs are produced both by females and males, they are a prerequisite for the mating and according to some researchers they are species - specific and could be considered as a reliable index of the species identity (Henry et al., 2001; Noh and Henry, 2010).
To date, the songs of four cryptic species recorded in North America (C. plorabunda, Chrysoperla adamsi Henry, Wells and Pupedis, Chrysoperla johnsoni Henry, Wells and Pupedis and Chrysoperla downesi (Banks) sensu stricto) (Henry, 1979b, 1993; Henry et al., 1993), six in Europe and Western Asia (Chrysoperla mediterranea Hölzel, Chrysoperla lucasina (Lacroix), C. carnea (Stephens), Chrysoperla pallida Henry, Brooks, Duelli and Johnson, Chrysoperla agilis Henry, Brooks, Duelli and Johnson, Chrysoperla zastrowi arabica) (Henry et al., 1996, 1999, 2002, 2003, 2006), one in Africa (Chrysoperla zastrowi zastrowi (Esben - Petersen)) (Henry et al., 2006) and two in Eastern Asia (Chrysoperla nipponensis (Okamoto) types A and B) (Henry et al., 2009) have been described in detail. The species C. plorabunda was confirmed to be a distinct species from C. carnea although it was considered to be its synonym for a long time (Henry, 1979a, 1983, 1985a, b; Henry and Wells, 1990; Henry et al., 1993). There is a small variation between these songs even for populations originating from regions thousands of kimometers away (Henry and Wells, 1990). However, Tauber et al. (2000) argue that the description of new species based only on their songs is not reliable, since there are not enough data concerning songs variation in different seasons and geographical areas or connecting distinct songs with different biological traits.
In any case, when considering the use of Chrysoperla species in biological control there is an urgent need for a correct identification of the species to be released. There is a speculation that since C. carnea has been considered as a single species for a long period of time, sibling species may have been introduced as biological control agents in areas where another species of the - carnea group predominated (Senior and McEwen, 2001). Due to the well documented variation between the chrysopid species in certain life trait characteristics such as habitat and prey preference, seasonal life cycles and tolerance to several abiotic environmental factors (e.g., temperature or relative humidity) (Tauber et al., 2000), such releases of the inappropriate sibling species could be considered at least to some extent, as the causal factor of the recorded failures in the attempt to use chrysopids in biological control through augmentation releases.
THE USE OF CHRYSOPIDS IN BIOLOGICAL CONTROL
Field application: When considering the different biological control tactics used to control pests, four different methods could be used when releasing chrysopid species:
| • | Classical biological control, by importation of a new natural enemy |
| • | Augmentation by means of inoculative releases |
| • | Augmentation by means of inundative releases |
| • | Conservation |
In general, chrysopids have not been widely used in classical biological control and most work has been focused on their augmentation and conservation in the agroecosystems (Hagen et al., 1999). The large numbers of different chrysopid species occurring worldwidely have rendered the importation of an exotic chrysopid species to another country, useless.
Augmentation of natural enemies by means of inoculative releases aims to the provisional pest control for a long period of time through the reproduction and establishment of the natural enemy in the crop. Due to the great ability of chrysopid adults to disperse and the occasional need of immediate response in pest control, augmentation of chrysopids by means of inundative releases (mainly based on the ability of the individuals released to suppress pest populations) has been most commonly used in cases where release cost is not restrictive. A few researchers have also documented the efficacy of chrysopids in augmentation biological control (Ridgway and Jones, 1969; Daane et al., 1996; Daane and Yokota, 1997; Ehler et al., 1997; Knutson and Tedders, 2002).
Conservation techniques in biological control aim to the establishment of increased numbers of a chrysopid species within the field through the enhancement and manipulation of their habitat (e.g., crop fields). Research related with chrysopids has been mainly focused on the evaluation of certain chemicals or blends that could be sprayed on plants and act as attractants for the chrysopids and on the use of food supplements, such as artificial honeydews or pollen (Tauber et al., 2000; Nentwig et al., 2002; James, 2003, 2006; Toth et al., 2006, 2009; Venzon et al., 2006; Zhang et al., 2006; Yu et al., 2008). Other conservation methods that have application in the manipulation of chrysopids are the use of hibernation shelters for the protection of overwintering adults (McEwen and Sengonca, 2001; Wennemann, 2003; Weihrauch, 2008), cultural methods, such as intercropping (Senior and McEwen, 2001) or the use of flowering plants (Shrewsbury et al., 2004).
Release methods: Chrysopids are most commonly released as eggs or larvae, but there are also cases that adults have been used in biological control. Eggs have been tested most extensively in relation to their application in the field (Tauber et al., 2000). The release of adults has been considered to be problematic, due to the fact that they usually disperse and leave the target field before ovipositing (Duelli, 1984). This problem might be confronted by releasing adults previously fed in the insectary during preoviposition period, so as to be ready to oviposit after release in the field (Nordlund et al., 2001).
Chrysopid eggs mixed with rice hulls or vermiculite, attached on oviposition substrates or destalked are usually dispersed manually to ensure their uniform distribution in the field. Solid mediums (e.g., rice hulls) as carrying agents of the eggs have the disadvantage of not offering a good retention on the plant in comparison with liquid mediums (e.g., agar solutions) that help eggs to be easily attached on the leaves (Tauber et al., 2000). Chrysopid larvae can be released manually by placing rearing units on the plant or with the help of a paintbrush. They are also formulated mixed with rice hulls inside bottles and applied on the plants (Nordlund et al., 2001).
Advances in release methods of chrysopid eggs include the development of mechanical devices by commercial companies that use low pressure air and a liquid medium for the distribution and efficient adherence of the eggs on the plants. Furthermore, aerial application by means of model airplanes or helicopters, have been tested for use in augmentative biological control with chrysopids. The mechanical application of chrysopid eggs and larvae has been the subject of several studies mainly in terms of the effects of automation on their viability (Nordlund et al., 2001 and references therein).
Commercially available chrysopid species: To date, the most important and commonly used commercially available chrysopids are Chrysoperla species. The species C. carnea, Chrysoperla comanche Banks and C. rufilabris are being mass - reared and commercially sold by many companies in North America since years. Furthermore, C. carnea, C. externa and C. sinica are commercially available in Europe, Latin America and Asia, respectively (Tauber et al., 2000; Nordlund et al., 2001).
Concerning the different stages that could be purchased by a grower, chrysopid eggs, larvae, cocoons and adults are all commercially available by several companies and in different formulations. Eggs and larvae, usually sold in thousands, are cheaper than cocoons and adults. The top seller of all available species is C. carnea although there is a lot of concern whether this is the true C. carnea or not. According to Tauber et al. (2000), the identification of the species of the Chrysoperla stock should be the first crucial step at the onset of the culture to be mass - reared.
Besides different chrysopid formulations, food supplements, attractants as well as hibernation boxes are also available commercially to be used in conservation biological control. Food supplements for adults include bottles or bags containing yeast or pollen and nectar substitutes that could be mixed with water and applied as a paste or sprayed on the plants. By enhancing the availability of food for chrysopid adults, the companies that provide these supplements claim the increase in oviposition of the adults, their maintenance in the target field and subsequent successful pest control. Eggs of Ephestia kuehniella Zeller, is a factitious food commonly used for mass - rearing chrysopids, as well as other predatory insects. It is commercially available as a supplementary food for predatory insects within the context of biological control. In the case of chrysopids, it could be used as supplementary larval diet. However, to our knowledge there are no studies related to the benefits of using E. kuehniella eggs on the population of chrysopids under field conditions. Hibernation boxes also known as ‘lacewing chambers’ and several attractants are currently commercially available.
CHARACTERISTICS OF THE EFFICIENT BIOLOGICAL CONTROL AGENTS OF THE FAMILY CHRYSOPIDAE
The most efficient biological control agents of the genus Chrysoperla species share some common characteristics (presented below) that enhance their role in biological control, in relation to other chrysopid species.
Larval characteristics:
| • | Broad prey range: Chrysoperla sp. larvae are considered to be polyphagous and especially the larvae of commercially available species are recommended for use against several crop pests (Canard and Principi, 1984) |
| • | Easy mass - rearing: Chrysoperla larvae are easily reared on several factitious foods, such as E. kuehniella eggs and their mass - rearing has been automated by companies (Tauber et al., 2000) |
| • | Resistance to insecticides: Chrysopid larvae have been found to be highly resistant to the effects of pesticides in relation to other predatory insects (Nordlund et al., 2001). This advantage could enhance their use in IPM programs |
| • | Short developmental times: The relative rapid development of Chrysoperla larvae (Principi and Canard, 1984) is a desirable trait for their use in biological control, as it ensures the establishment of the released insects in the target crops |
| • | Predation efficiency: Chrysopid larvae consume a large amount of food so as to complete their development (Canard, 2001). For example, each C. carnea larva consumed on average 140 second instar nymphs of Myzys persicae Sulzer or 946 E. kuehniella eggs to complete its development (El-Arnaouty et al., 1996). Among larvae of chrysopid species, those of the third instar are the most voracious (Principi and Canard, 1984) |
| • | Low dispersal ability: Both chrysopid eggs and larvae are easily released in the field and the settlement of the population could be achieved due to low dispersal ability of the larvae. Furthermore, it is easier to achieve the desired number of released individuals when chrysopids are applied as larvae that are not suffering from predation by other predators as much as the eggs (Nordlund et al., 2001) |
Adult characteristics
| • | Easy mass-rearing: Chrysoperla sp. adults are not predaceous and can be easily mass - reared on a liquid diet that consists mainly of honey or pollen. This trait facilitates their rearing since there is no need for culturing an additional prey species and/or plants for the adults |
| • | High reproductive potential: The number of eggs laid by chrysopid females is affected by the quality and quantity of the larval and adult food, as well as by the environmental conditions (e.g. temperature, photoperiod, relative humidity) (Principi and Canard, 1984). In general, Chrysoperla females have a high reproductive potential. For example, the females of C. mediterranea laid approximately 2160 eggs during their lifetime at 20°C and of C. externa 2304 eggs at 25°C (Lee and Shih, 1982; Carvalho et al., 1996) |
| • | Attraction to protein hydrolysates: Mixing protein hydrolysates (mainly of yeast) with honey or sugar has been used as artificial honeydew of high quality (Hagen, 1950; Tauber et al., 2000). Furthermore, chrysopid adults are attracted to protein hydrolysates (Canard, 2001) and this behavior has been used in conservation biological control |
Besides Chrysoperla species, all of the above characteristics have been recorded in species of other genera, such as Ceraeochrysa (López-Arroyo et al., 1999a, b; Albuquerque et al., 2001), Mallada (Daane, 2001; New, 2002) and Dichochrysa (Principi, 1956; Pappas et al., 2007, 2008a, c, 2009). Furthermore, the larvae of the above mentioned genera could be protected by ants and other natural enemies by carrying ‘trash’ on their dorsa (Principi, 1956; Eisner et al., 1978). Due to the common desirable characteristics of Dichochrysa, Mallada and Ceraeochrysa with Chrysoperla species and their abundance is several crops, they could be assumed to also play an important role in biological control (López-Arroyo et al., 1999a, b; Tauber et al., 2000; Daane, 2001).
FUTURE ASPECTS AND CONSIDERATIONS IN THE USE OF CHRYSOPIDS IN BIOLOGICAL CONTROL
Within C. carnea complex many species are included that are not sufficiently studied in terms of their performance in the field as biological control agents. Given the commercial success of C. carnea, there is an urgent need other species such as C. agilis, C. lucasina, C. mediterranea or C. pallida in Europe to be included in screening studies for efficient natural enemies. Furthermore, the biology of species of the carnea compex is poorly studied, although there is scattered data to suggest their considerable variation in their performance in different habitats and biotic or abiotic factors (Tauber et al., 2000).
In order to enhance the role of Chrysopidae in biological control, the importance of other species than Chrysoperla should be evaluated. Among chrysopid species, Chrysoperla sp. are almost the only ones which are well studied and commercially available for use in biological control. By contrast, there are limited data on the biology of species belonging to the genera Dichochrysa, Mallada and Ceraeochrysa which could be also used commercially for release in biological control programs. For example, from the 130 known Dichochrysa species, only the biology of D. prasina, has been extensively studied under laboratory conditions (Pappas et al., 2007, 2008a, b, c, 2009). It has been found that D. prasina has those desirable traits that could support its amenability for mass - rearing and future commercialization (Pappas et al., 2007, 2008a, b, c, 2009). This is also the case for Ceraeochrysa and Mallada species. Ceraeochrysa cubana is considered a promising biological control agent in the Neotropics (López-Arroyo et al., 1999a, b; Albuquerque et al., 2001). Mallada signata is the only chrysopid species that has gained attention as an important indigenous predator in Australia (Horne et al., 2001), while the role of other chrysopids has been neglected (New, 2002).
Feeding habits of chrysopid species intended to be used in biological control should also be studied. They are all considered to be polyphagous predators of several soft - bodied arthropods. However, prey quality seems to be crucial for the successful completion of their immature development and subsequent adult performance (Principi and Canard, 1984). Therefore, prey preference or prey specificity of candidate chrysopid species should be further studied in detail so as they could be exploited for the control of the most suitable pests.
Intraguild Predation (IP) could severely affect the success of chrysopid releases in the field. A limited number of studies have focused on IP of C. carnea, C. rufilabris or C. prolabunda (Cisneros and Rosenhein, 1997; Lucas et al., 1997, 1998; Phoofolo and Obrycki, 1998; Dinter, 1998, 2002; White and Eigenbrode, 2000; Michaud and Grant, 2003; Mochizuki et al., 2006; Jazzar et al., 2008). The expansion of such studies to other combinations of chrysopid predators and prey species, as well as to field conditions could cast light on the optimization of chrysopid releases for biological control.
When considering the commercialization of chrysopid species, methods and rates of release should be evaluated both under laboratory and field conditions. There is limited data on the economics of chrysopid use in biological control in relation to release costs and subsequent pest populations suppress (Tauber et al., 2000). In a few studies (Gardner and Giles, 1996; Legaspi et al., 1996; Daane and Yokota, 1997), several factors, such as intraguild predation, release methods and rates that could influence the outcome of field releases have been identified. Such factors should be considered in future studies on the evaluation of the efficacy of chrysopids in the field. Studies on the improvement of conservation methods could also enhance the efficacy of naturally occurring and released chrysopids in biological control.
REFERENCES
- Albuquerque, G.S., C.A. Tauber and M.J. Tauber, 1994. Chrysoperla externa (Neuroptera: Chrysopidae): Life history and potential for biological control in Central and South America. Biol. Control, 4: 8-13.
CrossRef - Albuquerque, G.S., C.A. Tauber and M.J. Tauber, 2001. Chrysoperla externa and Ceraeochrysa spp.: Potential Use in Biological Control in the New World Tropics and Subtropics. In: Lacewings in the Crop Environment, McEwen, P.K., T.R. New and A.E. Whittington (Eds.). Cambridge University Press, Cambridge, pp: 408-423.
- Breene, R.G., Jr.R.L. Meagher, D.A. Nordlund and Y.T. Wang, 1992. Biological control of Bemisia tabaci (Homoptera: Aleyrodidae) in a greenhouse using Chrysoperla rufilabris (Neuroptera: Chrysopidae). Biol. Control, 2: 9-14.
CrossRef - Brooks, S.J., 1997. An overview of the current status of chrysopidae (Neuroptera) systematics. Deut. Entomol. Zeitschrift, 44: 267-275.
Direct Link - Bueno, A.F. and S. Freitas, 2004. Effect of the insecticides abamectin and lufenuron on eggs and larvae of Chrysoperla externa under laboratory conditions. BioControl, 49: 277-283.
Direct Link - Canard, M., 1988. Seasonal change in photoperiodic response of the larvae of the lacewing Nineta pallida. Entomol. Exp. Appl., 47: 153-159.
Direct Link - Canard, M., 1990. Effect of photoperiod on the first-instar development in the lacewing Nineta pallida. Physiol. Entomol., 15: 137-140.
Direct Link - Carvalho, C.F., M. Canard and C. Alauzet, 1996. Comparison of the Fecundities of the Neotropical Green Lacewing Chrysoperla externa (Hagen) and the West-Palearctic Chrysoperla mediterranea (Holzel) (Insecta: Neuroptera: Chrysopidae). In: Pure and Applied Research in Neuropterology, Proceedings of the 5th International Symposium on Neuropterology, Canard, M., H. Aspock and M.W. Mansell (Ed.). Giza, Egypt, pp: 103-107.
- Carvalho, C.F., B. Souza and T.M. Santos, 1998. Predation capacity and reproduction potential of Chrysoperla externa (Hagen) (Neuroptera: Chrysopidae) fed on Alabama argillacea (Hubner) eggs. Acta Zool. Fen., 209: 83-86.
Direct Link - Carvalho, G.A., C.F. Carvalho, B. Souza and J.L.R. Ulhoa, 2002. Selectivity of insecticides to Chrysoperla externa (Hagen) (Neuroptera: Chrysopidae). Neotrop. Entomol., 31: 615-621.
Direct Link - Carvalho, G.A., D. Bezerra, B. Souza and C.F. Carvalho, 2003. Effects of insecticides used on cotton crop on Chrysoperla externa (Hagen) (Neuroptera: Chrysopidae). Neotrop. Entomol., 32: 699-706.
Direct Link - Chang, Y.F., M.J. Tauber and C.A. Tauber, 1995. Storage of the mass-produced predator Chrysoperla carnea (Neuroptera: Chrysopidae): Influence of photoperiod, temperature and diet. Environ. Entomol., 24: 1365-1374.
Direct Link - Chang, Y.F., M.J. Tauber and C.A. Tauber, 1996. Reproduction and quality of F1 offspring in Chrysoperla carnea: Differential influenceof quiescence, artificially-induced diapause and natural diapause. J. Insect Physiol., 42: 521-528.
CrossRef - Chen, T.Y. and T.X. Liu, 2001. Relative consumption of three aphid species by the lacewing, Chrysoperla rufilabris and effects on its development and survival. BioControl, 46: 481-491.
Direct Link - Chen, T.Y. and T.X. Liu, 2002. Susceptibility of immature stages of Chrysoperla rufilabris (Neurop., Chrysopidae) to pyriproxyfen, a juvenile hormone analog. J. Applied Entomol., 126: 125-129.
Direct Link - Cheng, L.L., J.R. Nechols, D.C. Margolies, J.F. Campbell, P.S. Yang, C.C. Chen and C.T. Chiu, 2009. Foraging on and consumption of two species of papaya pest mites, Tetranychus kanzawai and Panonychus citri (Acari: Tetranychidae), by Mallada basalis (Neuroptera: Chrysopidae). Environ. Entomol., 38: 715-722.
CrossRef - Cheng, L.L., J.R. Nechols, D.C. Margolies, J.F. Campbell and P.S. Yang, 2010. Assessment of prey preference by the mass-produced generalist predator, Mallada basalis (Walker) (Neuroptera: Chrysopidae), when offered two species of spider mites, Tetranychus kanzawai Kishida and Panonychus citri (McGregor) (Acari: Tetranychidae), on papaya. Biol. Control, 53: 267-272.
CrossRef - Cisneros, J. and J. Rosenheim, 1997. Ontogenetic change of prey preference in the generalist predator Zelus renardii and its influence on predator-predator interactions. Ecol. Entomol., 22: 399-407.
CrossRefDirect Link - Cohen, A.C. and L.K. Smith, 1998. A new concept in artificial diets for Chrysoperla rufilabris: The efficacy of solid diets. Biol. Control, 13: 49-54.
CrossRef - Corrales, N. and M. Campos, 2004. Populations, longevity, mortality and fecundity of Chrysoperla carnea (Neuroptera, Chrysopidae) from olive-orchards with different agricultural management systems. Chemosphere, 57: 1613-1619.
CrossRef - Daane, K.M., G.Y. Yokota, Y. Zheng and K.S. Hagen, 1996. Inundative release of common green lacewings (Neuroptera: Chrysopidae) to suppress Erythroneura variabilis and E. elegantula (Homoptera: Cicadellidae) in vineyards. Environ. Entomol., 25: 1224-1234.
Direct Link - Daane, K.M and G.Y. Yokota, 1997. Release strategies affect survival and distribution of green lacewings (Neuroptera: Chrysopidae) in augmentation programs. Environ. Entomol., 26: 455-464.
Direct Link - Dinter, A., 1998. Intraguild predation between erigonid spiders, lacewing larvae and carabids. J. Applied Entomol., 122: 163-167.
CrossRef - Dinter, A., 2002. Microcosm studies on intraguild predation between female erigonid spiders and lacewing larvae and influence of single versus multiple predators on cereal aphids. J. Applied Entomol., 126: 249-257.
Direct Link - Doutt, R.L. and K.S. Hagen, 1949. Periodic colonization of Chrysopa californica as a possible control of mealybugs. J. Econ. Entomol., 42: 560-561.
CrossRefDirect Link - Doutt, R.L. and K.S. Hagen, 1950. Biological control measures applied against Pseudococcus maritimus on pears. J. Econ. Entomol., 43: 94-96.
Direct Link - Dutton, A., H. Klein, J. Romeis and F. Bigler, 2002. Uptake of Bt-toxin by herbivores feeding on transgenic maize and consequences for the predator Chrysoperla carnea. Ecol. Entomol., 27: 441-447.
Direct Link - Easterbrook, M.A., J.D. Fitzgerald and M.G. Solomon, 2006. Suppression of aphids on strawberry by augmentative releases of larvae of the lacewing Chrysoperla carnea (Stephens). Biocontrol Sci. Technol., 16: 893-900.
Direct Link - Eisner, T., K. Hicks, M. Eisner and D.S. Robson, 1978. Wolf-in-sheep's-clothing strategy of a predaceous insect larva. Science, 199: 790-794.
Direct Link - El-Arnaouty, S.A., A. Ferran and V. Beyssat-Arnaouty, 1996. Food Consumption by Chrysoperla carnea (Stephens) and Chrysoperla sinica (Tjeder) of Natural and Substitute Prey: Determination of Feeding Efficiency (Insecta: Neuroptera: Chrysopidae). In: Pure and Applied Research in Neuropterology, Proceedings of the 5th International Symposium on Neuropterology, Canard, M., H. Aspock and M.W. Mansell (Eds.). Giza, Egypt, pp: 109-117.
- Ferreira, A.J., G.A. Carvalho, M. Botton and O. Lasmar, 2006. Selectivity of insecticides used in apple orchards to two populations of Chrysoperla externa (Hagen, 1861) (Neuroptera: Chrysopidae). Cienc. Rural, 36: 378-384.
Direct Link - Fujiwara, C. and M. Nomura, 1999. Effects of photoperiod and temperature on larval development of Chrysoperla carnea Stephens (Neuroptera: Chrysopidae). Jap. J. Applied Entomol. Zool., 43: 175-179.
Direct Link - Furlong, M.J., Z. H. Shi, Y.Q. Liu, S.J. Guo, Y.B. Lu, S.S. Liu and M.P. Zalucki, 2004. Experimental analysis of the influence of pest management practice on the efficacy of an endemic arthropod natural enemy complex of the diamondback moth. J. Econ. Entomol., 97: 1814-1827.
PubMed - Gao, F., F. Chen and F. Ge, 2010. Elevated CO2 lessens predation of Chrysopa sinica on Aphis gossypii. Entomol. Exp. Appl., 135: 135-140.
Direct Link - Gardner, J. and K. Giles, 1996. Handling and environmental effects on viability of mechanically dispensed green lacewing eggs. Biol. Control, 7: 245-250.
CrossRef - Giles, K.L., R.D. Madden, M.E. Payton and J.W. Dillwith, 2000. Survival and development of Chrysoperla rufilabris (Neuroptera: Chrysopidae) supplied with pea aphids (Homoptera: Aphididae) reared on alfalfa and faba bean. Environ. Entomol., 29: 304-311.
Direct Link - Giolo, F.P., P. Medina, A.D. Grutzmacher and E. Vinuela, 2009. Effects of pesticides commonly used in peach orchards in Brazil on predatory lacewing Chrysoperla carnea under laboratory conditions. BioControl, 54: 625-635.
Direct Link - Godoy, M.S., G.A. Carvalho, J.C. Moraes, L.V. Cosme, M.M. Goussain, C.F. Carvalho and A.A. Morais, 2004. Selectivity of six insecticides used in citrus crops on pupae and adults of Chrysoperla externa (Hagen) (Neuroptera: Chrysopidae). Neotrop. Entomol., 33: 359-364.
Direct Link - Grimal, A., 1988. Photoperiodic developmental requirements of the lacewing Tjederina gracilis. Entomol. Exp. Appl., 47: 189-194.
Direct Link - Guo, J.Y., F.H. Wan, L. Dong, G.L. Lovei and Z.J. Han, 2008. Tri-trophic interactions between Bt cotton, the herbivore Aphis gossypii glover (Homoptera: Aphididae) and the predator Chrysopa pallens (Rambur) (Neuroptera: Chrysopidae). Environ. Entomol., 37: 263-270.
Direct Link - Hagen, K.S., N.J. Mills, G. Gordh and J.A. McMurtry, 1999. Terrestrial Arthropod Predators of Insect and Mite Pests, Chrysopidae. In: Handbook of Biological Control: Principles and Applications of Biological Control, Bellows T.S. and T.W. Fisher (Eds.). Academic Press, San Diego, California, pp: 415-423.
- Henry, C.S., 1979. Acoustical communication during courtship and mating in the green lacewing Chrysopa carnea (Neuroptera: Chrysopidae). Ann. Entom. Soc. Am., 72: 68-79.
Direct Link - Henry, C.S., 1979. The courtship call of Chrysopa downesi Banks (Neuroptera: Chrysopidae): Its evolutionary significance. Psyche, 86: 291-297.
Direct Link - Henry, C.S., 1983. Acoustic recognition of sibling species within the Holarctic lacewing Chrysoperla carnea (Neuroptera: Chrysopidae). Syst. Entomol., 8: 293-301.
CrossRef - Henry, C.S., 1985. The proliferation of cryptic species in Chrysoperla green lacewings through song divergence. Florida Entomol., 68: 18-38.
Direct Link - Henry, C.S., 1985. Sibling species, call differences and speciation in green lacewings (Neuroptera: Chrysopidae: Chrysoperla). Evolution, 39: 965-984.
Direct Link - Henry, C.S., 1993. Chrysoperla johnsoni Henry (Neuroptera: Chrysopidae): Acoustic evidence for full species status. Ann. Entomol. Soc. Am., 86: 14-25.
Direct Link - Henry, C.S. and M.M. Wells, 1990. Geographical variation in the song of Chrysoperla plorabunda (Neuroptera: Chrysopidae) in north America. Ann. Entomol. Soc. Am., 83: 317-325.
Direct Link - Henry, C.S., M.M. Wells and R.J. Pupedis, 1993. Hidden taxonomic diversity within Chrysoperla plorabunda (Neuroptera: Chrysopidae): Two new species based on courthship songs. Ann. Entomol. Soc. Am., 86: 1-13.
Direct Link - Henry, C.S., S.J. Brooks, J.B. Johnson and P. Duelli, 1996. Chrysoperla lucasina (Lacroix): A distinct species of green lacewing, confirmed by acoustical analysis (Neuroptera: Chrysopidae). Syst. Entomol., 21: 205-218.
Direct Link - Henry, C.S., S.J. Brooks, D. Thierry, P. Duelli and J.B. Johnson, 2001. The Common Green Lacewing (Chrysoperla carnea s. lat.) and the Sibling Species Problem. In: Lacewings in the Crop Environment, McEwen, P.K., T.R. New and A.E. Whittington (Eds.). Cambridge University Press, Cambridge, pp: 29-42.
- Henry, C.S., M.L.M. Wells and K.E. Holsinger, 2002. The inheritance of mating songs in two cryptic, sibling lacewing species (Neuroptera: Chrysopidae: Chrysoperla). Genetica, 116: 269-289.
Direct Link - Henry, C.S., S.J. Brooks, P. Duelli and J.B. Johnson, 2003. A lacewing with the wanderlust: The European song species Maltese, Chrysoperla agilis, sp.n., of the carnea group of Chrysoperla (Neuroptera: Chrysopidae.). Syst. Entomol., 28: 131-148.
CrossRef - Henry, C.S., S.J. Brooks, P. Duelli and J.B. Johnson, 2006. Courtship song of the South African lacewing Chrysoperla zastrowi (Esben-Petersen) (Neuroptera: Chrysopidae): Evidence for a trans-equatorial geographic range?. J. Nat. Hist., 40: 2173-2195.
Direct Link - Henry, C.S., A. Mochizuki, K. Nakahira, N. Haruyama and M. Nomura, 2009. Courtship songs of Chrysoperla nipponensis (Νeuroptera: Chrysopidae) delineate two distinct biological species in eastern Asia. Ann. Entomol. Soc. Am., 102: 747-758.
Direct Link - Hilbeck, A., M. Baumgartner, P.M. Fried and F. Bigler, 1998. Effects of transgenic Bacillus thuringiensis corn-fed prey on mortality and development time of immature Chrysoperla carnea (Neuroptera: Chrysopidae). Environ. Entomol., 27: 480-487.
Direct Link - Hilbeck, A., W.J. Moar, M.P. Carey, A. Filippini and F. Bigler, 1998. Toxicity of Bacillus thuringiensis Cry1Ab toxin to the predator Chrysoperla carnea (Neuroptera: Chrysopidae). Environ. Entomol., 27: 1255-1263.
Direct Link - Hilbeck, A., W.J. Moar, M. Pusztai-Carey, A. Filippini and F. Bigler, 1999. Prey-mediated effects of Cry1Ab toxin and protoxin and Cry2A protoxin on the predator Chrysoperla carnea. Entomol. Exp. Appl., 91: 305-316.
Direct Link - Hodek, I. and A. Honek, 1976. Action of photoperiod at cold in diapausing Chrysopa carnea females (Chrysopidae, Neuroptera). Oecologia, 25: 309-311.
Direct Link - Honek, A. and F. Koucourek, 1988. Thermal requirements for development of aphidophagous Coccinellidae (Coleoptera), Chrysopidae, Hemerobiidae (Neuroptera) and Syrphidae (Diptera): Some general trends. Oecologia, 76: 455-460.
Direct Link - Huang, N. and A. Enkegaard, 2010. Predation capacity and prey preference of Chrysoperla carnea on Pieris brassicae. BioControl, 55: 379-385.
Direct Link - Irwin, M.E., R.W. Gill and D. Gonzalez, 1974. Field-cage studies of native egg predators of the pink bollworm in southern California cotton. J. Econ. Entomol., 67: 193-196.
Direct Link - James, D.G., 2003. Field evaluation of herbivore-induced plant volatiles as attractants for beneficial insects: Methyl salicylate and the green lacewing, Chrysopa nigricornis. J. Chem. Ecol., 29: 1601-1609.
CrossRef - James, D.G., 2006. Methyl salicylate is a field attractant for the goldeneyed lacewing, Chrysopa oculata. Biocontrol Sci. Technol., 16: 107-110.
Direct Link - Jazzar, C., R. Meyhofer, L. Ebssa and H.M. Poehling, 2008. Two protagonists on aphidophagous patches: Effects of learning and intraguild predation. Entomol. Exp. Appl., 127: 88-99.
CrossRef - Jones, S.L., R.E. Kinzer, D.L. Bull, J.R. Ables and R.L. Ridway, 1978. Deterioration of Chrysopa carnea in mass culture. Ann. Entomol. Soc. Am., 71: 160-162.
Direct Link - Lawo, N.C. and J. Romeis, 2008. Assessing the utilization of a carbohydrate food source and the impact of insecticidal proteins on larvae of the green lacewing, Chrysoperla carnea. Biol. Control, 44: 389-398.
CrossRef - Legaspi, J.C., D.A. Nordlund and B.C. Legaspi Jr., 1996. Tri-trophic interactions and predation rates in Chrysoperla spp. attacking the silverleaf whitefly. Southwest. Entomol., 21: 33-42.
Direct Link - Li, Y., M. Meissle and J. Romeis, 2010. Use of maize pollen by adult Chrysoperla carnea (Neuroptera: Chrysopidae) and fate of Cry proteins in Bt-transgenic varieties. J. Insect Physiol., 56: 157-164.
Direct Link - Limburg, D.D. and J.A. Rosenheim, 2001. Extrafloral nectar consumption and its influence on survival and development of an omnivorous predator, larval Chrysoperla plorabunda (Neuroptera: Chrysopidae). Environ. Entomol., 30: 595-604.
Direct Link - Liu, T.X. and T.Y. Chen, 2000. Effects of the chitin synthesis inhibitor buprofezin on survival and development of immatures of Chrysoperla rufilabris (Neuroptera: Chrysopidae). J. Econ. Entomol., 93: 234-239.
CrossRef - Lopez, J.D., Jr.R.L. Ridgway and R.E. Pinnell, 1976. Comparative efficacy of four insect predators of the bollworm and tobacco budworm. Environ. Entomol., 5: 1160-1164.
Direct Link - Lopez-Arroyo, J.I., C.A. Tauber and M.J. Tauber, 1999. Comparative life histories of the predators Ceraeochrysa cincta, C. cubana and C. smithi (Neuroptera: Chrysopidae). Ann. Entomol. Soc. Am., 92: 208-217.
Direct Link - Mantoanelli, E., G.S. Albuquerque, C.A. Tauber and M.J. Tauber, 2006. Leucochrysa (Leucochrysa) varia (Neuroptera: Chrysopidae): Larval descriptions, developmental rates and adult color variation. Ann. Entomol. Soc. Am., 99: 7-18.
Direct Link - Mason, C.E., J.K. Sheldon, J. Pesek, H. Bacon, R. Gallusser, G. Radke and B. Slabauch, 2008. Assessment of Chrysoperla plorabunda longevity, fecundity and egg viability when adults are fed transgenic Bt corn pollen. J. Agric. Urban Entomol., 25: 265-278.
Direct Link - Matos, B. and J.J. Obrycki, 2006. Prey suitability of Galerucella calmariensis L. (Coleoptera: Chrysomelidae) and Myzus lythri (Schrank) (Homoptera: Aphididae) for development of three predatory species. Environ. Entomol., 35: 345-350.
Direct Link - McEwen, P.K., N.A.C. Kidd, E. Bailey and L. Eccleston, 1999. Small-scale production of the common green lacewing Chrysoperla carnea (Stephens) (Neuropt., Chrysopidae): Minimizing costs and maximizing output. J. Applied Entomol., 123: 303-306.
CrossRef - Medina, P., G. Smagghe, F. Budia, L. Tirry and E. Vinuela, 2003. Toxicity and absorption of azadirachtin, diflubenzuron, pyriproxyfen and tebufenozide after topical application in predatory larvae of Chrysoperla carnea (Neuroptera: Chrysopidae). Environ. Entomol., 32: 196-203.
Direct Link - Meier, M.S. and A. Hilbeck, 2001. Influence of transgenic Bacillus thuringiensis corn-fed prey on prey preference of immature Chrysoperla carnea (Neuroptera: Chrysopidae). Basic Applied Ecol., 2: 35-44.
CrossRef - Mellet, M.A. and AS. Schoeman, 2007. Effect of Bt-cotton on chrysopids, ladybird beetles and their prey: Aphids and whiteflies. Ind. J. Exp. Biol., 45: 554-562.
Direct Link - Michaud, J.P., 2001. Evaluation of green lacewings, Chrysoperla plorabunda (Fitch) (Neurop., Chrysopidae), for augmentative release against Toxoptera citricida (Hom., Aphididae) in citrus. J. Applied Entomol., 125: 383-388.
CrossRef - Michaud, J.P. and A.K. Grant, 2003. Intraguild predation among ladybeetles and a green lacewing: Do the larval spines of Curinus coeruleus (Coleoptera: Coccinellidae) serve a defensive function. Bull. Entomol. Res., 93: 499-505.
Direct Link - Milbrath, L.R., M.J. Tauber and C.A. Tauber, 1993. Prey specificity in Chrysopa: An interspecific comparison of larval feeding and defensive behavior. Ecology, 74: 1384-1393.
Direct Link - Miller, G.L., J.D. Oswald and D.R. Miller, 2004. Lacewings and scale insects: A review of predator/prey associations between the Neuropterida and Coccoidea (Insecta: Neuroptera, Raphidioptera, Hemiptera). Ann. Entomol. Soc. Am., 97: 1103-1125.
Direct Link - Mirmoayedi, A. and M. Maniee, 2008. Integrated control of pod borer, Helicoverpa armigera (Hub.) (Lepidoptera, Noctuidae), releasing Chrysoperla lucasina (Neuroptera, Chrysopidae) and treatment of insecticides. J. Entomol., 5: 322-327.
CrossRefDirect Link - Mirmoayedi, A. and M. Maniee, 2009. Integrated pest management of cotton's spiny bollworm (Earias insulana) with spray of diazinon and relaese of green lacewings. J. Entomol., 6: 56-61.
CrossRefDirect Link - Mochizuki, A., H. Naka, K. Hamasaki and T. Mitsunaga, 2006. Larval cannibalism and intraguild predation between the introduced green lacewing, Chrysoperla carnea and the indigenous trash-carrying green lacewing, Mallada desjardinsi (Neuroptera: Chrysopidae), as a case study of potential nontarget effect assessment. Environ. Entomol., 35: 1298-1303.
Direct Link - Nadel, H., M.W. Johnson, M. Gerik and K.M. Daane, 2007. Ingestion of spinosad bait GF-120 and resulting impact on adult Chrysoperla carnea (Neuroptera: Chrysopidae). Biocontrol. Sci., Technol., 17: 995-1008.
CrossRef - Nakahira, K. and R. Arakawa, 2005. Effect of photoperiod on the development and diapause of the green lacewing Chrysopa pallens (Neuroptera: Chrysopidae). Entomol. Sci., 8: 133-135.
CrossRef - Nakahira, K., R. Nakahara and R. Arakawa, 2005. Effect of temperature on development, survival and adult body size of two green lacewings, Mallada desjardinsi and Chrysoperla nipponensis (Neuroptera: Chrysopidae). Applied Entomol. Zool., 40: 615-620.
Direct Link - Nechols, J.R., M.J. Tauber and C.A. Tauber, 1987. Geographical variability in ecophysiological traits controlling dormancy in Chrysopa oculata (Neuroptera: Chrysopidae). J. Insect Physiol., 33: 627-633.
CrossRef - New, T.R., 2002. Prospects for extending the use of Australian lacewings in biological control. Acta Zool. Acad. Sci. Hung., 48: 209-216.
Direct Link - Noh, S. and C.S. Henry, 2010. Sexually monomorphic mating preferences contribute to premating isolation based on song in European green lacewings. Evolution, 64: 261-270.
CrossRef - Nordlund, D.A. and J.A. Correa, 1995. Improvements in the production systems for green lacewings: An adult feeding and oviposition unit and hot wire egg harvesting system. Biol. Control, 5: 179-188.
CrossRef - O'Neil, R.J., K.L. Giles J.J. Obrycki, D.L. Mahr, J.C. Legaspi and K. Katovich, 1998. Evaluation of the quality of four commercially available natural enemies. Biol. Control, 11: 1-8.
CrossRef - Pappas, M.L., G.D. Broufas and D.S. Koveos, 2007. Effects of various prey species on development, survival and reproduction of the predatory lacewing Dichochrysa prasina (Neuroptera: Chrysopidae). Biol. Control, 43: 163-170.
CrossRef - Pappas, M.L., G.D. Broufas and D.S. Koveos, 2008. Effect of temperature on survival, development and reproduction of the predatory lacewing Dichochrysa prasina (Neuroptera: Chrysopidae) reared on Ephestia kuehniella eggs (Lepidoptera: Pyralidae). Biol. Control, 45: 396-403.
CrossRef - Pappas, M.L., G.D. Broufas and D.S. Koveos, 2008. Effect of relative humidity on development, survival and reproduction of the predatory lacewing Dichochrysa prasina (Neuroptera: Chrysopidae). Biol. Control, 46: 234-241.
CrossRef - Pappas M.L., G.D. Broufas and D.S. Koveos, 2008. The two spotted spider mite, Tetranychus urticae (Acari: Tetranychidae), alternative prey for the lacewing Dichochrysa prasina (Neuroptera: Chrysopidae). Eur. J. Entomol., 105: 461-466.
Direct Link - Pappas M.L., G.D. Broufas and D.S. Koveos, 2009. Effect of prey availability on development and reproduction of the predatory lacewing Dichochrysa prasina (Neuroptera: Chrysopidae). Ann. Entomol. Soc. Am., 102: 437-444.
CrossRefDirect Link - Pathan, A.K., A.H. Sayyed, M. Aslam, M. Razaq, G. Jilani and M.A. Saleem, 2008. Evidence of field-evolved resistance to organophosphates and pyrethroids in Chrysoperla carnea (Neuroptera: Chrysopidae). J. Econ. Entomol., 101: 1676-1684.
Direct Link - Pathan, A.K., A.H. Sayyed, M. Aslam, T.X. Liu, M. Razaq and W.A. Gillani, 2010. Resistance to pyrethroids and organophosphates increased fitness and predation potential of Chrysoperla carnae (Neuroptera: Chrysopidae). J. Econ. Entomol., 103: 823-834.
CrossRefPubMedDirect Link - Patt, J.M., S.C. Wainright, G.C. Hamilton, D. Whittinghill, K. Bosley, J. Dietrick and J.H. Lashomb, 2003. Assimilation of carbon and nitrogen from pollen and nectar by a predaceous larva and its effects on growth and development. Ecol. Entomol., 28: 717-728.
CrossRef - Phoofolo, M.W. and J.J. Obrycki, 1998. Potential for intraguild predation and competition among predatory Coccinellidae and Chrysopidae. Entomol. Exp. Appl., 89: 47-55.
CrossRef - Pilcher, C.D., M.E. Rice and J.J. Obrycki, 2005. Impact of transgenic Bacillus thuringiensis corn and crop phenology on five nontarget arthropods. Environ. Entomol., 34: 1302-1316.
Direct Link - Qi, B., G. Gordon and W. Gimme, 2001. Effects of neem-fed prey on the predacious insects Harmonia conformis (Boisduval) (Coleoptera: Coccinellidae) and Mallada signatus (Schneider) (Neuroptera: Chrysopidae). Biol. Control, 22: 185-190.
Direct Link - Reddy, G.V.P., 2001. Comparative effectiveness of an integrated pest management system and other control tactics for managing the spider mite Tetranychus ludeni (Acari: Tetranychidae) on eggplant. Exp. Applied Acarol., 25: 985-992.
CrossRef - Rezaei, M., K. Talebi, V.H. Naveh and A. Kavousi, 2007. Impacts of the pesticides imidacloprid, propargite, and pymetrozine on Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae): IOBC and life table assays. BioControl, 52: 385-398.
CrossRef - Riddick, E.W., 2009. Benefits and limitations of factitious prey and artificial diets on life parameters of predatory beetles, bugs, and lacewings: A mini-review. BioControl, 54: 325-339.
CrossRef - Ridgway, R.L. and S.L. Jones, 1968. Field-cage releases of Chrysopa carnea for suppression of populations of the bollworm and tobacco bollworm on cotton. J. Econ. Entomol., 61: 895-898.
Direct Link - Ridgway, R.L. and S.L. Jones, 1969. Inundative releases of Chrysopa carnea for control of Heliothis on cotton. J. Econ. Entomol., 62: 177-180.
Direct Link - Rodrigo-Simon, A., R.A. De Maagd, C. Avilla, P.L. Bakker, J. Molthoff, J.E. Gonzalez-Zamora and J. Ferre, 2006. Lack of detrimental effects of Bacillus thuringiensis Cry toxins on the insect predator Chrysoperla carnea: A toxicological, histopathological, and biochemical analysis. Applied Environ. Microbiol., 72: 1595-1603.
CrossRef - Romeis, J., A. Dutton and F. Bigler, 2004. Bacillus thuringiensis toxin (Cry1Ab) has no direct effect on larvae of the green lacewing Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae). J. Insect Physiol., 50: 175-183.
PubMed - Silva, P.S., G.S. Albuquerque, C.A. Tauber and M.J. Tauber, 2007. Life history of a widespread Neotropical predator, Chrysopodes (Chrysopodes) lineafrons (Neuroptera: Chrysopidae). Biol. Control, 41: 33-41.
CrossRef - Tauber, M.J. and C.A. Tauber, 1972. Larval diapause in Chrysopa nigricornis: Sensitive stages, critical photoperiod and termination (Neuroptera: Chrysopidae). Entomol. Exp. Appl., 15: 105-111.
CrossRefDirect Link - Tauber, C.A., M.J. Tauber and J.R. Nechols, 1987. Thermal requirements for development in Chrysopa oculata: A geographical stable trait. Ecology, 68: 1479-1487.
Direct Link - Tauber, M.J., C.A. Tauber and S. Gardescu, 1993. Prolonged storage of Chrysoperla carnea (Neuroptera: Chrysopidae). Environ. Entomol., 22: 843-848.
Direct Link - Tauber, M.J., G.S. Albuquerque and C.A. Tauber, 1997. Storage of nondiapausing Chrysoperla externa adults: Influence on survival and reproduction. Biol. Control, 10: 69-72.
Direct Link - Tauber, M.J., C.A. Tauber and T.W. Hilton, 2006. Life history and reproductive behavior of the endemic Hawaiian Anomalochrysa hepatica (Neuroptera: Chrysopidae): A comparative approach. Eur. J. Entomol., 103: 327-336.
Direct Link - Toth, M., F. Szentkiralyi, J. Vuts, A. Letardi, M.R. Tabilio, G. Jaastad and G.K. Knudsen, 2009. Optimization of a phenylacetaldehyde-based attractant for common green lacewings (Chrysoperla carnea s.l.). J. Chem. Ecol., 35: 449-458.
CrossRefPubMedDirect Link - Thierry, D., R. Cloupeau and M. Jarry, 1992. La Chrysope commune Chrysoperla carnea (Stephens) sensu lato dans la centre de la France: Mise en evidence d'un complexe d'especes (Insecta: Neuroptera: Chrysopidae). In: Current Research in Neuropterology, Proceedings of the 4th International Symposium on Neuropterology, Canard, M., H. Aspock and M.W. Mansell (Ed.). Sacco, Toulouse, pp: 379-392.
- Toth, M., A. Bozsik, F. Szentkiralyi, A. Letardi and M.R. Tabilio et al., 2006. Phenylacetaldehyde: A chemical attractant for common green lacewings (Chrysoperla carnea s.l., Neuroptera: Chrysopidae). Eur. J. Entomol., 103: 267-271.
Direct Link - Turquet, M., J.J. Pommier, M. Piron, E. Lascaux and G. Lorin, 2009. Biological control of aphids with Chrysoperla carnea on strawberry. Acta Hortic., 842: 641-644.
Direct Link - Venkatesan, T., S.K. Jalali, K.S. Murthy, R.J. Rabindra and Y. Lalitha, 2009. Occurrence of insecticide resistance in field populations of Chrysoperla zastrowi Arabica (Neuroptera: Chrysopidae) in India. Indian J. Agric. Sci., 79: 910-912.
Direct Link - Venzon, M., M.C. Rosado, D.E. Euzebio, B. Souza and J.H. Schoereder, 2006. Suitability of leguminous cover crop pollens as food source for the green lacewing Chrysoperla externa (Hagen) (Neuroptera: Chrysopidae). Neotrop. Entomol., 35: 371-376.
PubMed - Villenave, J., D. Thierry, A.A. Mamun, T. Lode and E. Rat-Morris, 2005. The pollens consumed by common green lacewings Chrysoperla spp. (Neuroptera: Chrysopidae) in cabbage crop environment in Western France. Eur. J. Entomol., 102: 547-552.
Direct Link - Volkovich, T.A. and N.A. Blumental, 1997. Photo-thermoperiodic responses in some species of lacewings (Chrysopidae): Their role in diapause induction. Eur. J. Entomol., 94: 435-444.
Direct Link - Weihrauch, F., 2008. Overwintering of common green lacewings in hibernation shelters in the Hallertau hop growing area. Bull. Insectol., 61: 67-71.
Direct Link - Weisman, S., S. Okada, S.T. Mudie, M.G. Huson, H.E. Trueman, V.S. Haritos and T.D. Sutherland, 2009. Fifty years later: The sequence, structure and function of lacewing cross-beta silk. J. Struct. Biol., 168: 467-475.
PubMed - White, C. and S.D. Eigenbrode, 2000. Leaf surface waxbloom in Pisum sativum influences predation and intra-guild interactions involving two predator species. Oecologia, 124: 252-259.
CrossRefDirect Link - Yu, H., Y. Zhang, K. Wu, X.W. Gao and Y.Y. Guo, 2008. Field-testing of synthetic herbivore-induced plant volatiles as attractants for beneficial insects. Environ. Entomol., 37: 1410-1415.
CrossRefDirect Link - Zaki, F.N., M.F. El-Shaarawy and N.A. Farag, 1999. Release of two predators and two parasitoids to control aphids and whiteflies. J. Pest Sci., 72: 19-20.
CrossRefDirect Link - Zaki, F.N. and M.A. Gesraha, 2001. Production of the green lacewing Chrysoperla caranea (Steph.) (Neuropt., Chrysopidae) reared on semi-artificial diet based on the algae, Chlorella vulgaris. J. Applied Entomol., 125: 97-98.
CrossRefDirect Link - Zhang, Q.H., M. Sheng, G. Chen, J.R. Aldrich and K.R. Chauhan, 2006. Iridodial: A powerful attractant for the green lacewing, Chrysopa septempunctata (Neuroptera: Chrysopidae). Naturwissenschaften, 93: 461-465.
CrossRefDirect Link - Zheng, Y., K.S. Hagen, K.M. Daane and T.E. Mittler, 1993. Influence of larval dietary supply on the food consumption, food utilization efficiency, growth and development of the lacewing Chrysoperla carnea. Entomol. Exp. Appl., 67: 1-7.
CrossRefDirect Link - Zheng, Y., K.M. Daane, K.S. Hagen and T.E. Mittler, 1993. Influence of larval food consumption on the fecundity of the lacewing Chrysoperla carnea. Entomol. Exp. Appl., 67: 9-14.
CrossRef - Dong, K.Z., W.Z. Li, J.Z. Cui and X.K. Yang, 2004. Three new species of Dichochrysa (Insecta: Neuroptera: Chrysopidae) from China, with a checklist of Chinese Dichochrysa. Raffles. Bull. Zool., 52: 67-74.
Direct Link