
Shanmugam Manoharan
Department of Biochemistry, Faculty of Science, Annamalai University, Annamalainagar-608 002, Tamil Nadu, India
Simon Silvan
Department of Biochemistry, Faculty of Science, Annamalai University, Annamalainagar-608 002, Tamil Nadu, India
Krishnamoorthi Vasudevan
Department of Zoology, Faculty of Science, Annamalai University, Annamalainagar-608 002, Tamil Nadu, India
Subramanian Balakrishnan
Department of Biochemistry, Faculty of Science, Annamalai University, Annamalainagar-608 002, Tamil Nadu, India
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 1 | Page No.: 161-168







ABSTRACT
Piper longum (Piperaceae family) is used in Indian traditional medicine as a remedy for various disorders including diabetes mellitus. The aim of the present study was to investigate the antihyperglycemic and antilipidperoxidative effects of ethanolic extract of Piper longum dried fruits (PLEFet) in alloxan induced diabetic rats. Diabetes mellitus was induced in overnight fasted (12 h) Wistar rats by single intraperitoneal injection of freshly prepared solution of alloxan monohydrate (150 mg kg-1 bw) in physiological saline. The blood glucose level, carbohydrate metabolizing enzymes and the status of lipid peroxidation and antioxidants were assayed using specific colorimetric methods. Oral administration of PLEFet has shown significant antihyperglycemic, antilipidperoxidative and antioxidant effects in diabetic rats. PLEFet also corrected the metabolic alterations observed by the activities of several carbohydrate metabolizing enzymes (hexokinase, glucose-6-phosphatase, glucose-6-phosphate dehydrogenase, fructose 1.6-bisphosphatase and glycogen phosphorylase) in alloxan induced diabetic rats. The antihyperglycemic effect of PLEFet is comparable to that of the standard reference drug, glibenclamide. Present results indicate that PLEFet has potent antihyperglycemic and antilipidperoxidative effects in alloxan induced diabetic rats. PLEFet can therefore be used as an alternative remedy for diabetes and oxidative stress associated diabetic complications.
PDF Abstract XML References Citation
How to cite this article
Shanmugam Manoharan, Simon Silvan, Krishnamoorthi Vasudevan and Subramanian Balakrishnan, 2007. Antihyperglycemic and Antilipidperoxidative Effects of Piper longum (Linn.)Dried Fruits in Alloxan Induced Diabetic Rat. Journal of Biological Sciences, 7: 161-168.
DOI: 10.3923/jbs.2007.161.168
URL: https://scialert.net/abstract/?doi=jbs.2007.161.168
DOI: 10.3923/jbs.2007.161.168
URL: https://scialert.net/abstract/?doi=jbs.2007.161.168
INTRODUCTION
Diabetes mellitus, one of the leading metabolic syndromes, accounts for highest morbidity and mortality worldwide. Diabetes mellitus is characterized by abnormalities in carbohydrate, lipid and protein metabolism due to complete or relative insufficiency of insulin secretion from pancreatic β-cells and/or defect in insulin action (Unger and Foster, 1998). This syndrome affects approximately 5% of the world’s populations and almost 32 million people die due to diabetes across the world every year. It has been estimated that more than 18 million people in USA are affected by diabetes mellitus every year (Boyle et al., 2001). The current annual incidence rate (30 million) of diabetes mellitus in India will be expected to be double by the year 2025 due to it’s immense population (King et al., 1998). Alloxan induces diabetes mellitus in a wide variety of animal species by causing damage to pancreatic β-cells (Mallaisse, 1982). Alloxan induced diabetes is thus widely used as an experimental model to evaluate the antidiabetic effect of medicinal plants and their constituents.
Free radical is defined as an atom or molecule that contains one or more unpaired electrons, which are generally unstable and highly reactive (Mahdi, 2002). Free radicals can cause oxidative damage to DNA, lipids and proteins, which inturn result in the pathogenesis of several disorders including cancer and diabetes mellitus. The major targets of free radicals are membrane lipids, particularly polyunsaturated fatty acids (PUFA) where they initiate a sequential process known as lipidperoxidation (Slater, 1984). Peroxidation of membrane lipids may cause impairment of membrane function, reduce cell fluidity, inactivation of membrane-bound receptors and enzymes and increase permeability to ions and eventually membrane rupture.
Reactive Oxygen Species (ROS) play an important role in the pathogenesis and development of diabetic complications (Halliwell and Gutteridege, 1984). Oxidative stress has been implicated in the pathogenesis of tissue damage in Type I and Type II diabetes. Oxidative stress induced by alloxan is one of the proposed mechanisms for its diabetogenic action (Seghrouchni et al., 2002). Organism has however evolved sophisticated antioxidant defense system for protection against free radical mediated oxidative damages at different sites (Sies, 1993). The antioxidant system includes enzymatic (superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx)) and non-enzymatic antioxidant (reduced glutathione (GSH), Vitamin C and Vitamin E) (Mc Crod et al., 1971; Kirkman et al., 1987).
Several medicinal plants and herbs are still used by village and tribal populations of India and other developing countries for the treatment of various diseases including diabetes mellitus (Joshi, 2000). More than 150 medicinal plants are presently used in folk medicine for the treatment of diabetes mellitus in India. Piper longum, a well-known Indian medicinal spice, possesses diverse biological activities and pharmacological functions. It is widely used in Siddha, Ayurveda and Unani systems of medicine, particularly for diseases of the respiratory tract. It is considered beneficial in arthritis, peptic ulcers, viral hepatitis, fertility and diabetes. Piper longum can be used with adjuvants in anti-TB treatment and as hepatoprotective agent (Atal et al., 1985). Experimental studies have also shown their anticancer activity (Sunila and Kuttan, 2004; Selvendiran et al., 2003).
No scientific data regarding the antihyperglycemic and antilipidperoxidative effects of ethanolic extract of Piper longum dried fruits are however available except in the treatise of Ayurvedic and Siddha medicine. Thus, the present study was designed to evaluate and provide scientific evidence for the antihyperglycemic and antilipidperoxidative effects of Piper longum dried fruits in alloxan induced experimental diabetes mellitus.
MATERIALS AND METHODS
Animals: Albino Wistar male rats 7 to 8 weeks old and weighing 150-200 g were used for the present study. The animals were obtained from Central Animal House, Rajah Muthiah Institute of Health Sciences, Annamalai University, India and were maintained at 12 h light-dark cycles. The animals were randomized into experimental and control groups and were housed 4 or 5 in a polypropylene cage. Standard pellets obtained from Mysore Snack Feed Ltd, Mysore, India, were used as a basal diet during the experimental period. The control and experimental animals were provided food and drinking water ad libitum.
Chemicals: Alloxan was purchased from Sigma Aldrich Chemicals, Pvt., Ltd., Bangalore. All other chemicals and reagents used were of analytical grade.
Plant material: Piper longum dried fruits was purchased from traditional market in Chidambaram, Tamilnadu and identified by the Botanist, Dr. S. Sivakumar, Reader, Department of Botany, Annamalai University. A voucher specimen was also deposited in the Department of Botany, Annamalai University.
Preparation of plant extract
Ethanolic extract preparation: Five hundred gram of fresh Piper longum dried fruits was powdered and then soaked in 1500 mL of 95% of ethanol overnight. After filtration, the residue obtained was again resuspended in equal volume of 95% ethanol for 48 h and filtered again. The above two filtrates were mixed and the solvent was evaporated in a rotavapor at 40°50°C under reduced pressure. A 12% semisolid dark brown material obtained was stored at 0-4°C until used. A known volume of the ethanolic residual extract is suspended in distilled water and was orally administered to the animals by gastric intubation using a force-feeding needle during the experimental period.
Induction of diabetes mellitus: Diabetes mellitus was induced in overnight fasted (12 h) Wistar rats by single intraperitoneal injection of freshly prepared solution of alloxan monohydrate (150 mg kg-1 bw) in physiological saline (Al-Shamaony et al., 1994).
Experimental design: The local institutional animal ethics committee, Annamalai University, Annamalainagar, India approved the experimental design. A total number of 30 rats (18 diabetic rats, 12 control rats) were used and the rats were divided into 5 groups of six each.
Group I served as normal animals and received 2 mL of distilled water (instead of ethanolic extract) by gastric intubation using force-feeding needle. Group II animals were treated with single intraperitoneal injection of alloxan monohydrate (150 mg kg-1 bw) after overnight fasting for 12 h. Determining the blood glucose concentration 3 days and 5 days after alloxan treatment assessed the diabetic condition. The rats with blood glucose level above 260 mg dL-1 and urinary sugar (+++) were selected for the experimental study. Group III animals were received 2 mL of the water solution of the residual PLEFet (300 mg kg-1 bw) once daily, for 45 days after the diabetic state was assessed in alloxan induced diabetic rats. Group IV animals were received the reference drug glibenclamide (600 μg kg-1 bw) in 2 mL of distilled water once daily for 45 days after the diabetic state was assessed in alloxan induced diabetic rats. Group V animals were treated with PLEFet (300 mg kg-1 bw) alone for 45 days in order to evaluate the hypoglycemic effect of the PLEFet in control rats. After the experimental period, all animals were sacrificed by cervical dislocation and biochemical studies were conducted in blood, plasma, erythrocytes, erythrocyte membrane, liver and kidney of control and experimental animals in each group.
Biochemical estimations: Plasma was separated from heparinized blood by centrifugation at 3000 rpm for 15 min. After plasma separation, the buffy coat was removed and the packed cells were washed thrice with physiological saline. A known volume of erythrocytes was lysed with hypotonic phosphate buffer at pH 7.4. The hemolysate was separated by centrifugation at 10,000 rpm for 15 min at 20°C. The erythrocyte membrane was isolated according to the procedure of Dodge et al. (1968) with a change in buffer according to Quist (1980). Liver and kidney samples from animals were weighed and homogenized using appropriate buffer in an all glass homogenizer with Teflon pestle using specified medium and then used for biochemical estimations.
Blood glucose was determined by the method of Sasaki et al. (1972) using O-toluidine reagent. Total haemoglobin was measured by the method of Drabkin and Austin (1932) and glycosylated haemoglobin was determined by the method of Sudhakar and Pattabiraman (1981). Plasma insulin was estimated by ELISA method using Boehinger Mannheim GmbH Kit (Anderson et al., 1993). Liver glycogen content was measured by the method of Morales et al. (1973). The activities of hexokinase, glucose-6-phosphatase, glucose-6-phosphate dehydrogenase, fructose 1,6-bisphosphatase and glycogen phosphorylase were estimated according to the methods of Brandstrup et al. (1957), Koida and Oda (1959), Ellis and Kirkman, (1961), Gancedo and Gancedo (1971) and Shull et al. (1956), respectively.
TBARS in plasma, erythrocyte membranes, liver and kidney were assayed according to the methods of Yagi (1978) Donnan (1950) and Ohkawa et al. (1979), respectively. Lipidhyropperoxides in plasma, liver and kidney were assayed according to the method of Jiang et al. (1932). Non-enzymatic antioxidants, Vitamin-C and reduced glutathione were measured according to the methods of Omaye et al. (1979) and Beutler and Kelley (1963), respectively. The enzymatic antioxidants, superoxide dismutase, catalase and glutathione peroxidase activities were estimated according to the methods of Kakkar et al. (1984) Sinha (1972) and Rotruck et al. (1984), respectively.
Statistical analysis: The data are expressed as mean±SD. Statistical comparisons were performed by one way analysis of variance (ANOVA) followed by Duncan’s Multiple Range Test (DMRT). The results were considered statistically significant if the p-values were less than 0.05.
RESULTS
Blood glucose and glycosylated haemoglobin levels were significantly increased whereas plasma insulin, total haemoglobin and liver glycogen were decreased in diabetic animals (Group II) as compared to control animals (Group I). The urinary sugar was found to be (+++) in diabetic animals. The status of blood glucose, urinary sugar, glycosylated haemoglobin, total haemoglobin and plasma insulin and liver glycogen were restored in diabetic rats after treated with PLEFet (Group III) and diabetic rats Treated with glibenclamide (Group IV). PLEFet showed a comparable effect to that of glibenclamide. Wistar rats treated with PLEFet alone (Group V) showed no significant difference in blood picture and liver glycogen content as compared to control rats (Group I) (Table 1).
| Table 1: | Blood glucose, urine sugar, plasma insulin, total hemoglobin, glycosylated hemoglobin and liver glycogen concentrations in control and experimental animals in each group |
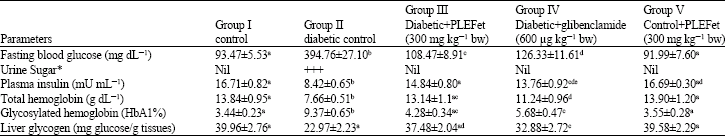 | |
| Values are given as mean±SD (n = 6 rats); Values not sharing a common superscript letter(s) differ significantly at p<0.05 (DMRT); PLEFet--Piper longum ethanolic dried fruits extract;-(+++) Indicates more than 1% sugar | |
| Table 2: | Activities of hexokinase, glucose-6-phosphatase, glucose-6-phosphate dehydrogenase, fructose 1,6 bisphosphatase and glycogen phosphorylase in liver and kidney of control and experimental animals in each group |
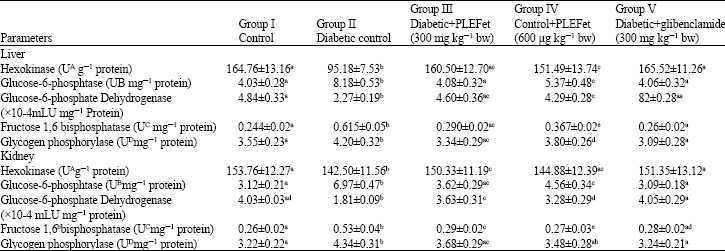 | |
| Values are given as mean±SD (n = 6 rats); Values not sharing a common superscript letter differ significantly at p<0.05 (DMRT); A-Micromoles of glucose phosphorylated/min; b-Micromoles of Pi liberated/min; C-Micromoles of Pi liberated/hour; D-Micrograms of Pi liberated/min. Piper longum ethanolic dried fruits extract | |
| Table 3: | Levels of TBARS and Lipid hydroperoxides in plasma, erythrocyte membrane and tissues of control and experimental animals in each group |
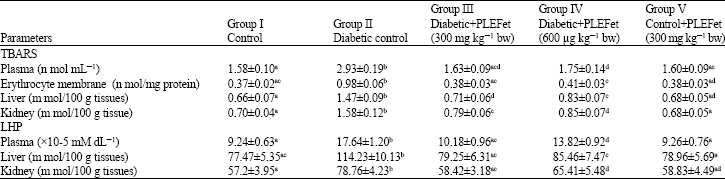 | |
| Values are given as mean±SD (n = 6 rats); Values not sharing a common superscript letter differ significantly at p<0.05 (DMRT); PLEFet-Piper longum ethanolic dried fruits extract | |
Glucose-6-phosphatase, glycogen phosphorylase and fructose-1, 6-bisphosphatase activities were significantly increased whereas the activities of hexokinase and glucose-6-phosphate dehydrogenase were decreased in diabetic rats as compared to control rats (Table 2). However the activities of glucose-6-phosphatase, hexokinase, fructose-1, 6-bisphosphatase, glucose-6-phosphate dehydrogenase and glycogen phosphorylase were returned to near normal range in diabetic rats treated with PLEFet and diabetic rats treated with glibenclamide. Wistar rats treated with PLEFet alone showed no significant difference in the activities of enzymes as compared to control rats.
TBARS and LHP levels were significantly increased in alloxan induced diabetic rats as compared to control rats. Treatment of alloxan induced diabetic rats with PLEFet for 45 days normalized the plasma and tissue TBARS and LHP.
PLEFet also showed comparable effect to that of glibenclamide. Wistar rats treated with the PLEFet alone showed no significant difference in TBARS and LHP levels as compared to control rats (Table 3).
CAT, SOD and GPx activities were significantly decreased in alloxan induced diabetic rats as compared to control rats. Treatment of alloxan induced diabetic rats with PLEFet for 45 days restored the activities of CAT, SOD and GPx. The effect of PLEFet was comparable to that of glibenclamide. Wistar rats treated with PLEFet alone showed no significant difference in enzymatic antioxidants activities as compared to control rats (Table 4).
The vitamin-C and glutathione levels were significantly decreased in plasma, liver and kidney diabetic rats as compared to control animals (Table 5). The levels of vitamin-C and glutathione were returned to near normal range in diabetic rats treated with PLEFet and diabetic rats treated with glibenclamide. The PLEFet showed also comparable effect to that of glibenclamide. Wistar rats treated with PLEFet alone showed no significant difference in vitamin-C and glutathione levels as compared to control rats.
| Table 4: | Activities of enzymatic antioxidants in plasma, erythrocytes, liver and kidney of control and experimental animals in each group |
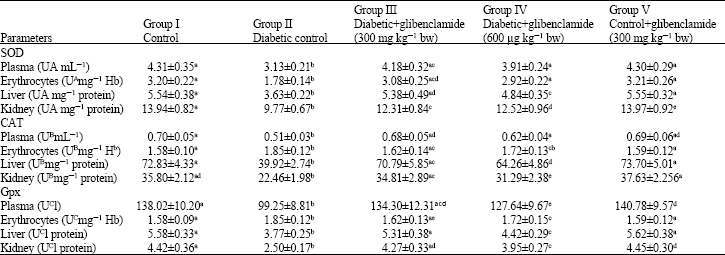 | |
| Values are given as mean±SD (n = 6 rats) Values not sharing a common superscript letter differ significantly at p<0.05 (DMRT); A-The amount of enzyme required to inhibit 50% NBT reduction; B-Micromoles of H 2O2 utilized/Sec. C-Micromoles of glutathione utilized/min; PLEFet-Piper longum ethanolic dried fruits extract ss | |
| Table 5: | Levels of non enzymatic antioxidants in plasma, erythrocyte membrane, liver and kidney of control and experimental animals in each group |
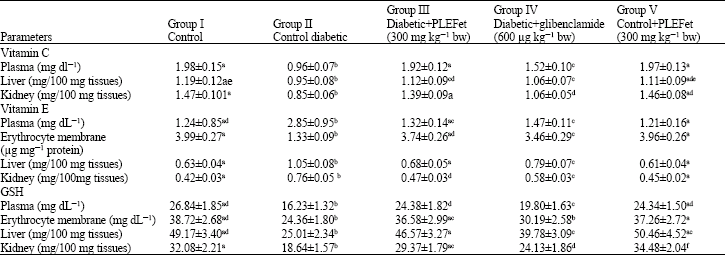 | |
| Values are given as mean±SD (n = 6 rats) Values not sharing a common superscript letter differ significantly at p<0.05 (DMRT); PLEFet-Piper longum ethanolic dried fruits extract | |
DISCUSSION
In the present study, we have investigated the antihyperglycemic and antilipidperoxidative effects of ethanolic extract of dried fruits of Piper longum (PLEFet) in alloxan induced diabetic rats. Increase in blood glucose was accompanied by an increase in glycosylated Hb and decrease in plasma insulin, liver glycogen content and altered activities of carbohydrate metabolizing enzymes in diabetic rats as compared to control rats. Elevated TBARS and decline in antioxidants status in blood and tissues were also noticed in alloxan induced diabetic rats. Oral administration of PLEFet at a dose of 300mg kg-1 bw to diabetic rats for 45 days restored the above said biochemical variables.
Alloxan induces diabetes mellitus in a wide variety of animal species by causing damage to pancreatic β-cells which in turn result in poor utilization of glucose by hepatic and extrahepatic tissues of diabetic rats due to impairment in insulin secretion and action (Malaisse et al., 1982). Oral administration of PLEFet for 45 days restored the blood glucose in diabetic rats, which indicates that the extract stimulated the activity of the liver to maintain the normal homeostasis of blood glucose during diabetes. This was also evident by increase in liver glycogen content and plasma insulin level after 45 days in diabetic rats treated with PLEFet (Ceriello, 2000). The antidiabetic effect of PLEFet suggests that they stimulated insulin secretion from surviving pancreatic β-cells.
The observed increase in glycosylated Hb and decrease in total Hb in diabetic rats indicates poor glycemic control mechanism (Koenig et al., 1976). The increase in total hemoglobin and decrease in glycosylated hemoglobin in diabetic rats treated with PLEFet indicates its ability to prevent the glycosylation process and there by improved the glycemic control mechanisms.
Two major hormones, insulin and glucagon maintain glucose homeostasis. Decrease in the enzymatic activity of hexokinase and increase in the activity of glycogen phosphorylase observed in the present study are responsible for the depletion of liver glycogen. The altered activities of hexokinase, glucose-6-phosphatase, glucose-6-phosphate dehydrogenase, fructose-1, 6-bisphosphatase and glycogen phosphorylase enzymes in diabetic rats are probably due to insulin insufficiency or defect in insulin action (Clement et al., 1996). The observed decrease in glucose-6-phosphatase, glycogen phosphorylase and fructose-1, 6-bisphosphatase activities and increase in glycogen content, glucose-6-phosphate dehydrogenase and hexokinase activities in diabetic rats treated with PLEFet suggest that the PLEFet have the capacity to correct the alterations of carbohydrate metabolizing pathway in diabetes mellitus, as evident by increased plasma insulin and liver glycogen content.
Increase in plasma insulin and decrease in blood glucose levels observed in diabetic rats treated with PLEFet also suggest its ability to correct the metabolic alterations in the activities of carbohydrate metabolizing enzymes. Elevated TBARS and decline in antioxidants status was observed in diabetic rats. Determination of plasma TBARS and lipidperoxides is considered as the most reliable marker to assess the extent of tissue damage in pathological conditions. Increased plasma TBARS and lipidperoxides observed in diabetic rats could be due to over production and diffusion of lipidperoxidation by products from damaged pancreas, liver, kidney and erythrocytes with subsequent leakage in to plasma (Ravi et al., 2004). Oral administration of PLEFet restored the level of TBARS in diabetic rats, which indicates its antilipidperoxidative (scavenging excess ROS) potential during diabetes.
Vitamin C and reduced glutathione, a well-known antioxidants scavenges a wide variety of reactive oxygen species and protect cells from oxidative damage. Lowered levels of GSH and vitamin C are probably due to their utilization by tissues to combat the deleterious effects of ROS mediated oxidative damage.
SOD protects tissues by catalyzing the removal of superoxide radical that damage the membrane structure and function (Grankvist et al., 1979). Catalase is responsible for the detoxification of significant amounts of H2O2. Reduced activities of SOD and CAT observed during diabetes therefore cause number of deleterious effects to cells and tissues due to the accumulation of potent superoxide radical and hydrogen peroxide. Lowered activities of these enzymes were reported by several authors in diabetic conditions (Punitha and Manoharan, 2006; Punitha et al., 2006). Glutathione peroxidase converts H2O2 to H2O at the expense of reduced glutathione (Sakurai and Ogiso, 1991). Lowered activity of GPx in diabetes mellitus is therefore due to low availability of GSH (Tannaka et al., 2002). Prsent results corroborate these observations. Enhancement in SOD, CAT and GPx activities and the levels of GSH and vitamin C in diabetic rats treated with PLEFet clearly shown that it has free radical scavenging principles that alleviated the pathological effects of reactive oxygen species.
Our results thus clearly demonstrated that the ethanolic extract of dried fruits of Piper longum (PLEFet) has potent antihyperglycemic and antilipidperoxidative effects in alloxan induced diabetic rats. Further studies are warranted to isolate and characterize the bioactive antidiabetic principles from the dried fruits of Piper longum, which can therefore be used as an alternative remedy for the treatment of diabetes mellitus and oxidative stress associated diabetic complications.
REFERENCES
- Al-Shamaony, L., S.M. Al-Khazraji and H.A.A. Twaiji, 1994. Hypoglycaemic effect of Artemisia herba alba. II. Effect of a valuable extract on some blood parameters in diabetic animals. J. Ethnopharmacol., 43: 167-171.
CrossRefPubMedDirect Link - Anderson, L., B. Dinesen, P.N. Jorgesen, F. Poulsen and M.F. Roder, 1993. Enzyme immunoassay for intact human insulin in serum or plasma. Clin. Chim. Acta, 39: 578-582.
PubMedDirect Link - Atal, C.K., R.K. Dubey and J. Singh, 1985. Biochemical basis of enhanced drug bioavailability by piperine: Evidence that piperine is a potent inhibitor of drug metabolism. J. Pharmacol. Exp. Ther., 232: 258-262.
PubMedDirect Link - Boyle, J.P., A.A. Honrycult, K.M. Narayan, T.J. Hoerger, L.S. Geiss, H. Chen and T.J. Thompson, 2001. Projection of diabetes burden through 2050. Impact of changing demography and disease prevalence in the US. Diabetes Care, 24: 1936-1940.
CrossRefDirect Link - Brandstrup, N., J.E. Kirk and C. Bruni, 1957. The hexokinase and phosphoglucoisomerase activities of aortic and pulmonary artery tissue in individuals of various ages. J. Gerentol., 12: 166-171.
PubMedDirect Link - Donnan, S.K., 1950. The thiobarbituric acid test applied to tissues from rats treated in various ways. J. Biol. Chem., 1982: 415-419.
Direct Link - Drabkin, D.L. and J.M. Austin, 1932. Spectrophotometric studies: I. Spectrophotometric constants for common hemoglobin derivatives in human, dog and rabbit blood. J. Biol. Chem., 98: 719-733.
Direct Link - Grankvist, K., S. Marklund, J. Sehlin and I.B. Taljedal, 1979. Superoxide dismutase, catalase and scavengers of hydroxyl radical protect against the toxic action of alloxan on pancreatic islet cells in vitro. Biochem. J., 182: 17-25.
PubMed - Halliwell, B. and J.M.C. Gutteridge, 1984. Lipid peroxidation, oxygen radicals, cell damage and antioxidant therapy. Lancet, 323: 1396-1397.
CrossRefDirect Link - Kakkar, P., B. Das and P.N. Viswanathan, 1984. A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem. Biophys., 21: 130-132.
PubMedDirect Link - King, H., R.E. Aubert and W.H. Herman, 1998. Global burden of diabetes, 1995-2025: Prevalence, numerical estimates and projections. Diabetes Care, 21: 1414-1431.
CrossRefPubMedDirect Link - Koenig, R.J., C.M. Peterson, R.L. Jones, C. Saudek, M. Lehrman and A. Cerami, 1976. Correlation of glucose regulation and hemoglobin A1C in diabetes mellitus. New Eng. J. Med., 295: 417-420.
CrossRefDirect Link - Malaisse, W.J., 1982. Alloxan toxicity to the pancreatic β-cell: A new hypothesis. Biochem. Pharmacol., 31: 3527-3534.
CrossRefDirect Link - McCord, J.M., B.B. Keele Jr. and I. Fridovich, 1971. An enzyme-based theory of obligate anaerobiosis: the physiological function of superoxide dismutase. Proc. Natl. Acad. Sci. USA., 68: 1024-1027.
PubMedDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Omaye, S.T., J.D. Turnbull and H.E. Sauberlich, 1979. Selected method for the determination of ascorbic acid in animal cells, tissues and fluids. Methods Enzymol., 62: 3-11.
PubMed - Punitha, R. and S. Manoharan, 2006. Antihyperglycemic and antilipidperoxidative effects of Pongamia pinnata (Linn.) Pierre flowers in alloxan induced diabetic rats. J. Ethnopharmacol., 105: 39-46.
Direct Link - Punitha, R., K. Vasudevan and S. Manoharan, 2006. Effects of Pongamia pinnata flowers on blood glucose and oxidative stress in alloxan induced diabetic rats. Ind. J. Pharmacol., 38: 62-63.
Direct Link - Quist, E.H., 1980. Regulation of erythrocyte membrane shape by Ca2+. Biochem. Biophys. Res. Commun., 92: 631-637.
CrossRefPubMedDirect Link - Ravi, K., B. Ramachandran and S. Subramanian, 2004. Protective effect of Eugenia jambolana seed kernel on tissue antioxidants in streptozotocin-induced diabetic rats. Biol. Pharm. Bull., 27: 1212-1217.
CrossRefDirect Link - Rotruck, J.T., A.L. Pope, H.E. Ganther, A.B. Swanson, D.G. Hafeman and W.G. Hoekstra, 1973. Selenium: Biochemical role as a component of glutathione peroxidase. Science, 179: 588-590.
CrossRefPubMedDirect Link - Sakurai, K. and T. Ogiso, 1991. Inhibitory effect of glutathione on the generation of hydroxyl radicals in the reaction system of glutathione-alloxan. Chem. Pharm. Bull., 39: 737-742.
Direct Link - Sasaki, T., S. Matsy and A. Sonae, 1972. Effect of acetic acid concentration on the colour reaction in the O-toludine boric acid method for blood glucose estimation. Rinsh Kagaku, 1: 346-353.
Direct Link - Seghrouchni, I., J. Drai, E. Bannier, J. Riviere, P. Clamard, I. Garcia, J. Orgiazzi and A. Revolm, 2002. Oxidative stress parameters in type I, type II and insulin treated type 2 diabetes mellitus; insulin treatment efficiency. Clin. Chim. Acta, 321: 89-96.
CrossRef - Selvendiran, K., J. Prince Vijeya Singh, K. Baba Krishnan and D. Sakthisekaran, 2003. Cytoprotectivity effect of piperine against benzo[a]pyrene induced lung cancer with reference to lipidperoxidation and antioxidant system in Swiss albino mice. Fitoterapia, 74: 109-115.
Direct Link - Shull, K.H., J. Ashmore and J. Mayer, 1956. Hexokinase, glucose-6-phosphatase and phosphorylase levels in hereditarily obese-hyperglycemic mice. Arch. Biochem. Biophys., 62: 210-216.
CrossRefDirect Link - Sinha, A.K., 1972. Colorimetric assay of catalase. Anal. Biochem., 47: 389-394.
CrossRefPubMedDirect Link - Slater, T.F., 1984. Free-radical mechanisms in tissue injury. J. Biochem., 222: 1-15.
CrossRefPubMedDirect Link - Nayak, S.S. and T.N. Pattabiraman, 1981. A new colorimetric method for the estimation of glycosylated hemoglobin. Clinica Chimica Acta, 109: 267-274.
CrossRefPubMedDirect Link - Sunila, E.S. and G. Kuttan, 2004. Immunomodulatory and antitumor activity of Piper longum Linn. and piperine. J. Ethnopharmacol., 90: 339-346.
CrossRefDirect Link - Tannaka, Y., P.O. Tran, J. Harmon and R.P. Robertson, 2002. A role for glutathione peroxidase in protecting pancreatic beta cells against oxidative stress in a model of glucose toxicity. Proc. Natl. Acad. Sci. USA., 99: 12363-12368.
Direct Link - Yagi, K., 1987. Lipid peroxides and human diseases. Chem. Physiol. Lipids, 45: 337-351.
CrossRefDirect Link