
Karina Manayeva
Graduate School of Dairy Sciences, Rakuno Gakuen University, Ebetsu, Japan
Buho Hoshino
Department of Environmental Symbiotic, College of Agriculture, Food and Environment Sciences, Rakuno Gakuen University, Ebetsu, Japan
LiveDNA: 81.14171
Hiromasa Igota
Department of Environmental Symbiotic, College of Agriculture, Food and Environment Sciences, Rakuno Gakuen University, Ebetsu, Japan
Takashi Nakazawa
Graduate School of Dairy Sciences, Rakuno Gakuen University, Ebetsu, Japan
Ganzorig Sumiya
Department of Environmental Symbiotic, College of Agriculture, Food and Environment Sciences, Rakuno Gakuen University, Ebetsu, Japan
International Journal of Zoological Research
Year: 2017 | Volume: 13 | Issue: 1 | Page No.: 26-37







ABSTRACT
Background: Tibetan antelope (Pantholops hodgsonii), an endemic species of the Tibetan Plateau listed on IUCN Red List, migrates over the long distances for calving. It has an unequal feature that only females do seasonal migration. However, the problems of seasonal migration and inhabitation of Tibetan antelope have not yet been clarified. Methodology: To identify the ecological drivers of the Tibetan antelope migration in terms of geographical characteristics, topographical variables of their home ranges and the factors affecting the habitat selection within the seasons, there are utilized the Argos transmitter satellite tracking of the Tibetan antelope. The analysis was made by using the LoCoH Hull method for the home range estimation made for this species for the first time. Results: Tibetan antelope has spent more time on the way back from calving, than the way forward. The annual habitation area varies from 2024-2908 km2 with average of 2409 km2 and the wintering place locates in a valley with the average HR of 441 km2. Conclusion: The results of monitoring and LoCoH Hull Home Range (HR) analysis showed that the calving ground location continues to be stable, while wintering places changes easily, one-third of samples changed its wintering pastures and could be adjacent to different groups. This is an important results that leads to the protection and management of endangered species on the Red List like Tibetan antelope.
PDF Abstract XML References Citation
Received: October 26, 2016;
Accepted: November 27, 2016;
Published: December 15, 2016
How to cite this article
Karina Manayeva, Buho Hoshino, Hiromasa Igota, Takashi Nakazawa and Ganzorig Sumiya, 2017. Seasonal Migration and Home Ranges of Tibetan Antelopes (Pantholops hodgsonii) Based on Satellite Tracking. International Journal of Zoological Research, 13: 26-37.
DOI: 10.3923/ijzr.2017.26.37
URL: https://scialert.net/abstract/?doi=ijzr.2017.26.37
DOI: 10.3923/ijzr.2017.26.37
URL: https://scialert.net/abstract/?doi=ijzr.2017.26.37
INTRODUCTION
Tibetan antelope, commonly called "Chiru" (Pantholops hodgsonii) is a large bovine species (Bovidae, Artiodactyla), endemic to the Tibetan Plateau. This species inhabits high-elevation alpine and desert steppe with flat to uneven terrain in the Tibetan Plateau and only recently has been studied a little more in detail. Due to their highly valued wool the antelopes were heavily hunted in Tibet and the population once had sharply declined to nearly extinct in the past several decades1-3. Since early 1990, hunting on Tibetan antelopes have been prohibited and several reserves to protect their habitats were established in China. Although the population of chiru now is one of the best-protected wildlife in China, the antelope is listed as endangered species because of exploitation and competition with domestic livestock of pastoralists, the current populations estimated to around 100,000 animals4. Long-distance seasonal migrations of large herds of ungulates are widely knows they are driven by needs, such as food, water, avoiding predation and harsh climatic conditions5,6. Some, at least 5 populations of the Tibetan antelope are migratory, moving up to 300-400 km, others are non-migratory1. The Qinghai-Tibet railway, which started in July, 2006, cuts across the range of the Tibetan antelope population and may disturb its migratory. The infrastructure development including railway construction in the Tibetan Plateau was found to be the main factor that threatens this species now and in the future7.
Because of its distribution in highly elevated and harsh environment of the Tibetan Plateau, the biology and ecology of this unique species remain poorly understood. Despite the fact that seasonal migration of chiru and some of the calving places (Huiten or Zhuonai lake) were known as since 19th century8, many details of such behavior are still unknown. Recent studies in China revealed the presence of several calving places and implicated that female antelopes may travel up to 250 km from winter pastures to a summer calving sites1,9-13.
The aim of this study is to clarify the seasonal migration pattern and home ranges of the Tibetan antelope using the satellite tracking data derived from Argos transmitter collars. Such survey was done for the first time on the Tibetan antelope. High-resolution presence data collected from transmitters and in a training area were combined with environmental factors to create the models of species occurrence, which will be extrapolated to a projection area. It is expected to use the methods for the quantification of area of occupancy for highly endangered mammals in one of the most remote and barely studied regions of the world.
MATERIALS AND METHODS
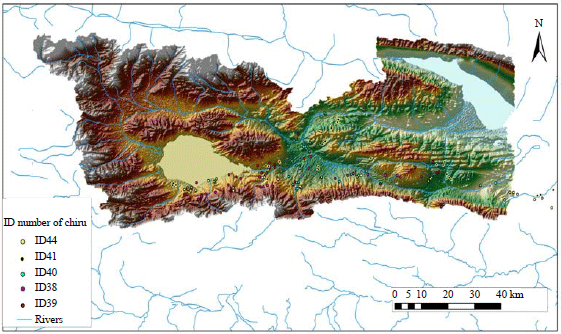
Study sites and satellite tracking data: The Hoh Xil nature reserve area is an isolated region in the Northwestern part of the Tibetan Plateau in China (Fig. 1). It is Chinese least and the world's third-least populated area. The region covers 83,000 km² at an average elevation of 4800 m, between the Tanggula and Kunlun mountain chains in the border areas of Southwest Chinese Tibet Autonomous region, Northwest China's Qinghai province and the Xinjiang Uyghur Autonomous region. It is one of the major headwater sources of the Yangtze river and Yellow river. Around 45000 km², at an average elevation of 4600 m was made into a Hoh Xil national nature reserve in 1995. The Qinghai-Tibet railway runs along the Eastern boundary of the reserve.
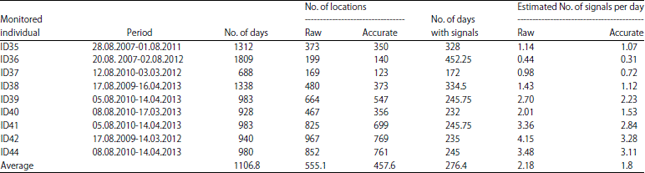
In this study, there are captured 9 female Tibetan antelopes near the Wu-Bei underpass (N35°15'; E93°09', altitude 4597 m) of Qinghai-Tibet railway using nets, provide by common morphometric measurements and collared than with satellite-based Argos Platform Transmitter Terminals (PTT) (model ST-20 A-3210, Telonics Inc., USA) in August, 2007 (2 females), August, 2009 (2 females) and August, 2010 (5 females). The duty cycles of the collars were settled on: 4 h on/92 h off and 6 h on/90 h off and the average frequency of received signals appeared to be 2.18 and 1.8 for accurate locations and each transmitting process (Table 1). Only relatively accurate locations were used for analysis and those data that fit into a known migration corridor7,12 were correspondent to the maximum ground travel limits of the animals were included. Erroneous locations that necessitated an unrealistically high speed of travel were removed14.
Data analysis: The satellite tracking of the migrating of Tibetan antelopes was successful, previous analyses of 2 tracked individuals, till 2009, showed the exact locations of summer (calving), intermediate and winter pastures, temporal distribution patterns, association of summer habitat selection with conditions of pastures and so on. Analysis of distances, durations and other specifics of migration of 9 Tibetan antelopes were made on the base of satellite information in according to physical parameters of the animals and habit. To utilize Argos PTT tracking data, in this study, there are used software ArcGIS10.1, ENVI 5.3, Surfer 11 software, ASTER Global Digital Elevation Model (GDEM, a product of METI and NASA), three-dimensional terrain data, water resources parameters, field census, habitat mapping and local convex Hull (LoCoH) Home Range (HR) models were used. Using the satellite image analysis software ENVI 5.3, there are calculated tilt angle (Slope), shadow (Shaded Relief) and slope orientation (Aspect) were calculated.
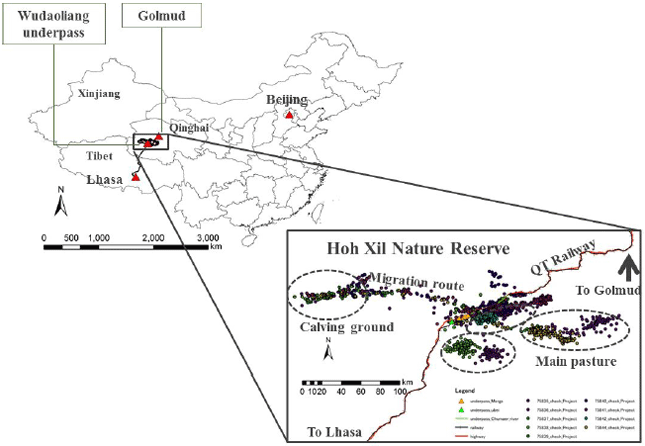 | |
| Fig. 1: | Map of study site and habitat of Tibetan antelope. Red and black line shows Qinghai-Tibet highway and railway, points shows Argos tracking ground signals of Tibetan antelope |
| Table 1: | Monitoring periods and amount of locations for collared individuals of Tibetan antelope based on satellite tracking Argos data |
 | |
A LoCoH non-parametric kernel method was applied to construct the animal HR models. This method generalizes the Minimum Convex Polygon (MCP) method and was shown to be more appropriate than parametric kernel methods for constructing HRs and Utilization Distributions (UDs), because of its ability to identify hard boundaries (e.g., rivers and cliff edges) and convergence to the true distribution as sample size increases15,16. The software for all three methods of local convex Hull home range estimation (k, r and a-LoCoH) is available at http://locoh.cnr.berkeley.edu. The k-LoCoH home range with isopleth’s values (No. of nearest neighbors) 10-15-20-25 was calculated for each individual separating migration from wintering area.
RESULTS
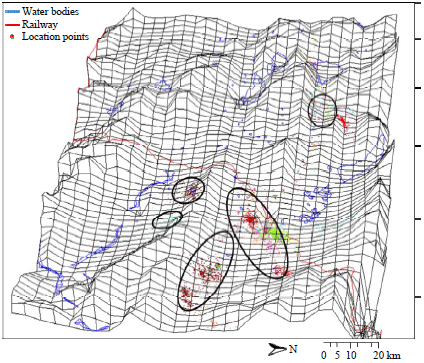
Satellite tracking of behaviour and migration patterns of Tibetan antelope: The analysis of obtained data shows that 7 of the 9 observed females migrated in the similar way to each other following a latitudinal gradient and shared the same area (such as surrounding of Huiten Nur lake area) for calving. However, there two individuals did not migrate (Fig. 2).
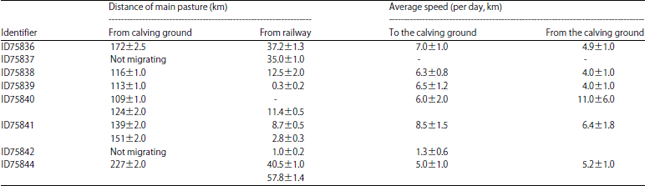
Two of the 7 migrating individuals (ID36 and ID38) used the Southward habitation area. When the migration began, they crossed a hill through a valley (altitude 4700 m) before joining the main migration corridor. The ID37, which didn’t migrate, shared almost the same area with ID44 in winter, which located in a distance of 35-40 km Eastward from the railway, the core area of the second non-migrating, ID42 is near the railway, 1.2 km from Wu-Bei underpass on the Southern part of main pastures of ID35 (2008 and 2011), ID39, ID40 and ID41. The ID44 used two areas for pasture: One used in autumn and spring, which located farther (to the East) and the other used in wintertime. The distance between two pastures is around 12 km. The ID41 also used a different habitation area for autumn and spring of which altitude is higher than winter pasture. This area was located Northeastward from the place used in winter. The distance between two places is 25 km. The ID41 spent some time between two zones in late autumn and early spring. Antelope ID41 come near the calving ground on winter 2012-2013 and have not come back to the previous wintering pasture area. The core area of ID35 was located near the highway in winter 2007-2008 and 2010-2011, in winter seasons 2008-2009 and 2009-2010 antelope changed its core area and inhabits more to the East, the distance between two different areas is around 62 km. Table 2 shows the distances of wintering HRs from the railway and the highway. Hence, 1:3 of samples changes its wintering pastures irregularly and could adjacent to different groups. The reasons of ID35, ID41 and ID44 instability are considered to be the changes in environment and adaptation, especially to the railway construction and surrounding area dynamics. The evidences are also seen in the dates of migration, its shifts and delays near the railway and the highway (Table 3).
The features of migration parameters are shown in the Fig. 3. Main habitation areas were commonly used for 8-9 months from mid-August to mid-May. The average period of migration is 83±5 days with 24±3 days spent on the calving ground, 22±4 days on the migration route to the calving place and 31±8 days on the migration route from the calving place, including 12±10 days spent near the highway, delaying migration and looking for a proper time and place to cross it. All antelopes crossed under particular bridges in a specific way, according to the highway traffic load, river banks circulations, number of Tibetan antelope groups (lines), especially after the calving season during the migration. That the timing of the giving birth is very important. The migration started from the beginning of June, 2008 and 2009, in late-May, 2010, approximately in the middle of May, 2011 and in early-May, 2012. The terminations of migration were in the end of August, 2008 and 2009, in the middle of August, 2010 and in the beginning of August, 2011 and 2012. Hence, the period of migration shifted to the earlier dates from 2010 for for about 2.5 weeks.
 | |
| Fig. 2: | Positions of wintering places of 9 tracked Tibetan antelopes and locations during the migrations and calving (PTTs locations, Aster DEM by ENVI©) |
| Table 2: | Statistical results of migration parameters of individuals of Tibetan antelope based on satellite tracking Argos data |
 | |
| Table 3: | Statistical results of migration periods (days) of individuals of Tibetan antelope based on satellite tracking Argos data |
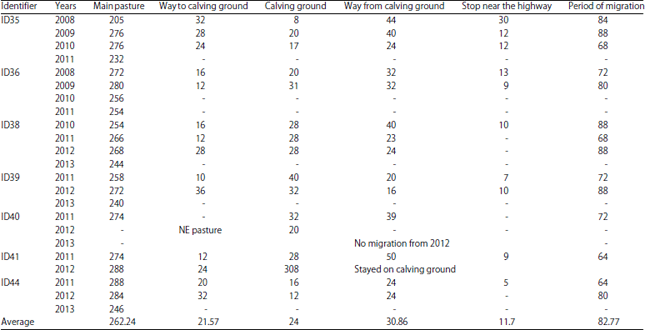 | |
The ID44 began migration at 4-8th June, 2011. This individual (and ID36) spent more time on main pastures than others, in spite of relatively long distance from calving place and the average speed of migration was lower but the antelope returned from calving to core area in a short period. Other individuals used more time for returning and average speeds were lower in a case of delaying on intermediate areas of a migration route. Tibetan antelopes ID38 and ID39 spent more time in a calving ground. All investigated animals had lowly averages of moving speeds on the way back to main pastures, when young followed the females. On the way back from the calving ground the Tibetan antelope with collar ID35 spent from 21-35 days in 2008 moving along the highway upward before crossing. The ID36 also spent around 13 days in 2008 before crossing the highway and railway and before reaching the wintering area. All migrating Tibetan antelopes delayed their migration for 5-12 days because of staying near the highway. The ID41 started migration from calving ground on 14th July, 2011 and appeared near the highway on 22nd July (93.84 km), stayed there until 30th July and reached the main pasture on 3rd August. Next year, in 2012, this antelope initiated migration earlier, on about 17th May (comparing with approximate 31st May, 2011) and stayed near the calving ground for wintering. The ID40 moved to NNE from calving ground on 3rd July, 2011 and stayed on alternative pasture from 10th July until the 30th July, 2011. On 3rd August it appeared near the highway and reached the main, wintering pasture on 11th August.
 | |
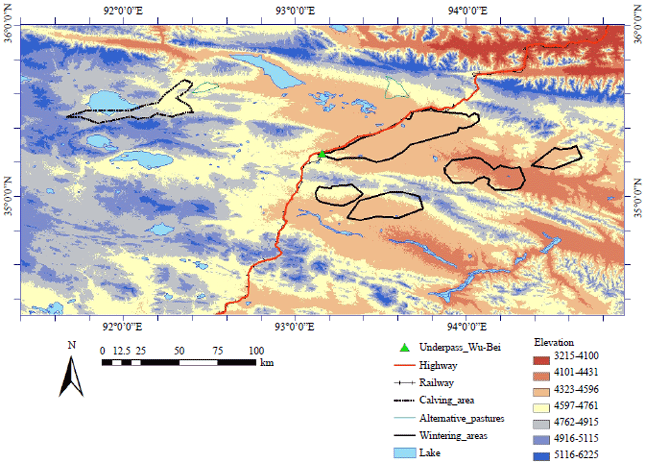
| Fig. 3: | Migration routes and a calving ground with locations of studied Tibetan antelopes at Huiten Nur lake, Hoh Xil national reserve, Qinghai, China. Details of slope, aspect and shaded relief from the satellite based on the digital topographic model |
 | |
| Fig. 4: | Home range polygons and range of the calving ground |
The topography map (Fig. 4) shows the features of the habitation area of the studied Tibetan antelopes (elevation), the valley of wintering pastures, the migration route and a position of calving ground. The detailed relief and the proximity of rivers of the narrow area of migration route near the calving ground at the Huiten Nur (Chinese name is Zhuonai-Hu) lake was shown on the calving ground map. Also, the layout of core areas and calving area location of all monitored Tibetan antelopes are seen on the topographic model and the hull of migration route of ID44 which almost corresponds to other samples, represents the model of corridor of migration route, made by the LoCoH home range method.
LoCoH Hull home range models: The actual sizes and locations of main habitation home ranges, migration and calving areas of each individual have been calculated (Fig. 5, 6). According to the home range model, the annual habitation area including migration and calving ground of all 9 monitored antelopes varies from 2024-4111 km2, with an average of 2652 km2. Therefore, the annual habitation areas vary in 2 times. The area will vary from 2024-2908 km2 with an average of 2409 km2 if to exclude the habitation area of ID35 is excluded, which changed its main pastures.
The wintering place was located in a valley, with an elevation about 4500±100 m. The calving place was highly elevated, it rises further 400-500 m and locates near the Huiten Nur lake on a flat area and it is surrounded by hills. The migration corridor lied on a rising valley with average slopes of 15-20 degrees and had a narrow area between the hills, closer to the calving ground.
The average home range of wintering places was 441 km2, with minimal of 246.39 and maximal of 644.07 km2, the area of ID35 had been excluded because it consists of two core areas and has a size of 1939.32 km2. The isopleths area of the home range of ID35, seen on the Fig. 5a has higher value of two different core used areas.
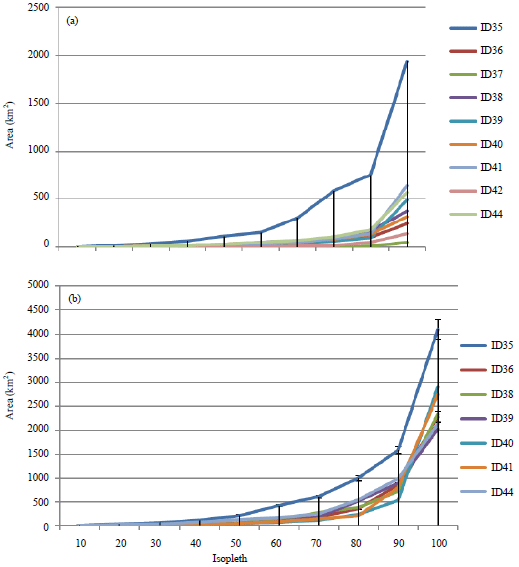 | |
| Fig. 5(a-b): | Areas of (a) Wintering HRs and (b) Annual HRs isopleths of studied Tibetan antelopes (Hull LoCoH) |
In addition, the areas of ID37 and ID42 were excluded, due to that the both animals did not migrate and the areas are smaller than the others migrating ones: 46.65 and 138.01 km2. The most intensively used areas, showed on maps by more intensive grey colors characterizes of the Chiru’s potential inhabitants locations appearing near the riverbanks in valley on elevation 4300-4600 m. The value of isopleths from 10-50 represents the most intensive used area and could be named as a core area, it has a small diameter, the value of isopleths from 60-100 shows less intensively used area.
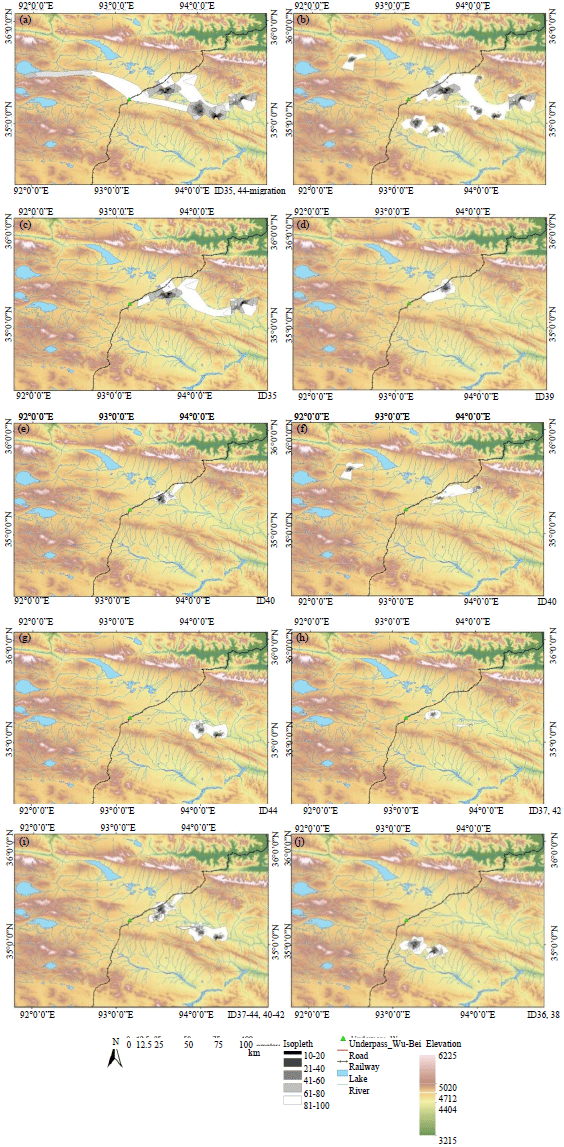 | |
| Fig. 6(a-j): | Hull LoCoH home ranges of wintering pastures of the Tibetan antelope. (a) Annual home ranges of ID44 (migration included) and ID35, (b) Home ranges of all studied individuals, (c-g) Home ranges of each individual, inhabits near the Qinghai-Tibet railway and highway (d, e, f), in valley (g) and in both places (c), (h) Non-migrating individuals, (i) Home ranges of non-migrating individuals locates in the similar area with migrating individuals and (j) Home ranges of individuals, inhabits beyond the hill |
| Table 4: | Percentages of frequently used area isopleths of wintering area home ranges (Hull LoCoH) |
 | |
| Table 5: | Percentages of isopleths of wintering home ranges from annual home ranges (Hull LoCoH) |
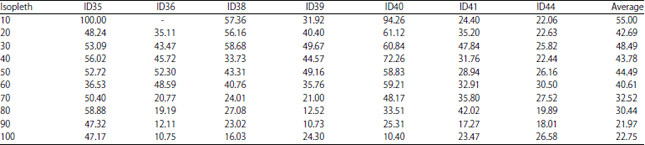 | |
Percentages of value 50, 70, 80 isopleths from the value 100 isopleths of wintering areas are shown in (Table 4).
The percentages of wintering HRs isopleths from the annual ones and the sizes of isopleths are shown in Table 5. Wintering HR isopleths are from 21.97-55% in average from annual HR (which includes calving ground and migration route) with more percentages of low value (10-50) isopleths. The HR locations are divided into 3 groups: The first locates near the highway and railway, second locates in the valley 35-40 km Eastward from the railway and the third is beyond the hill in another valley. Also some individuals changed their main habitation areas within the wintering, 9 months period and in different years. Other classification is made by migration: Migrating or non-migrating individuals. Therefore, the caught samples appear to be representational and illustrate the different types of HRs.
The shapes of home ranges are commonly round or oval with most frequently used areas in the middle, tend to Southward. Except the home ranges of ID35 and ID41 that changed their core areas in different years, its HRs have S-shape. In addition, the HR of ID44 is divided into 2 core areas, because this representative changed the main pasture within the seasons (autumn-spring and winter).
The HRs locates near the artificial barriers-railway and a highway had the parallelepiped shape, which clearly showed the dependence of the distribution on the limiting barriers. Also, the valley, surrounded by hills and the close located of rivers with most frequently used areas are seen on the LoCoH HR maps of Fig. 6a-j. The individuals, inhabited at the other valley, beyond the hill use that valley for their migration. However, the accession to the primary migration route is necessary in case of sharing one mutual calving ground. The local people questionnaire data tells that previously Tibetan antelopes inhabited in Southern valley followed another route and united to the main migration stream farther to the Northwest. However, the railway construction changed their route making them getting together before crossing the railway-highway in more convenient place (e.g., Wu-Bei under path).
DISCUSSION
Some of observed Tibetan antelopes used pastures with different conditions, such as altitude, in autumn and winter, some of them can change the main habitation area and then after 2 years, return to the previous place. Numerous factors could be stimuli for the beginning of migration, such as the changes of day length, precipitation, pregnancy term or physiological mechanism of motherhood. Until now, there were no significant data to support the hypotheses of the stimulus for the beginning of migration. The reasons of migration include the dependence on earth surface characteristics, climate conditions, stable fresh water resource, escape from predators, protection of newborns, probable seasonal changes in diet, etc. "Nutritious way" may play an important role (the high vegetation value on the migration route when females are on gestation and lactation). Argos telemetry monitoring provides a year-round and remote data, the knowledge of the species in spatial and temporal scale.
Indirect division into classes leads to separate analysis of the migrating and non-migrating individuals (ID37 and ID42). Individuals which came from different valleys and had to cross the hill (ID36 and ID38) showed the importance of one mutual migration corridor, as well as similar timing of migration to the mutual calving ground. The direction of migration corresponds to the common topography. Females move to the calving ground towards the valley. The number of calving grounds are limited17,10, therefore Tibetan antelopes inhabiting in the neighboring valleys had to cross the natural barriers such as hills and rivers to form large herds and migrated in groups by the same migration route to the calving area. The calving ground location continues to be stable, while wintering places change easily. Understanding of details of female Tibetan antelopes migration into particular sites on definite time to high altitude severe area, the sizes and positions of home ranges, the behavioral features and preferences have an important implication in Tibetan antelope general biology and conservation.
The migration discontinuance may be caused by barrenness of females, health conditions and so on. In the study, there is estimate evidence of negative influences of the highway and the railway constructions in interaction on female and offspring seasonal migration for breeding, such as migration of 1 individual with 2 years of observation and its stay near the calving ground in winter 2012-2013 and long stops near the highway and the railway. The combination of railway and highway in combination has an effect on delays of the migration (up to 23±8 days), causing stress and becoming a serious obstacle for migrating females. Animals stay next to the linear barriers before crossing and the dates of migration have been shifted to earlier dates. However, the degree of that influence remains unclear according to the available information, it is concluded that the adaptation is satisfied and even though animals tolerate the inconveniences and has to adjust and shift being under the continuous stress.
Tibetan antelopes spent more time on the way back from the calving ground. Females with calves concentrate next to the highway and form herds of several thousands (observed during the field survey). Such bunching and later fragmented crossing of the obstacles by small groups may cause the reshuffle of primary families in parcels and companions and change in hierarchy of herds, which constitute mainly by kinship. Further survey of Tibetan antelope herds’ hierarchy is required for estimation of its influence on reproduction, grazing, movement and so on. In addition, the larger sizes of herds cause the less pronounced behavioral aspects of formation and dynamics of herds’ units. Due to exposure of anthropogenic factors complex on the Tibetan antelope and its habitat, the spatial arrangement of population (ways, terms and intensity of migration and areas of wintering and calving) is changed and fragmented. It leads to intrapopulational and interpopulational ties violations, food competition growth and to irrational usage of forage resources (overgrazing in some places and underutilization in another). All these must be considered in the development of measures to preserve the population of such stenobiontic and endemic species as the Tibetan antelope.
The increase of highway and railway traffic complicates its intersection and normal course of ungulates migration. It is naturally expected that traffic volume will increase significantly in the future. To prevent the migration delays and the concentration of big-sized herds and mixing of herds, it is necessary to provide the faster and safer crossing of artificial barriers. Therefore, the construction of special transitions for migrating wildlife, which will ease the transportation ways crossings, should be provided. The optimum construction of such transition for cautious and timid ungulates of open spaces (flatlands and plateaus) is considered be the laying of road sections in shallow underground tunnel, slightly recessed metro-like, in wildlife migration routes intersections. Wherein the ground surface of transport tunnel occurrence will look like untransformed. The construction has to be considered with the relative locations of frequently used by Tibetan antelope passes under the railway. To estimate a definite frequently used railway underpasses, for advising the location and structures of the crossing construction, further survey is required.
The population is under the threat from habitat loss and gaps in ecological knowledge. A better understanding of the drivers of Tibetan antelope migration at multiple scales is a key step towards addressing these threats. As far as the region economic development increases (including tourism), it requires strong conservation strategy of species and arrangement of its less stressful migration way. The preservation of Tibetan antelope habitat, including migration routes by minimizing of anthropogenic influences has a significant role for survival of this endemic species.
CONCLUSION
For the first time ever, the Argos tracking has been made on the Tibetan antelope and the received location data provided of the detailed information about spatial and temporal patterns of the movements of this rare endemic animal. The results of monitoring and LoCoH Hull Home Range (HR) analysis showed that the calving ground location continues to be stable, while wintering places changes easily, one-third of samples changed its wintering pastures and could be adjacent to different groups. The annual habitation area varies from 2024-2908 km² with average of 2409 km² and the wintering place locates in a valley with the average HR of 441 km². The most intensively used areas characterizes of its locations appearing near the riverbanks in valley on elevation 4300-4600 m. The HRs classifications were made by migration existence (Two of studied Tibetan antelopes are non-migratory) and stability of HRs within the seasons and years. One studied individual stayed on a calving ground on the last year of our survey. The HRs, located next to the artificial barriers (railway and a highway) had the parallelepiped shape, which shows its dependence on those factors. The width of the migration corridor was wider in neighboring to linear barriers area, evidencing that the artificial constructions influenced on the migration behavior.
SIGNIFICANCE STATEMENT
For the first time ever, the Argos tracking has been made on the Tibetan antelope and the received location data provided of the detailed information about spatial and temporal patterns of the movements of this rare endemic animal. The results of monitoring and home range analysis showed that the calving ground location continues to be stable, while wintering places changes easily, one-third of samples changed its wintering pastures and could be adjacent to different groups. This is an important results that leads to the protection and management of endangered species on the Red List like Tibetan antelope.
ACKNOWLEDGMENTS
This study was supported by Grant-in-Aid for Scientific Research (A) (No. 19255005, Project leader is: Prof. Buho Hoshino and core member of project including Prof. N. Ohtaishi, Prof. K. Kaji, Prof. M. Kaneko, Dr. T. Yoshida, Dr. M. Motokawa, Prof. R. Masuda) by the Japan Society for the Promotion of Science (JSPS). The authors would like to thank the State Forestry Administration of China (Beijing) for providing us with the permission to perform a field work. We have succeeded under the full cooperation of Dr. Z. Jiang, Prof. Liu, Chuguang, Prof. Wu, Xiaomin, Dr. Mahmut, Halik, Mr. T. Nakazawa on the installation the satellite transmitter Argos and participated field work and captured and collared animals by the Argos satellite tracking system. We also thank Mr. Bian Kun, Mr. Hongfeng Zhang, Mr. Tuo Feng and all our research colleagues who helped during the field survey on behalf of Shaanxi Institute of Zoology (Northwest Institute of Endangered Zoological Species), Xi-An, China.
REFERENCES
- Brantingham, P.J., J.W. Olsen and G.B. Schaller, 2001. Lithic assemblages from the Chang Tang region, Northern Tibet. Antiquity, 75: 319-327.
CrossRefDirect Link - Leslie, Jr. D.M. and G.B. Schaller, 2008. Pantholops hodgsonii (Artiodactyla: Bovidae). Mammalian Species, 817: 1-13.
CrossRefDirect Link - Ruan, X.D., P.J. He, J.L. Zhang, Q.H. Wan and S.G. Fang, 2005. Evolutionary history and current population relationships of the chiru (Pantholops hodgsonii) inferred from mtDNA variation. J. Mammal., 86: 881-886.
Direct Link - Singh, N.J., I.A. Grachev, A.B. Bekenov and E.J. Milner-Gulland, 2010. Saiga antelope calving site selection is increasingly driven by human disturbance. Biol. Conserv., 143: 1770-1779.
CrossRefDirect Link - Ito, T.Y., B. Lhagvasuren, A. Tsunekawa, M. Shinoda, S. Takatsuki, B. Buuveibaatar and B. Chimeddorj, 2013. Fragmentation of the habitat of wild ungulates by anthropogenic barriers in Mongolia. PLoS One, Vol. 8.
CrossRef - Xia, L., Q. Yang, Z. Li, Y. Wu and Z. Feng, 2007. The effect of the Qinghai-Tibet railway on the migration of Tibetan antelope Pantholops hodgsonii in Hoh-Xil National Nature Reserve, China. Oryx, 41: 352-357.
CrossRef - Schaller, G.B. and R. Junrang, 1988. Effects of a snowstorm on Tibetan antelope. J. Mammal., 69: 631-634.
CrossRefDirect Link - Schaller, G.B., A. Kang, X. Cai and Y. Liu, 2005. Migratory and calving behavior of Tibetan antelope population. Acta Theriol. Sin., 26: 105-113.
PubMedDirect Link - Bleisch, W.V., P.J. Buzzard, H. Zhang, D. Xu, Z. Liu, W. Li and H. Wong, 2009. Surveys at a Tibetan antelope Pantholops hodgsonii calving ground adjacent to the Arjinshan nature reserve, Xinjiang, China: Decline and recovery of a population. Oryx, 43: 191-196.
CrossRefDirect Link - Hoshino, B., Z. Jiang, C. Liu, T. Yoshida and M. Halik et al., 2011. Preliminary study on migration pattern of the Tibetan antelope (Pantholops hodgsonii) based on satellite tracking. Adv. Space Res., 48: 43-48.
CrossRefDirect Link - Hays, G.C., S. ArKesson, B.J. Godley, P. Luschi and P. Santidrian, 2001. The implications of location accuracy for the interpretation of satellite-tracking data. Anim. Behav., 61: 1035-1040.
CrossRefDirect Link - Getz, W.M., S. Fortmann-Roe, P.C. Cross, A.J. Lyons, S.J. Ryan and C.C. Wilmers, 2007. LoCoH: Nonparameteric kernel methods for constructing home ranges and utilization distributions. PloS One, Vol. 2.
CrossRef - Guisan, A. and N.E. Zimmermann, 2000. Predictive habitat distribution models in ecology. Ecol. Model., 135: 147-186.
CrossRefDirect Link