
I.V. Utama, A.B.I. Perdamaian
Laboratory of Genetics and Breeding, Faculty of Biology, Universitas Gadjah Mada, Yogyakarta, Indonesia
B.S. Daryono
Laboratory of Genetics and Breeding, Faculty of Biology, Universitas Gadjah Mada, Yogyakarta, Indonesia
International Journal of Poultry Science
Year: 2018 | Volume: 17 | Issue: 10 | Page No.: 486-492







ABSTRACT
Background: The native Indonesian chicken, the Pelung, is widely known as a preferred source of meat and eggs but it has the disadvantage of relatively slow growth. Recently, ♂Pelung chickens were crossed with ♀ F1 chickens (♀ Pelung×♂ Broiler chicken) to generate the first back-crossed (BC1) chicken for use as a new breed. Molecular markers were observed within the Growth hormone (Gh) gene. Avian Gh (AY461843) is one of the most important genes influencing growth-related traits; specifically, the Gh intron 3 Single Nucleotide Polymorphism (SNP) G+1705A is significantly associated with the growth traits of Chinese chicken breeds. This study aimed to produce a new, fast-growing chicken line that is more homozygous for morphological traits than its parents and to observe the Gh gene intron 3 SNP G+1705A polymorphism in this BC1 chicken. Methodology: Day-old chickens (DOCs) were intensively reared for 7 weeks and weighed weekly. Qualitative and quantitative characteristics and the Feed Conversion Ratio (FCR) of the BC1 chickens were determined at the seventh week of observation. Restriction Fragment Length Polymorphism (RFLP) analysis was performed to detect genotype differences among the BC1 populations. Results: In both sexes, weight gain was faster in the BC1 chickens (919.9 grams) than in the Pelung chickens (434.74 g) but lower than in the broiler (1.500 g) chickens. The FCR for the BC1 line (2.32) was between that of the Pelung (3.35) and broiler (1.55) chickens. The BC1 hybrid chicken genotype was 89% dominant homozygous (GG) and 11% heterozygous (GA). Conclusion: In this study, the body parameters did not significantly differ among the genotypes (GA and GG). The BC1 chicken was deemed satisfactory for use as a meat chicken.
PDF Abstract XML References Citation
Received: February 28, 2018;
Accepted: August 21, 2018;
Published: September 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
I.V. Utama, A.B.I. Perdamaian and B.S. Daryono, 2018. Plumage Uniformity, Growth Rate and Growth Hormone Polymorphism in Indonesian Hybrid Chickens. International Journal of Poultry Science, 17: 486-492.
DOI: 10.3923/ijps.2018.486.492
URL: https://scialert.net/abstract/?doi=ijps.2018.486.492
DOI: 10.3923/ijps.2018.486.492
URL: https://scialert.net/abstract/?doi=ijps.2018.486.492
INTRODUCTION
Numerous researchers have genetically improved Indonesian native chickens and have successfully created several local meat breeds. Recently, Pelung chickens, the heaviest local chickens, were successfully mated with broilers to derive a fast-growing line, despite a lack of uniformity in morphological traits1-3.
The Pelung was originally desired by chicken fanciers as an ornamental breed but scientific reports have shown better growth-related trait performance when compared to other local breeds. Seven-week-old Pelung can reach 500 g: adult males weigh approximately 5.5 kg, while females weigh approximately 3.5 kg. Interestingly, crossing Pelung with another local breed will produce hybrid chickens that weigh over 750 g, which is greater than both parents at ten weeks old. Heterosis underlies this excellent hybrid chicken growth performance. In terms of egg productivity, female Pelung naturally start laying at seven months old and yield 39-70 eggs each year.
In this study, female F1 chickens (♀Pelung×♂ Broiler chicken) were back-cross mated with their fathers (male Pelung chickens) to produce a new chicken line that is more homozygous for morphological traits and has a better body weight than that of the parents. Feed optimization to match the new metabolism of this hybrid chicken was studied to understand the genetic improvement effort4 and to determine whether further enhancements are needed.
Plumage and shank color as well as the final weight uniformity of the flock are important. If at least 80% similarity is reached, a chicken line can be released to market. A uniform chicken growth rate ensures no delay in harvest time and the combination of plumage and shank color is an important identifying character.
In chickens, genetic selection for growth can be performed more precisely and more accurately using molecular markers and the genes underlying chicken growth may be candidate molecular markers. One of the most influential genes determining chicken growth is chicken growth hormone (Gh). In the chicken genome, Gh (AY461843) is located on 1q45 and is 4,101 bp long, consisting of 5 introns and 4 exons. Single Nucleotide Polymorphism (SNP) on intron number 3 (G1705A) was reported to be significantly associated with growth6.
Growth Hormone (GH) is produced by somatotrophs (acidophilic cells), which are generally found in the anterior pituitary7. In birds, GH has a single amino acid chain that is released from secretory granules into the bloodstream by the stimulation of hypothalamic releasing factors8. Circulating GH increases cellular metabolism and induces cells to enter the mitotic phase.
Genomic Gh sequences and the expression product (i.e., protein) have been intensively studied in commercial chicken lines. Contrary to the Gh polymorphisms in their commercial counterparts, these polymorphisms in local chickens and their association with growth rates are unknown. This research aimed to identify Gh polymorphisms and their association with the growth rate in BC1 chicken lines.
Restriction Fragment Length Polymorphism (RFLP) is currently used to detect polymorphisms in specific regions of the chicken genome. This method utilizes the EcoRV restriction enzyme to recognize GATATC sequences and digestion is applied to detect disparities among progenies.
In this study, the growth rate, shank and plumage color uniformity and growth hormone gene polymorphism of first back-crossed chickens were observed.
MATERIALS AND METHODS
Resource populations and chicken rearing: Three female F1 chickens (♀Pelung×♂ Broiler chicken) were crossed with one male Pelung and semi-intensively maintained from October to December 2015 and the laboratory works was finished at late 2016. All parents were given free access to feed and water and housed in 7×5×2-m pens. After artificial incubation, 28 day-old chickens (DOCs) were produced that consisted of 15 females and 13 males. All DOCs were maintained following broiler welfare standards for 7 weeks and weighed weekly. The Feed Conversion Ratio (FCR) was calculated from the final chicken body weight and total consumed feed data using the following formula9:
Where
| fw | = | Final weight |
| iw | = | Initial weight |
| fi | = | Feed intake |
Qualitative and quantitative characters: Both qualitative character (comb type, plumage and shank color) and quantitative character (body height, body width, body length, beak length, head length, head width, comb height, comb length, chest diameter, back length, wing length, neck length and femur length) assessments were performed during the 7th week of observation.
Amplification and population genotyping: Blood samples were collected from the wing axillary vein using a syringe and subsequently stored in vacutainers filled with EDTA before preservation in a refrigerator. Prior to using the salting-out method10 with modification for DNA isolation, 50 μL of the nucleated chicken blood samples were thawed and diluted with Phosphate Buffer Saline (PBE) to 250 μL. 900 μL Low salt buffer (1.21% Tris HCl [10 mM] pH 7.6, 0.75% KCl [10 mM], 2% MgCl2, 0.7% EDTA [2 mM]) and 50 μL Triton-X detergent was added to the mixtures and followed by incubation at 37°C for 5 min. The mixture was then centrifuged at 12.000 rcf for 3 min at room temperature. The supernatant was removed and the pellet was retained for further processing. The pellet was resuspended in 300 μL High salt buffer (2.5% Tris HCl [10 mM] pH 7.6, 0.14% KCl [10 mM], 2.4% MgCl2, 0.14% EDTA [2 mM], 0.9% NaCl [0.4 M]) and 40 μL 10% SDS solution prior to second incubation at 37°C for 5 min. After being incubated, 100 μL 6 M NaCl was added to the mixture and vortexed. The mixture was then centrifuged at 12.000 rcf for 5 min at room temperature. The supernatant was placed into new microtube for further processing. Approximately 440 μL isopropanol (equal to the mixture volumes) was added then inversely mix prior to centrifuge at 12.000 rcf for 10 min. The pellet was resuspended in 100 μL 70% ethanol before 12.000 rcf for 5 min. Discard the supernatant and repeated the resuspend step for three times before air dried. The dry DNA pellet was eluted in 50 μL TE buffer.
The quality of DNA samples was achieved by agarose gel electrophoresis (MUPID-exU, Japan) and quantified by spectrophotometry (Shimadzu, Japan). The genomic DNA amplification procedure followed the manufacturer’s protocol (Kapa Biosystems, United States) using a gradient thermal cycler (Bio-Rad, United States). Twenty five μL of PCR cocktail was freshly prepared by mixing 12.5 μL of Kapa fast-start ready mix, 1.25 μL of forward primer, 1.25 μL of reverse primer, 8 μL of ddH2O and 2 μL of DNA sample, which was then vortexed for 5 seconds followed by spin-down at 2000 rpm. All primers were 10×diluted with Tris-EDTA (TE) prior to use. The machine was set to initial denaturation at 95°C for 5 min before 35 cycles of denaturation at 95°C for 30 sec each. The sample was then annealed at 60°C for 20 sec and extended at 72°C for 30 sec followed by post extensions for 5 min at 72°C. The primers used in this study refer to the previous study11: F: 5-TCCCAGGCTGCGTTTTGTTCTC (forward) and 5'-ACGGGGGTGAGCCAGGACTG-3' (reverse). After band integrity and purity were verified by agarose gel electrophoresis, 10 μL of the amplicon of each experimental chicken was mixed with the restriction enzyme EcoRV (8U), 5 μL of buffer and 34 μL of ddH2O, which was then centrifuged. These mixtures were incubated for 16 h at 37°C to digest the desired DNA sequence following previous methods12. Digestion products were subjected to 2% agarose gel electrophoresis prior to being photographed on a UV transilluminator.
RESULTS AND DISCUSSION
There are numerous chicken breeds in Indonesia that have resulted from both conventional and traditional breeding techniques. The Pelung breed is one of the traditional breeding products (Fig. 1) used as an ornamental chicken. However, the Pelung has potential for use as a meat chicken due to its heavy body weight.
Chicken growth is determined by Quantitative Trait Loci (QTL) that are influenced by internal (genetics, sex) and external (rearing management and nutrition) factors. The genetic factors underlying chicken growth concurrently consist of an SNP, DNA methylation and other epigenetic factors within the genome that contribute to males gaining more weight than females10. Maintaining the optimum chicken environment with respect to temperature, humidity, flock density and feed enhances the growth rate because little energy must be allocated to maintaining homeostasis. Furthermore, chicken feed requirements increase with age because of decreasing intestinal nutrition-ingestion efficiency.
Body weight was measured until the seventh week of age because the chicken growth rate increased up to this point and then declined13. Figure 2 shows the sexual dimorphism observed in BC1 males (n = 13) and females (n = 15) compared with that in the Pelung chicken.
Based on Fig. 2, the average body weights of male and female BC1 chickens were 919.9±105.72 and 852.5±82.98 g, respectively, while the weights of the male and female Pelung chickens reached only 434.74±47.15 and 404.57±42.53 g in the seventh week of observation.
 | |
| Fig. 1: | The morphological characteristics of male Pelung chickens. The typical wild type shank (yellow and black) and plumage (black and red) color, heavy body weight and crowing duration indicate breed purity |
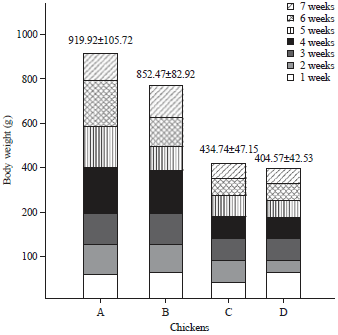 | |
| Fig. 2: | Body weight comparison (grams) of male (A) and female (B) first back-crossed (BC1) chickens to male (C) and female (D) pelung chickens at 7 weeks of age |
| Table 1: | Qualitative characters of first back-crossed (BC1) chickens at seven weeks of age |
 | |
The disparity between the average body weights of BC1 chickens and those of Pelung chickens was consistent with our hypothesis that weight gain was more rapid in BC1 than in Pelung for both sexes. These large differences might be due to the accumulation of growth-related genes inherited from the broiler ancestors.
Based on the current research, the FCR of BC1 chickens (2.32) was higher than that of the broilers (1.55) but less than that of the Pelung (3.35) chickens14 despite an intensive rearing program. Therefore, the BC1 FCR was between that of the Pelung and broiler chickens but relatively closer to that of the broiler chicken. This suggests that the BC1 chickens in the present study have good prospects for becoming meat chickens because their FCR value was better than that of the Pelung chicken, although further improvement is needed to reach the broiler FCR value.
In terms of qualitative traits (Table 1), divergence was observed in plumage colors but uniformity was observed in comb type and shank color. The chicken population was dominated by white-colored plumage (male: 46% and female: 47%) followed by brownish (male: 31% and female: 26%), brown (male: 23% and female: 20%) and black (male: 0% and female: 7%). The genetic dose effect of multiallelic plumage traits emerged in the plumage colors of many BC1 chickens after segregation from the wild-type plumage (e+) of male heterozygote and brown (eb) heterozygote female F1 Pelung parents. The single-comb type (rprp) and black shank (ZidW; ZidZid) were inherited from both parents, which possessed similar characters.
These results (Table 2) show that BC1 chickens were quantitatively more similar to broiler chickens than to Pelung chickens. Of the 15 characteristics observed in BC1 chickens, seven (chicken height, body height, comb height, comb length, body width, wing length and femur length) were not significantly different from those of broilers, while the other characteristics differed significantly.
Compared to the characteristics of Pelung, five characteristics (comb height, comb length, body length, body width and chest circumference) of BC1 chickens were not significantly different but 10 other characteristics differed significantly.
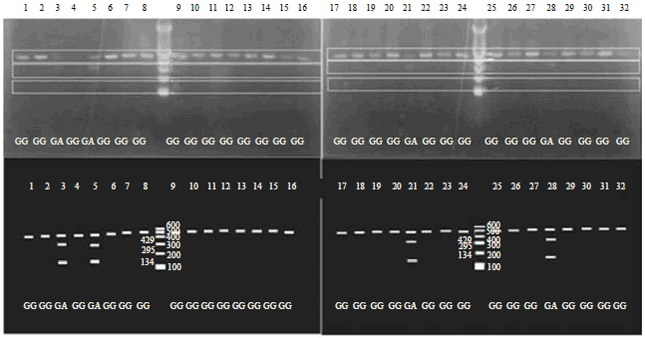 | |
| Fig. 3: | Genotypes of Pelung, F1 and BC1chickensbased on Gh intron 3 polymorphism |
| Pelung: 1: First filial (F1): 2-4 and first back-crossed (BC1) chicken: 5-32. Twenty-eight chickens were GG and four were GA | |
| Table 2: | Comparison of quantitative characters among first back-crossed (BC1), broiler and Pelung chickens at seven weeks of age |
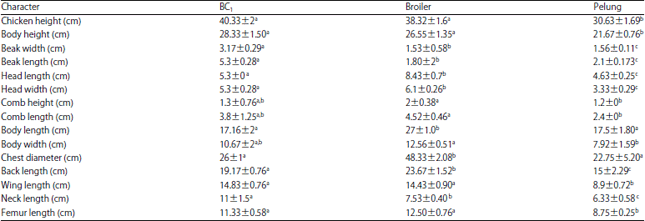 | |
From the 15 characteristics compared quantitatively, the value similarity of BC1 chickens with broiler chickens was 47% (7 characteristics), while the value similarity with Pelung chickens was 33% (5 characteristics).
Based on our study, salting out DNA extracting method10 with modification was success to yield sufficient DNA starting materials for molecular works. The genomic DNA concentration and purity was proved by gel electrophoresis (no smear was detected) and spectrophotometry (260:280 ratio was around 1.8; concentration above 40 ng mL–1). Compared to the available commercial DNA extraction kit, our method cost less and can be performed at low sophisticated laboratory but no excess reagent was compromised neither the Taq polymerase nor the restriction enzyme works.
Based on our results (Fig. 3), seven-eighths of the chickens were homozygous dominant (GG) for Gh intron 3. Those DNA fragments, which were uncut by restriction enzyme EcoRV, were 429 bp long. Four subjects were heterozygous (GA) for Gh, which had three fragments (429, 295 and 134 bp long). Based on gender, all male BC1 chickens had the homozygous-dominant (GG) genotype and 3 of 15 female BC1 chickens had the heterozygous genotype (GA) while the rest were homozygous dominant (GG). The Pelung chickens had the GG genotype, while F1 chickens had the GG (1 chicken) and GA (2 chickens) genotypes.
These results might have been due to the more frequent emergence of the GG and GA genotypes than the AA genotype, where G is the dominant allele and A is the recessive allele11. As previously reported, the AA genotype has a positive effect on the growth rate of chickens when compared with the GA and GG genotypes. Polymorphisms in the Gh intron 3 (G+1705A) of BC1 chickens were similar to those reported to be significantly associated with growth traits in previous studies6. A previous report6 found that noncoding SNPs, such as those in the region 5 ‘UTR, 3' UTR and introns, might affect gene expression levels because the noncoding regions were usually regulatory elements. In this study, mutations were identified as the purine base transition from Guanine (G) to adenine (A). The nucleotide sequence polymorphisms in intron 3 affect the regulation of Gh expression. However, mutations in the third intron might not alter gene expression but would affect the amount of GH secretion15. According to previous research6, genomic Gh expression is influenced by four SNPs in the 5 ‘UTR, one SNP in the3’ UTR, five SNPs in the exon and 36 SNPs in the intron.
In this study, body parameters did not differ significantly among genotypes (GA and GG). The potential for similar body parameters comes was observed in other SNP studies, with SNP G+1705A not being the sole genetic factor contributing to more desirable characteristics. The chickens used in this research were genetically different from the F2 population used in a previous study6 and thus might have slightly different molecular controls. Moreover, the genotypes of the heavy-weight BC1 chickens and the light-weight Pelung breed were similar, justifying our hypothesis. Further analyses will be required to determine the other molecular controls that determine BC1 body parameters.
A total of 89% (25 chickens) of the BC1 chickens had the dominant homozygous genotype (GG), 11% (3) had the heterozygous genotype (GA) and 0% had the homozygous recessive genotype (AA). Theoretically, according to Mendelian inheritance, BC1 chickens should have three genotypes: homozygous dominant (GG), heterozygous (GA) and homozygous recessive (AA).
Chicken GH is a peptide hormone produced by somatotropins (acidophilic cells) located in the anterior pituitary gland. Stimulated by growth hormone-releasing hormone (GHRH), GH is released into the blood vessels throughout the body and affects growth by stimulating cell metabolism and division8. Ultimately, the chicken growth rate was not only influenced by GH but also by other biological substances, such as IGFs PIT-1 and Ins, at all omic levels. Based on our research, the BC1 chickens were satisfactory as local meat chickens.
SIGNIFICANCE STATEMENT
In this study, selective breeding was used to obtain genetically improved local chickens and the most efficient mating scheme to retrieve a meat chicken line was discovered. Growth hormone polymorphism would be an applicable molecular marker for other local chickens in Indonesia.
ACKNOWLEDGMENTS
The authors would like to acknowledge the Ministry of Research and Higher Education of Indonesia (Ristekdikti) for a research grant (Insentif Riset Sinas No. 584/LPPM/2015) and the Innovation Center for Agro-Technology (Pusat Inovasi Agro Teknologi; PIAT) of UGM for providing the breeding facilities.
REFERENCES
- Saragih, H.T.S.S.G. and B.S. Daryono, 2012. Effect of high-protein diet on body weight and pectoralis thoracicus muscle performance on pelung and broiler chicken (Gallus gallus domesticus). Anim. Prod., 14: 199-204.
Direct Link - Retnoaji, B., R. Wulandari, L. Nurhidayat and B.S. Daryono, 2016. Osteogenesis study of hybrids of Indonesia’s native chicken pelung (Gallus gallus domesticus) with broiler (Gallus gallus domesticus). Asian J. Anim. Vet. Adv., 11: 498-504.
CrossRefDirect Link - Saragih, H.T.S.S.G., I. Roosdianto and B.S. Daryono, 2017. Pectoralis thoracicus muscle performance of hybrid chicken (F1) derived from crossbreed between broiler and pelung (Gallus gallus gallus). J. Kedokteran Hewan, 11: 62-64.
Direct Link - Perdamaian, A.B.I., H.T.S.S.G. Saragih and B.S. Daryono, 2017. Effect of varying level of crude protein and energy on insulin-like growth factor-i expression level in Indonesian hybrid chicken. Int. J. Poult. Sci., 16: 1-5.
CrossRefDirect Link - Tanaka, M., Y. Hosokawa, M. Watahiki and K. Nakashima, 1992. Structure of the chicken growth hormone-encoding gene and its promoter region. Genetics, 112: 235-239.
CrossRefPubMedDirect Link - Nie, Q., B. Sun, D. Zhang, C. Luo and N.A. Ishag et al., 2005. High diversity of the chicken growth hormone gene and effects on growth and carcass traits. J. Heredity, 96: 698-703.
CrossRefDirect Link - Yan, B., X. Deng, J. Fei, X. Hu, C. Wu and N. Li, 2003. Single nucleotide polymorphism analysis in chicken growth hormone gene and its associations with growth and carcass traits. Chin. Sci. Bull., 48: 1561-1564.
CrossRefDirect Link - Scanes, C.G. and S.J. Bowen, 1984. The role of growth hormones in the domestic fowl. Proceedings of the 33rd Annual National Breeder's Roundtable, May 3, 1984, Henry VIII Inn & Lodge, St. Louis, Missouri, pp: 42-69.
Direct Link - Zuidhof, M.J., B.L. Schneider, V.L. Carney, D.R. Korver and F.E. Robinson, 2014. Growth, efficiency, and yield of commercial broilers from 1957, 1978, and 2005. Poult. Sci., 93: 2970-2982.
CrossRefDirect Link - Suguna, S., D.H. Nandal, S. Kamble, A. Bharatha and R. Kunkulol, 2014. Genomic DNA isolation from human whole blood samples by non enzymatic salting out method. Int. J. Pharm. Pharm. Sci., 6: 198-199.
Direct Link - Anh, N.T.L., S. Kunhareang and M. Duangjinda, 2015. Association of chicken growth hormones and insulin-like growth factor gene polymorphisms with growth performance and carcass traits in Thai broilers. Asian-Aust. J. Anim. Sci., 28: 1686-1695.
CrossRefDirect Link - Maniatis, G., N. Demiris, A. Kranis, G. Banos and A. Kominakis, 2013. Genetic analysis of sexual dimorphism of body weight in broilers. J. Applied Genet., 54: 61-70.
CrossRefDirect Link - North, M.O. and D.D. Bell, 1990. Commercial Chicken Production Manual. 4th Edn., Van Nostrand Reinhold, New York, USA.
Direct Link - Zhang, X.L., X. Jiang, Y.P. Liu, H.R. Du and Q. Zhu, 2007. Identification of AvaI polymorphisms in the third intron of GH gene and their associations with abdominal fat in chickens. Poult. Sci., 86: 1079-1083.
CrossRefDirect Link