
A. B.I. Perdamaian
Laboratory of Genetics and Breeding, Faculty of Biology, Universitas Gadjah Mada, Jalan Teknika Selatan Sekip Utara, 55281 Yogyakarta, Indonesia
H.T. S.S.G. Saragih
Laboratory of Animal Structure and Development,
Faculty of Biology, Universitas Gadjah Mada, Jalan Teknika Selatan Sekip Utara, 55281 Yogyakarta, Indonesia
B. S. Daryono
Laboratory of Genetics and Breeding, Faculty of Biology, Universitas Gadjah Mada, Jalan Teknika Selatan Sekip Utara, 55281 Yogyakarta, Indonesia
International Journal of Poultry Science
Year: 2017 | Volume: 16 | Issue: 1 | Page No.: 1-5







ABSTRACT
Objectives: The aims of this study were to investigate the effect of crude protein and energy level in feed on insulin-like growth factor-I (IGF-1) expression level in a hybrid chicken (female Layer chicken×male Pelung chicken). Methodology: To study the correlation between feed type and IGF-1 expression level. Day-old chick (DOC) hybrids were divided into four groups and each group comprised 10 individuals. First group was fed with feed A [22% Crude Protein (CP) and 3100 kcal kg–1 Metabolizable Energy (ME)], second group was fed with feed B (20% CP and 3300 kcal kg–1 ME), third group was fed with feed C (21% CP and 2800 kcal kg–1 ME) and fourth group was fed with standard feed after starving for 3 days post hatching. Chickens were observed from post-hatch to the 7th day and weighed (at 0, 3 and 7 days) and five individuals were sacrificed at the 7th day for IGF-1 analysis. Total RNA was isolated from the liver for gene expression analysis. Real-time PCR was used to study the IGF-1 expression profiles of each group after being normalized with β-actin. Results: The highest body weight was recorded for group 1, followed by group 2, 3 and 4. The IGF-1 expression level was noticeably high for feed A, B and C groups by 4.82, 3.82 and 1.9 fold, respectively, compared with the starved group. The weight highly correlated with IGF-1 expression level (R2 = 0.8). This moderate growth response to nutrition makes the hybrid chicken a suitable meat-type chicken. Conclusion: The result suggested that feed containing 22% CP and 3100 kcal kg–1 ME is a promising starter for the Indonesian hybrid chicken.
PDF Abstract XML References Citation
Received: October 05, 2016;
Accepted: November 18, 2016;
Published: December 15, 2016
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
A. B.I. Perdamaian, H.T. S.S.G. Saragih and B. S. Daryono, 2017. Effect of Varying Level of Crude Protein and Energy on Insulin-like
Growth Factor-I Expression Level in Indonesian Hybrid Chicken. International Journal of Poultry Science, 16: 1-5.
DOI: 10.3923/ijps.2017.1.5
URL: https://scialert.net/abstract/?doi=ijps.2017.1.5
DOI: 10.3923/ijps.2017.1.5
URL: https://scialert.net/abstract/?doi=ijps.2017.1.5
INTRODUCTION
Insulin-like growth factor-1 (IGF-1) was confirmed to be positively correlated to growth rate in chicken. Chicken IGF-1 hormone mainly play roles in cell proliferation and increasing the metabolism rate1. Hepatic IGF-1 expression is partly regulated by the nutrition quality and quantity is mediated by growth hormone (GH)2. Synergy between the genetic potential and nutrition intake determines the meat-type chicken performance3.
Metabolizable Energy (ME) and Crude Protein (CP) levels are the two main variables that directly affect the cost of the feed. These parameters are important for developing an affordable feed that is also suitable for day-old chick (DOC) growth in Indonesia. Choosing the right dietary energy and protein source was important owing to the constant increase of feed demands in Indonesia4. Broiler chicken strains in Asia require 21% CP and 2800 kcal kg–1 ME5, whereas the most cultivated local chicken in Indonesia, Gallus gallus domesticus var. Kampong was reported to require 19% CP and 2900 kcal kg–1 ME6. However, the requirement of the new Indonesian hybrid chicken remains unknown.
The Indonesian hybrid chicken was obtained by crossing female layer chicken with a male Pelung chicken. Pelung chicken was selected as the parental line because it is the heaviest local chicken in Indonesia7; layer chickens are widely accepted for mass egg production. The Indonesian hybrid chicken is expected to possess the growth performance trait and provide economic benefits. Here, IGF-1 expression level was used to evaluate the appropriate dietary protein and energy level for the Indonesian hybrid chicken using two-step real-time PCR.
MATERIALS AND METHODS
Feed formulation: All feed formulae (A, B and C) were prepared using similar ingredients and produced by the local poultry feed mill (Smart-G, Semarang, Indonesia). The difference in energy and crude protein levels among these formulae occurred because of the alteration in the amount of fat and protein sources. Feed A contained 3100 kcal kg–1 ME and 22% CP, feed B contained 3300 kcal kg–1 ME and 20% CP and feed C contained 2800 kcal kg–1 ME and 21% CP.
Animals and experimental procedures: The experiments involving animals in this project were approved by the Universitas Gadjah Mada ethics committee. Female Layer chickens were obtained from a commercial egg producer and Pelung chickens were obtained from a university farm breeder. Naturally mated in 5×5×5 m3 pens, the chickens were given free access to feed and water. Eggs were collected for artificial incubation by a machine. The DOC hybrid chickens were divided into four groups (n = 10 per group). The first group (G-1) was fed with feed A, the second group (G-2) was fed with feed B, the third group (G-3) was fed with feed C and the fourth group (G-4) was starved for 3 days post-hatching before being fed with feed A. Chickens were given free access to water and feed. At 0, 3 and 7 days of age, chickens in all the groups were weighed. Five chickens from each group were randomly sampled for slaughtering to obtain liver samples at 7 days of age.
RNA isolation and cDNA synthesis: Total RNA isolation was directly performed after liver sampling using a commercial kit (Total RNA Mini Kit (Tissue), Geneaid, South Korea). Total RNA concentrations were measured by Qubit 2.0 Fluorometer (Invitrogen, USA). Total cDNA was produced using oligo (dT) primers by reverse transcription of 1 μL total RNA using a cDNA synthesis kit (Revert aid, Thermo Fisher Scientific, USA). Total RNA was subjected to DNase treatment before being reverse transcribed (DNase I, RNase-free Thermo Fisher Scientific). Total cDNA was subjected to quantification by real-time PCR using a commercial qPCR kit (Kapa Sybr Fast qPCR kits, USA) on CFX 96 thermal cycler (Bio-Rad Laboratories, USA). The IGF-I cDNA was amplified by specific primers (Forward: 5-TGG CCT GTG TTT GCT TAC CTT-3 and Reverse: 5-TAC GAA CTG AAG AGC ATC AAC CA-3) and β-actin (Forward: 5-CCC CAT GCC ATC CTC CGT CTG-3 and Reverse: 5-CCT CGG GGC ACC TGA ACC TCTC-3)8.
All specific primers were obtained from IDT (Singapore). The cycling procedure comprised a denaturation step (95°C for 3 min) and 40 cycles of amplification (denaturation at 95°C for 3 sec and annealing-extension at 60°C for 30 sec). The melting temperature was 75°C for IGF-1 primer and 84°C for β-actin primer. Each sample was run in triplicates. The gene expression level was estimated by comparing the Ct value of each group with that of the starved group (Ct)9.
Data analysis: Body weights and IGF-1 expression variance in each group because of the feed type were analyzed by one way ANOVA (p<0.05), followed by Tukey’s test. Correlation between body weights and IGF-1 expression level was evaluated by Pearson’s correlation test10. All data obtained were analyzed using SPSS 1311.
RESULTS
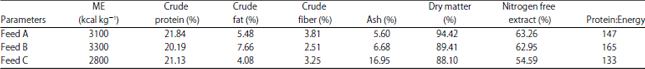
Effects of ME and CP on total weight: The feed content was derived using the proximate test and calorimeter bomb (Table 1). Proximate test results did not reveal any difference in feed quality between the formulated and commercial feeds.
The effect of varying CP and ME levels on the total weight of Indonesian hybrid chicken in each group is shown in Table 2. The weight gain of G-1 and G-2 was higher than that of G-3 and G-4 at the 3rd and 7th days (p<0.05). Feed intakes during 7 days in each feed group were not statistically different (p<0.05).
Effects of feed type on IGF-1 expression level: The IGF-1 expression level in the 7 days old hybrid chicken is shown in Fig. 1. The graph exhibits the highest gene expression level in G-1, followed by G-2 and G-3. The gene expression level in G-3 was higher than that in G-4, although both groups showed similar total weight.
Correlation between total weight and IGF-1 expression level: Correlation between gene expression and total weight of 7 days old chick after being subjected to Pearson’s correlation test showed that the total weight of the Indonesian hybrid chicken was highly correlated with IGF-1 expression level (Fig. 2) (R2 = 0.8). The heavier chicken showed a higher IGF-1 expression level (p<0.05).
DISCUSSION
These results indicate that suitable ME and CP levels for optimum growth of the Indonesian hybrid chicken was 3100 kcal kg–1 and 22% (protein:energy ratio, 141). Energy requirement of the hybrid chicken did not match the recommendation for the Indonesian local chicken6, which was 2700 kcal kg–1 or that of the Layer chicken, which was 2900 kcal kg–1 12. The difference in energy requirements occurred owing to higher metabolism of the hybrid chicken than that of the local pure breed chicken (Kampong chicken); the hybrid chicken needed more energy because of its higher growth rate13. Energy and protein requirement of the hybrid chicken were similar to those of the Super chicken (offspring of male Bangkok chicken and female Layer chicken) at 7 days of age14. This similarity was observed because of the same female parent, but the hybrid chicken from the Pelung sire seemed to weight more than the hybrid chicken from the Bangkok sire14.
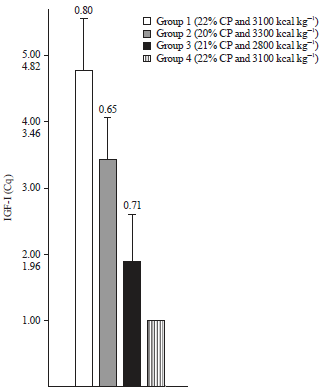 | |
| Fig. 1: | Relative IGF-1 expression level in the liver of the 7 days old Indonesian hybrid chicken after feeding (n = 5) and administration of normalized β-actin compared with that in the liver of the starved chicken. Standard deviation is mentioned on each bar |
| Table 1: | Proximate test on feed formula |
 | |
| Table 2: | Total weight of the Indonesian hybrid chicken in each feed group |
 | |
| aSignificant compared with groups 1, 2 and 3 (p<0.05), bSignificant compared with groups 1 and 2 (p<0.05) | |
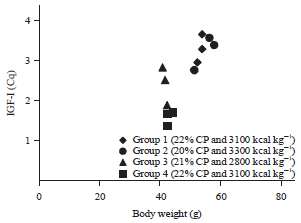 | |
| Fig. 2: | Correlation between the total weight of the 7 days-old Indonesian hybrid chicken and fold change in IGF-1 expression level |
Disparity in growth performance between the fed group and G-4 indicated that growth hormone sensitivity to nutrition was high in the hybrid chicken i.e., the growth hormone will increase the weight of the hybrid chicken with increased dietary intake. Similar results were reported for broiler chickens7, but a greater contrast was observed between the fed and starved groups with respect to weight gain. Starved broiler chickens at 5-7 days of age could catch up performance at harvest. However, this did not occur in the Indonesian hybrid chicken at first 3 days post-hatching. The ability of the hybrid chicken to compensate for the weight loss after starving was not well supported because they did not have as responsive an endocrinal system as the broiler chicken15.
The effect of dietary energy on the performance of growing birds depends on the capacity of the bird to alter feed intake for meeting changing demands with respect to their calories16. Unequal capacity in each strain was because the broiler chicken was formed after long genetic selection, whereas, the Indonesian hybrid chicken was an F1 resulting from the Pelung and layer chicken. The Indonesian hybrid chicken was suggested to be more heterozygous than the broiler chicken.
Dietary proteins and energy content influence the IGF-1 mRNA abundance in rat liver17. The nitrogen source from the digested protein in the feed is required for new protein synthesis2. The nitrogen source and amino acid trigger GH-IGF-1 system activation via neurotransmitters18,19. Dopamine and other neurotransmitter influence GHRH secretion in the hypothalamus during the absence of somatostatin (somatotropin-releasing inhibitory factor)19. Furthermore, circulating GH in the liver induces IGF-1 expression18. High protein in the feed increases IGF-1 mRNA levels. The fold change value of G-1 suggested that feed A dietary energy accommodated growth that allowed high expression of IGF-1 in the liver. The G-2 that shows similar weight gain at 7 days of age exhibits lower fold change of IGF-1. Therefore, the fold change in G-3 was the lowest, indicating that this CP and ME combination did not accommodate tissue development. Compared with feeds A and B, feed C did not have enough energy for sufficient body metabolism. Low ME in Isonitrogenous diet treatment resulted in low growth performance in broiler chickens12. Feed C had a high protein level but did not improve weight gain of the chicken likely owing to the lower ME. Sufficient energy derived from glucose, fatty acid and other energy sources drives the metabolism of the whole body.
The growth rate of G-4 was higher than that of G-3, but the gene expression level of G-4 was lower than that of G-3 because the re-feeding program 3 days post-hatching in G-4 prompted a very low gene expression even when the best feed was administrated. Low gene expression indicated that optimum gene expression was not observed if administration of the feed was delayed. Weight gain between G-3 and G-4 was not statistically different because of the involvement of extra-hepatic IGF-1 mRNA expression in the target tissue. In contrast, after 3 h of re-feeding, IGF-1 mRNA increased to 3-fold of that before starving in broiler chickens but decreased after 7 h nearly to that before starving. Feed containing adequate protein drives the processes of the whole body in rats17.
Dietary energy and IGF-1expression level were highly correlated with total weight in 7 days-old chicks (R2 = 0.8). We suggest using 22% CP and 3100 kcal kg–1 ME feed for the Indonesian hybrid chicken in the starter phase.
SIGNIFICANCE STATEMENTS
New hybrid chickens, obtained by crossing female layer with male Pelung chicken (the Indonesian native chicken), were evaluated to be positioned as meat-type in Indonesia. Here we attempted to develop a starter-feed formula that fulfilled the hybrid chicken growth requirement.
ACKNOWLEDGMENT
The authors would like to thank PIAT UGM and Dr. Yuny Erwanto, Faculty of Animal Science, UGM for providing the breeding facility. This study was supported by Ministry of Research, Technology and Higher Education (grant No. RT-2015-0238, 2015). The authors would like to thank Enago (www.enago.com) for the English language review.
REFERENCES
- Giachetto, P.F., E.N. Guerreiro, J.A. Ferro, M.I.T. Ferro, R.L. Furlan and M. Macari, 2003. Performance and hormonal profile in broiler chickens fed with different energy levels during post restriction period. Pesquisa Agropecuaria Brasileira, 38: 697-702.
CrossRefDirect Link - Kaput, J. and R.L. Rodriguez, 2004. Nutritional genomics: The next frontier in the postgenomic era. Physiol. Genomics, 16: 166-177.
CrossRefDirect Link - Fekete, S. and D. Brown, 2007. Veterinary aspects and perspectives of nutrigenomics: A critical review. Acta Veterinaria Hungarica, 55: 229-239.
CrossRefDirect Link - Nawaz, H., T. Mushtaq and M. Yaqoob, 2006. Effect of varying levels of energy and protein on live performance and carcass characteristics of broiler chicks. J. Poult. Sci., 43: 388-393.
CrossRefDirect Link - Lin, S., H. Li, H. Mu, W. Luo and Y. Li et al., 2012. Let-7b regulates the expression of the growth hormone receptor gene in deletion-type dwarf chickens. BMC Genom., Vol. 13.
CrossRefDirect Link - Livak, K.J. and T.D. Schmittgen, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25: 402-408.
CrossRefDirect Link - Benjamini, Y. and Y. Hochberg, 1995. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B: (Methodol.), 57: 289-300.
Direct Link - Morris, T.R., 1968. The effect of dietary energy level on the voluntary calorie intake of laying birds. Br. Poult. Sci., 9: 285-295.
CrossRefDirect Link - Kim, J.W., 2010. The endocrine regulation of chicken growth. Asian-Austr. J. Anim. Sci., 23: 1668-1676.
CrossRefDirect Link - Saragih, H.T.S.S.G., R.T. Utomo, A.B.I. Perdamaian, U.E. Puspita and I. Lesmana et al., 2016. The effect of early posthatch local feed in pectoralis muscle of jawa super chicks (Gallus gallus domesticus). Proceedings of the 1st International Conference on Science and Technology, November 11-13, 2015, Yogyakarta, Indonesia.
- Kamran, Z., M. Sarwar, M. Nisa, M.A. Nadeem, S. Mahmood, M.E. Babar and S. Ahmed, 2008. Effect of low-protein diets having constant energy-to-protein ratio on performance and carcass characteristics of broiler chickens from one to thirty-five days of age. Poult. Sci., 87: 468-474.
CrossRefDirect Link - Rosa, P.S., D.E. Faria Filho, F. Dahlke, B.S. Vieira, M. Macari and R.L. Furlan, 2007. Effect of energy intake on performance and carcass composition of broiler chickens from two different genetic groups. Revista Brasileira Ciencia Avicola, 9: 117-122.
CrossRefDirect Link - Emler, C.A. and D.S. Schalch, 1987. Nutritionally-induced changes in hepatic Insulin-like Growth Factor I (IGF-I) gene expression in rats. Endocrinology, 120: 832-883.
CrossRefDirect Link - Scacchi, M., A.I. Pincelli and F. Cavagnini, 2003. Nutritional status in the neuroendocrine control of growth hormone secretion: The model of anorexia nervosa. Front. Neuroendocrinol., 24: 200-224.
CrossRefPubMedDirect Link - Garcia-Tornadu, I., G. Risso, M.I. Perez-Millan, D. Noain and G. Diaz-Torga et al., 2010. Neurotransmitter modulation of the GHRH-GH axis. Pituitary Today, 38: 59-69.
CrossRefDirect Link