
Fawziah A. Al-Salmi
Department of Biology, Faculty of Science, Taif University 888, Taif, Saudi Arabia
Rasha Al-Eisa
Department of Biology, Faculty of Science, Taif University 888, Taif, Saudi Arabia
Reham Z. Hamza
Department of Biology, Faculty of Science, Taif University 888, Taif, Saudi Arabia
Howayda E. Khaled
Department of Zoology, Faculty of Science, Suez University, Suez, Egypt
Nahla S. El-Shenawy
Department of Zoology , Faculty of Science, Suez Canal University, Ismailia, 41522, Egypt
LiveDNA: 20.782
International Journal of Pharmacology
Year: 2019 | Volume: 15 | Issue: 2 | Page No.: 248-256







ABSTRACT
Background and Objective: Sodium Valproate (SV) is a medicine that is used to treat epilepsy and to prevent headache. In this study, L-cysteine (LC) was used to decrease the stress and biochemical variations induced by SV treatment. The present study investigated the defensive actions of LC vs, SV that induced testicular impairment. Materials and Methods: The rats were split into six groups (n = 10) as following: 1st control animals were saline-treated, 2nd and 3rd groups were administrated two doses of SV (100 and 500 mg kg–1 b.wt.) presenting low and high doses, respectively, 4th group was treated with 100 mg kg–1 of LC, in addition 5th and 6th groups were treated with SV-LD+LC and SV-HD+LC, respectively. The experiment was run for 30 successive days. Weights of the testis, serum testosterone, testicular oxidative/anti-oxidant capacity and histopathological damage scores of testis were recorded. Results: The SV group had significantly increased the tissue oxidative stress markers and significantly declined all antioxidant enzymes activities as compared to the control group. When the animals treated with combination of LC and any dose of SV, levels of oxidative parameters significantly declined, as well as the anti-oxidant significantly elevated compared to the SV-LD and SV-HD groups. The Johnsen's testicular score values showed improvement when LC was co-treated with the SV. Conclusion: Current results indicated that LC had partial protective effects against SV-induced testis damage at the biochemical and histopathological levels that could be due to the enhanced tissue anti-oxidant capacity.
PDF Abstract XML References Citation
Received: October 16, 2018;
Accepted: November 21, 2018;
Published: January 15, 2019
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Fawziah A. Al-Salmi, Rasha Al-Eisa, Reham Z. Hamza, Howayda E. Khaled and Nahla S. El-Shenawy, 2019. Protective Effect of L-cysteine Against Sodium Valproate-induced Oxidant Injury in Testis of Rats. International Journal of Pharmacology, 15: 248-256.
DOI: 10.3923/ijp.2019.248.256
URL: https://scialert.net/abstract/?doi=ijp.2019.248.256
DOI: 10.3923/ijp.2019.248.256
URL: https://scialert.net/abstract/?doi=ijp.2019.248.256
INTRODUCTION
L-cysteine (LC) is one of the important exogenous antioxidants that shelter against oxidative impairment. This effect could be related to the LC itself (inactivated the free radicals) or to the induction of reduced glutathione (GSH) formation1. Ahmed et al.2 evaluated some anti-oxidants activity of LC and they found that it antagonizes cisplatin (CP) induced testicular injury of rats. It also reduced the lipid peroxidation (LPO) and enhanced the antioxidant levels of the rats treated with CP3.
The epileptics require suitable treatment to prevent mortality4. Sodium Valproate (SV) is one of the anti-epileptic drugs that have a diverse mechanism of actions which were attributing to its toxicity and efficacy. It was also used in bipolar disorder, migraine prophylaxis, schizophrenia and depression5. Acid SV is the famous drug that can control all types of seizures incorporated with the idiopathic epilepsies6.
Hamza et al.7 found that LC administration inhibited liver injury and improved the redox state of rats due to SV-toxicity as well as improved the nephrotoxicity induced by SV8. Moreover, the protective effect of LC in brain tissue of rat was mediated through attenuation of oxidative stress, suggesting a therapeutic role for LC in the side effect of epilepsy, when treated with SV9.
Animal models had revealed that SV causes reversible changes in sperm motility and count in addition, the histoarchitecture of the testis10,11. The histopathological examination showed that SV (400 mg kg–1) had caused degeneration of germinal cells and decrease in the Johnsen’s scoring after 14 days of exposure12.
Spermatogenesis process occurs by the action of many hormones including Follicle-stimulating Hormone (FSH), which is secreted by the pituitary gland to stimulate the Sertoli cells. Luteinizing Hormone (LH) secreted from Leydig cell, stimulates testosterone synthesis. Therefore, these hormones were used as markers of testicular activity in males13. Henriksen et al.14 reported that rats have shown a decline in testosterone levels caused by losing the germ cell.
There is no data available on the effects of LC against LPO, NO⋅ and OH– as well as the histological construction of the rat testis tested with SV. It is commonly known that LC prevents neurotoxicological damages as well as the neoprene and hepatotoxicity. So, the study was concerned to describe the injurious effects of SV on the testis weight and oxidative damage. The mechanisms of this protective effect against testicular toxicity may be due to the enhanced tissue anti-oxidant capacity. Thus, this study was full of clinical importance as it focuses on the testicular damage in case of treatment with SV and the role of LC that could reduce the testis damage. Therefore, the aim of the existing study was to research the possible protective importance of LC in the testis toxicity of male rats induced by SV.
MATERIALS AND METHODS
Chemicals: All chemicals were of high analytical grade and obtained from Sigma Chemical Co., St. Louis, Mo., USA.
Experimental animals and design: The Wistar rats weighing 200±5 g were brought from Faculty of Veterinary Medicine, Zagazig University. They were housed under standard laboratory conditions with 12/12 light and dark cycle, with suitable temperature and humidity. The European Community Directive (86/609/EEC) on animal care guidelines has been followed for the Care and Use of Laboratory Animals 8th Edition. Animal experiments received approval from the Ethical Committee of the Pharmacy Faculty of Zagazig University (No. P22/2/2015). The experiment was carried out during July-September, 2015.
The rats were allocated to one group served as control and five-treated groups of 10 animals each. All the rats were orally medicated by gastric tube for 30 successive days. The 1st control group received saline, the 2nd and 3rd groups were given the lower and the higher doses of SV (SV-LD, SV-HD; 100 and 500 mg kg–1, respectively) according to Khan et al.6, 4th group was treated with 100 mg kg–1 L-cysteine2, 5th and 6th groups were given SV-LD+LC and SV-HD+LC as the above doses, respectively. The higher dose of SV was selected as described before by Leclair-Visonneau et al.15 depending on the clinical experiment.
Organo-somatic index of the testis: The body weight of each rat was recorded in the beginning and on the determinative of the experiment. Then, the rats were sacrificed and testis was removed to weight them. From these values, the Organo-Somatic Index (OSI) of testis was calculated using the equation cited by Chirumari and Reddy16.
Preparation of testis homogenates: The testis was homogenized in 10 mL cold buffer per gram tissue using a Potter-Elvehjem homogenizer in 10 volumes of buffer17. The homogenate was centrifuged at 10,000 Xg or 2500 Xg for 20 min at 4°C for estimating antioxidant enzymes assays and MDA level, respectively. The supernatant was preserved in a deep freeze until being used for various assays.
Determination of oxidative biomarkers: The LPO was measured as malondialdehyde (MDA) as described by Ohkawa et al.18. 1,1,3,3-tetra-ethoxy propane as standard was used to determine MDA levels which expressed as μM g–1 tissue. Nitric oxide (NO∙) reacts with oxygen to produce stable products (nitrate and nitrite) that had been estimated using Griess reagent19.
Determination of hydroxyl radical scavenging activity of L-cysteine: The evaluation of hydroxyl radical capacity of LC was measured according to Halliwell et al.20. The percentage inhibition was calculated using the equation:
where, A0 was the absorbance of the control reaction and A1 was the absorbance in presence of the sample and reference.
Determination of enzymatic anti-oxidant biomarkers: The superoxide dismutase (SOD) activity was expressed as U mg–1 protein according to the method of Marklund and Marklund21. Catalase (CAT) activity was presented as mM mg–1 protein according to Aebi22. Glutathione peroxidase (GPx) activity was evaluated by the Hafeman et al.23. The activity of GPx was expressed in terms of mM GSH consumed/min/g wet weight tissue. Glutathione reductase (GRX) activity was measured by Couri and Abdel-Rahman24. The unit of GRx activity was defined as an increase in the log (GSH) of 0.001 per min.
Determination of non-enzymatic anti-oxidant: Total anti-oxidant capacity was evaluated using ferric reducing anti-oxidant power assay25.
Determination the testosterone hormone level: Plasma testosterone levels were measured by electrochemiluminescence by automata (Elecsys, Roche Diagnostics)26.
Histological evaluation: A part of the testis was fixed in 10% neutral buffered formalin and embedded in paraffin. The paraffin blocks were sectioned into slices in 5 μ thickness, mounted onto slides and stained with hematoxylin and eosin as described27. The histological evaluations, general characters of tissues, the progressive degeneration of the tubule, proceeding to the loss of spermatogonia and number of Sertoli cells were examined semi quantitatively under a light microscope. The data in each group were reported as Means±SE based on the sum of the score histological criteria of spermatogenesis28.
Statistical analysis: All analyses were performed using SPSS statistical version 20 (SPSS Inc., USA). Data were presented as mean values ±SE (n = 10). To assess significant differences among treated-groups, statistical analysis was performed using two-way analysis of variance (ANOVA). The criterion for statistical significance was set at p<0.05.
RESULTS
Organo-somatic index and biochemical parameters: The OSI of the testis in the SV group was significantly declined as compared to the control rats, according to the dose by 41.2 and 57.1% for SV-LD and SV-HD, respectively (Fig. 1). There was a significant rise in OSI of the testis of the groups received LC with different doses of SV by 1.2- and 1.7-fold for SV-LD+LC and SV-HD+LC, respectively.
The levels of testosterone in control and LC groups were approximately the same. The testosterone level diminished only in rats that were treated with SV-HD. The LC did not improve the level of testosterone when given with SV-LD and SV-HD as compared with SV groups (Fig. 2).
The testicular oxidative parameters of NO and LPO levels were significantly elevated by SV in a dose-dependent manner after 30 days of treatment (Fig. 3).
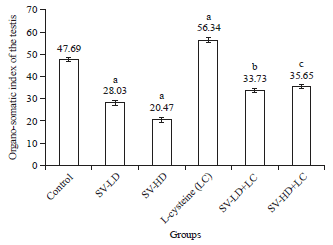 | |
| Fig. 1: | Effect of sodium valproate and L-cysteine separately or in combination with the organo-somatic index of the testis tissue of rats |
| Values were expressed as Mean±SE, (n = 10), aSignificant difference as compared to the control group, bSignificant difference as compared to SV-LD, cSignificant difference as compared to SV-HD | |
The LC declined all the oxidative parameters as compared to control rats, especially the LPO level which was significantly decreased. The decline in intratesticular oxidative levels was noticed in LC and SV combined groups as compared to its related group of SV (low or high dose). There were no significant alternations in the NO levels.
L-cysteine -OH radical activity was evaluated and showed that the concentration 100 μg mL–1 had high -OH free radical activity that confirmed the LC anti-oxidant activity (Fig. 4).
Data in Table 1 revealed that there was no remarkable change in SOD, CAT, GPx, GRx and TAC of control rats and those treated with LC. In contrast, the reduction in all the previous mentioned anti-oxidant enzymes activities and TAC level were recorded in rats treated with SV-LD or SV-HD for 30 days. Administration of the LC with different doses of SV resulted in a significant elevation of anti-oxidant enzyme activity and improved the TAC content as compared to its related group of SV.
Histopathology evaluation: The histological appearance of the testicular tissues of the control group was normal (Fig. 5a). Control group showed normal cells of spermatogenic, spermatocytes and rounded spermatids attached in the layers of seminiferous tubules. Also, LC-treated rats had normal seminiferous tubules with normal spermatogenesis (Fig. 5b). SV-LD induced histopathological variations in the testis such as necrosis, germ cell degeneration, desquamation, edema and congestion in addition to degeneration and atrophy of seminiferous tubules (Fig. 5c). It was observed that there was a marked increase in necrotic and degenerative changes in germinal cells of rats that received SV-HD (Fig. 5d). The improvement was also observed in SV-LD and LC treated groups where there was partial restoration of germinal cells in addition, seminiferous tubules lined by several layers of spermatogenic cells up to sperm formation but surrounded by mild edematous stroma (Fig. 5e). The well-defined germinal cells with the reduction in necrosis, edema and congestion were found in the animals that received SV-HD and LC together (Fig. 5f).
Histological Activity Index (HAI) was evaluated using the degree of microscopic lesions in testis tissue as the effect of SV and LC separately or in combination (Fig. 6). A decrement in scoring was noticed in the SV exposed group depend on the dose as compared to the normal animals. An improvement in score was noted in the rats that were given LC with SV, thus showing its protective action.
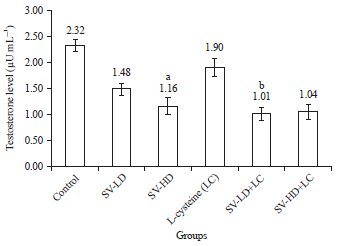 | |
| Fig. 2: | Effect of sodium valproate and L-cysteine separately or in combination, on testosterone levels of the testis tissue of rats |
| Values were expressed as Mean±SE, (n = 10), aSignificant difference as compared to the control group, bSignificant difference as compared to its related group of SV-LD | |
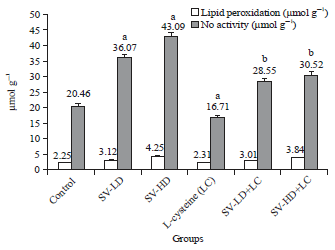 | |
| Fig. 3: | Effect of sodium valproate and L-cysteine separately or in combination on the levels of lipid peroxidation and nitric oxide of the testis tissue of rats |
| Values were expressed as Mean±SE, n = 10, aSignificant difference as compared to the control group, bSignificant difference as compared to its related group of SV-LD | |
| Table 1: | Effect of sodium valproate and L-cysteine separately or in combination on enzymatic and non-enzymatic antioxidant of the testis of male rats |
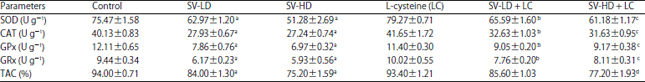 | |
| Values were expressed as Mean±SE, (n = 10), Rats treated with low and high doses of SV (100 and 500 mg kg–1, respectively), aSignificant difference as compared to the control group, bSignificant difference as compared to SV-LD,cSignificant difference as compared to SV-HD, dSignificant difference as compared to SV-LD+LC | |
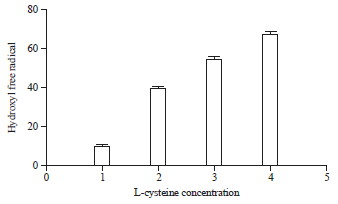 | |
| Fig. 4: | Free hydroxyl radical activity (–OH) of L-cysteine was presented as μM g–1 |
DISCUSSION
This study was designed to assess the effects of LC intake on SV-induced testis toxicity of male rats after exposure to two different doses. According to the literature, this is the first study that evaluates the defensive effects of LC against testicular damage induced by SV in experimental animals.
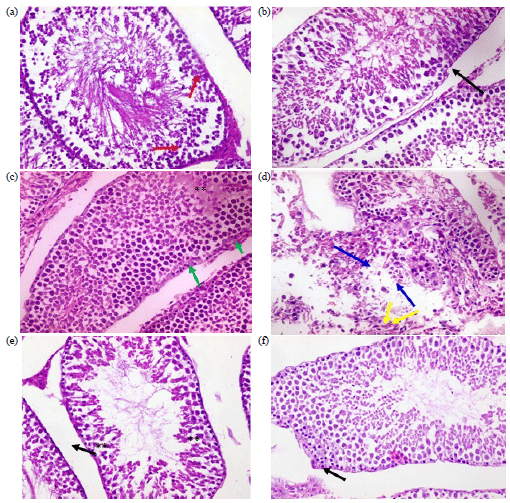 | |
| Fig. 5(a-f): | Histopathological slides of the testis stained with haematoxylin and eosin in (a) Control groups of rat showing normal seminiferous tubules lined by several layers of spermatogenic cells (Red arrow) up to sperm formation (400X), (b) Cross section of rat testis treated with L-cysteine (400 mg kg–1) showing normal seminiferous tubules (Black arrow) with normal spermatogenesis (400X), (c) Sodium valproate (100 mg kg–1) treated rats showing distended seminiferous tubules (Blue arrow) by 1 and 2 spermatogenic cells with few spermatids and number of sperms (**). i.e., maturation arrest (400X), (d) Sodium valproate (500 mg kg–1) showing necrosis and ruptured tubules (Blue arrow) with infiltration by macrophages (Black arrow) and plasma cells (400X), (e) Low dose of SV and L-cysteine showing partial restoration of germinal cells as well as seminiferous tubules lined by several layers of spermatogenic cells (**) up to sperm formation but surrounded by mild edematous stroma (Black arrow) (400X) and (f) High dose of SV and L-cysteine showing reduction in necrosis, edema and congestion of testicular tissue with seminiferous tubules surrounded by edematous stroma (Black arrow) containing small groups of Leydig cells (400X) |
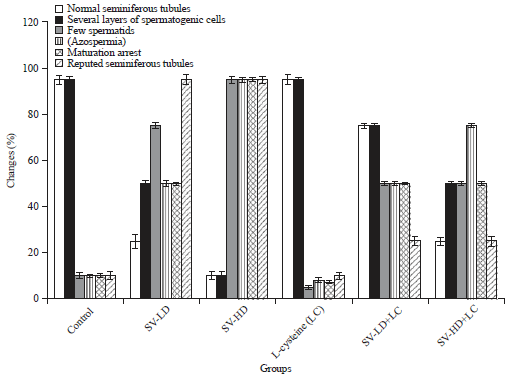 | |
| Fig. 6: | Histological Activity Index (HAI) was assessed as percentage based on the degree of microscopic lesions in testis tissue as the effect of sodium valproate and L-cysteine separately or in combination on this basis - no lesions (0-10%), ---+ lesions found in 1-3 rats (25%), ++-- lesions found in 4-6 rats (50%), +++- lesions found in 7-8 rats (75%), ++++ lesions found in 9-10 rats at least (95%) (n = 10) |
The reason to select the high dose of SV was to mimic the accidental exposure or increasing the time of exposure to the low dose.
The results established that the testosterone level had significantly declined in SV groups when compared with control. This observation is parallel to previous report by Bauer et al.29 and Vijay et al.30 who found that valproate acts directly on the testis, to inhibit testosterone synthesis by inhibiting the effect of the Leydig cells. Moreover, Soliman and Abla31 had reported that there was a decline in the levels of testosterone, FSH and LH after exposure to SV that diminished by part the germ cell number. The intensity of damage occurred by SV was partially prevented by LC that may be influencing testosterone synthesis.
In conformity with the obtained results, SV affected the reproductive capacity in general as it is effective at the biochemical and cellular levels of the testis. It is acceptable to suggested that valproate could not be the first anti-epileptic drug of consideration for the patient32.
It was found that there was a correlation between the histological and oxidative status induced by SV. The testis is the major target organ for oxidative stress that could be due to its high content of polyunsaturated lipids membrane33. In the present study, SV-treatment induced LPO as indicated by a significant elevation in TBARS level that causes irreparable cell damage in testis tissue by interfering with plasma membrane lipids.
The results showed that the concentrations of LPO and NO were significantly elevated, which were incorporated with a reduction in anti-oxidant enzymes (SOD, CAT, GPx and GRx) activities in SV-treated groups. However, there was a significant decline in LPO values in SV+LC treated-group when compared with SV-treated rats. Similarly, Vidya and Subramanian34 found that SV-treatment was found to raise the levels of MDA and hydroperoxides, in addition, the decline in the enzymatic anti-oxidants and level of glutathione (GSH) of rats. In the erythrocytes of children with epilepsy treated with SV, the activities of anti-oxidant enzymes were insignificantly lower, whereas the MDA concentration was insignificantly higher35.
The data presented were in parallel with Zhang et al.36 who found the MDA levels in neutrophils of SV-treated patients were higher, while the activities of SOD and CAT were significantly declined than the control groups. Thus, the testicular damage that was induced in rats could be attributed to the oxidative stress provoked by SV.
The LC has antioxidant characters because of its ability to go through the redox reactions. Cysteine can be formed in the human body under normal physiological conditions if there was enough quantity of methionine. Cysteine's anti-oxidant properties were found in the tripeptide GSH37. Therefore, one of the important aims of the current research was to highlight the role of LC in reducing oxidative stress and in case of treatment with SV was to increase the GSH content of the cell. This appeared clearly in SV-LD and SV-HD treated-groups where GPx and GRx activities decreased in a dose-dependent manner. These results proved that LC had anti-oxidant activity due to its mechanism of action. The cysteine could be offered the role of increasing the beneficial antioxidant GSH, which was diminished in oxidative stress status. The LC could be used as antioxidant but it was the first time, according to our knowledge to be used with SV.
Previous study by Hamza et al.7, El-Shenawy and Hamza8 and Jain et al.38 showed that LC was a strong anti-oxidant compound and protects liver, kidney and brain tissues against the oxidative stress. Droge39 revealed that LC treatment intercepts oxidative stress and thus the obtained result was committed by our conclusion. This effect may be directly concomitant to the LC itself 1 whereas, the conservation of free sulfhydryl groups was appraised crucial to the biological functions of many proteins39.
The histopathological discovery showed that SV-treatment caused major structural changes in testis compared to the control rats. However, it was determined that LC treatment alleviated the toxic effects of SV on testis when given together with SV. Girish et al.12 found that the testis was extremely sensitive to the side effect of SV.
Moreover, histopathological observation in the testis of rats treated with SV revealed vascular changes in the form of congestion and interstitial edema and degenerative alternation with the presence of multinucleate spermatid giant cells. Similarly, Cansu10, Bairy et al.11 and Girish et al.12 reported that SV-treatment caused necrosis, atrophy in the seminiferous tubules, multinucleated giant cell formation, interstitial edema with congestion, a reduction in the germinal cell count and impaired spermatogenesis. A decline in Johnsen’s testicular score was also realized in the histological observation as an effect of SV treatment. The histo-architecture of the testis and Johnsen’s scores were partially-restored to nearly normal by LC treatment with SV.
The SV can disrupt the cellular mechanisms in different ways that can cause toxicity. It may induce the formation of LPO, NO and OH–, which were chemical mechanisms capable of changing the structure and function of the testis. The antioxidant properties of LC may play a critical role in preventing the toxicity of drugs, especially that of epilepsy. The mechanism elaborated in the protection of LC against SV induced testicular toxicity was unknown but our obtained results confirmed that the anti-oxidant activities of LC that were carried out by performing –OH radical activity, confirmed its anti-oxidant activity, that may contribute by way or another in the protection of testis tissues from oxidative stress induced by SV treatment either in the lower or the higher dose.
CONCLUSION
Sub-chronic exposure to two doses of SV altered the anti-oxidant enzymatic activities and induced oxidative damage in addition to the histopathological alterations of the testis in a dose-dependent manner. The co-administration of SV with LC showed the beneficial effect on all the previous parameters. The histoarchitecture of the testis was also partially-restored by LC confirming its protective effect against SV. This action of LC may be closely related to its anti-oxidant capacities and its ability to scavenge free radicals and thus protect testicular tissues from oxidative damage of SV.
SIGNIFICANCE STATEMENT
This study discovered the protective effect of L-cysteine against the toxicity of sodium valproate.
The study can be beneficial for declining the side effect of sodium valproate on the testes using L-cysteine.
This study will help the researchers to uncover the crucial areas of sodium valproate toxicity that many researchers were not able to explore.
REFERENCES
- Kerksick, C. and D. Willoughby, 2005. The antioxidant role of glutathione and N-acetyl-cysteine supplements and exercise-induced oxidative stress. J. Int. Soc. Sports Nutr., Vol. 2.
CrossRefDirect Link - Ahmed, E.A., H.M. Omar, S.K.A. Elghaffar, S.M.M. Ragb and A.Y. Nasser, 2011. The antioxidant activity of vitamin C, DPPD and l-cysteine against cisplatin-induced testicular oxidative damage in rats. Food Chem. Toxicol., 49: 1115-1121.
CrossRefPubMedDirect Link - Sayed, M.M., K.M. Hassanein and W. Senosy, 2014. Protective effects of thymoquinone and l-cysteine on cadmium-induced reproductive toxicity in rats. Toxicol. Rep., 1: 612-620.
CrossRefDirect Link - Tiamkao, S., K. Sawanyawisuth and A. Chancharoen, 2013. The efficacy of intravenous sodium valproate and phenytoin as the first-line treatment in status epilepticus: A comparison study. BMC Neurol., Vol. 13.
CrossRefDirect Link - Khan, M., S.T. Khattak and M. Elahi, 2011. Gross hepatic changes in developing albino rats exposed to valproic acid. J. Ayub. Med. Coll. Abbottabad, 23: 48-50.
Direct Link - Hamza, R.Z., H.A.A. Ismail, H.E. Khaled and N.S. El-Shenawy, 2015. Antioxidant effect of L-cysteine on sodium-valproate-induced oxidative stress in rat liver: Biochemical and ultrastructural approaches. Toxicol. Environ. Chem., 97: 1041-1056.
CrossRefDirect Link - El-Shenawy, N.S. and R.Z. Hamza, 2016. Nephrotoxicity of sodium valproate and protective role of L-cysteine in rats at biochemical and histological levels. J. Basic Clin. Physiol. Pharmacol., 27: 497-504.
CrossRefDirect Link - Hamza, R.Z. and N.S. El-Shenawy, 2017. The beneficial effects of L-cysteine on brain antioxidants of rats affected by sodium valproate. Hum. Exp. Toxicol., 36: 1212-1221.
CrossRefDirect Link - Cansu, A., 2010. Antiepileptic drugs and hormones in children. Epilepsy Res., 89: 89-95.
CrossRefDirect Link - Bairy, L., V. Paul and Y. Rao, 2010. Reproductive toxicity of sodium valproate in male rats. Indian J. Pharmacol., 42: 90-94.
CrossRefDirect Link - Girish, C., O. Shweta, V. Raj, S. Balakrishnan and R.G.B. Varghese, 2014. Ellagic acid modulates sodium valproate induced reproductive toxicity in male Wistar rats. Indian J. Physiol. Pharmacol., 58: 416-422.
Direct Link - Sofikitis, N., N. Giotitsas, P. Tsounapi, D. Baltogiannis, D. Giannakis and N. Pardalidis, 2008. Hormonal regulation of spermatogenesis and spermiogenesis. J. Steroid Biochem. Mol. Biol., 109: 323-330.
CrossRefDirect Link - Henriksen, E., H. Harkovita and M. Parvinem, 1995. Testosterone inhibits and induced apoptosis in rats seminiferous tubule in a stage-specific manner: In situ quantification in squash preparations after administration of ethane dimethane sulfonate. Endocrinology, 136: 3285-3291.
PubMed - Leclair-Visonneau, L., T. Rouaud, B. Debilly, F. Durif and J.L. Houeto et al., 2016. Randomized placebo-controlled trial of sodium valproate in progressive supranuclear palsy. Clin. Neurol. Neurosurg., 146: 35-39.
CrossRefDirect Link - Chirumari, K. and P.K. Reddy, 2007. Dose-dependent effects of fluoride on neurochemical milieu in the hippocampus and neocortex of rat brain. Res. Rep. Fluoride, 40: 101-110.
Direct Link - Lebda, M., S. Gad and H. Gaafar, 2014. Effects of lipoic acid on acrylamide induced testicular damage. Materia Socio-Medica, 26: 208-212.
Direct Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Marcocci, L., J.J. Maguire, M.T. Droylefaix and L. Packer, 1994. The nitric oxide-scavenging properties of Ginkgo biloba extract EGb 761. Biochem. Biophys. Res. Commun., 201: 748-755.
CrossRefDirect Link - Halliwell, B., J.M. Gutteridge and O.I. Aruoma, 1987. The deoxyribose method: A simple "test-tube" assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem., 165: 215-219.
CrossRefDirect Link - Marklund, S. and G. Marklund, 1974. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem., 47: 469-474.
CrossRefPubMedDirect Link - Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Hafeman, D.G., R.A. Sunde and W.G. Hoekstra, 1974. Effect of dietary selenium on erythrocyte and liver glutathione peroxidase in the rat. J. Nutr., 104: 580-587.
CrossRefPubMedDirect Link - Couri, D. and M.S. Abdel-Rahman, 1979. Effect of chlorine dioxide and metabolites on glutathione dependent system in rat, mouse and chicken blood. J. Environ. Pathol. Toxicol., 3: 451-460.
PubMedDirect Link - Prieto, P., M. Pineda and M. Aguilar, 1999. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem., 269: 337-341.
CrossRefDirect Link - Wheeler, M.J., 1995. The determination of bio-available testosterone. Ann. Clin. Biochem., 32: 345-357.
CrossRefDirect Link - Johnsen, S.G., 1970. Testicular biopsy score count: A method for registration of spermatogenesis in human testes: Normal values and results in 335 hypogonadal males. Hormones, 1: 2-25.
CrossRefPubMedDirect Link - Bauer, J., S. Blumenthal, M. Reuber and B. Stoffel-Wagner, 2004. Epilepsy syndrome, focus location and treatment choice affect testicular function in men with epilepsy. Neurology, 62: 243-246.
CrossRefDirect Link - Vijay, P., R. Yeshwanth and K.L. Bairy, 2008. The effect of sodium valproate on the biochemical parameters of reproductive function in male albino Wistar rats. Indian J. Pharmacol., 40: 248-250.
Direct Link - Soliman, G.A. and A. Abla, 1999. Effects of antiepileptic drugs carbamazepine and sodium valproate on fertility of male rats. DTW. Deutsche Tierarztliche Wochenschrift, 106: 110-113.
Direct Link - Mikkonen, K., L.K. Vainionpaa, A.J. Pakarinen, M. Knip, I.Y. Jarvela, J.S. Tapanainen and J.I. Isojarvi, 2004. Long-term reproductive endocrine health in young women with epilepsy during puberty. Neurology, 62: 445-450.
PubMedDirect Link - Debnath, D. and T.K. Mandal, 2000. Study of quinalphos (an environmental oestrogenic insecticide) formulation (Ekalux 25 EC)‐induced damage of the testicular tissues and antioxidant defence systems in Sprague‐Dawley albino rats. J. Applied Toxicol.: Int. J., 20: 197-204.
CrossRefDirect Link - Vidya M. and P. Subramanian, 2006. Effects of ∝-ketoglutarate on antioxidants and lipid peroxidation products in rats treated with sodium valproate. J. Applied Biomed., 42: 141-146.
Direct Link - Sobaniec, W., E. Solowiej, W. Kulak, L. Bockowski, J. Smigielska-Kuzia and B. Artemowicz, 2006. Evaluation of the influence of antiepileptic therapy on antioxidant enzyme activity and lipid peroxidation in erythrocytes of children with epilepsy. J. Child Neurol., 21: 558-562.
CrossRefDirect Link - Zhang, Y.J., M. Zhang, X.C. Wang, Y.H. Yu, P.J. Jin and Y. Wang, 2011. Effects of sodium valproate on neutrophils' oxidative metabolism and oxidant status in children with idiopathic epilepsy. Zhonghua Er Ke Za Zhi, 49: 776-781.
Direct Link - Baker, D.H. and G.L. Czarnecki-Maulden, 1987. Pharmacologic role of cysteine in ameliorating or exacerbating mineral toxicities. J. Nutr., 117: 1003-1010.
CrossRefDirect Link - Jain, S.K., T. Velusamy, J.L. Croad, J.L. Rains and R. Bull, 2009. L-Cysteine supplementation lowers blood glucose, glycated hemoglobin, CRP, MCP-1 and oxidative stress and inhibits NF-κB activation in the livers of Zucker diabetic rats. Free Radical Biol. Med., 46: 1633-1638.
CrossRefDirect Link - Droge, W., 2005. Oxidative stress and ageing: Is ageing a cysteine deficiency syndrome? Philos. Trans. R. Soc. London B: Biol. Sci., 360: 2355-2372.
Direct Link