
Sawsan Abuhamdah
College of Pharmacy, Al Ain University of Science and Technology, Al Ain-Abu Dhabi, United Arab Emirates
LiveDNA: 962.16138
Duaa Thalji
Faculty of Pharmacy, Al-Isra University, Amman, Jordan
Naji Abuirmeileh
Department of Nutritional Sciences, Jordan University of Science and Technology, Irbid, Jordan
Anas Bahnassi
Coram CVS Specialty Infusion Services, Coram Health Services, Denver, CO, United State of America
Izzedin Salahat
Department of Medicinal Chemistry, Faculty of Pharmacy, University of Tabuk, Tabuk, Kingdom of Saudi Arabia
Amjad Abuirmeileh
Faculty of Pharmacy, Al-Isra University, Amman, Jordan
International Journal of Pharmacology
Year: 2017 | Volume: 13 | Issue: 6 | Page No.: 573-582







ABSTRACT
Background and Objective: Alzheimer’s disease (AD) is a progressive neurodegenerative disease where symptoms appear after significant neuronal loss. Current treatments for AD do not significantly alter the disease process, where they temporarily improve symptoms. In vitro data has indicated that low doses of vanillin reduce beta-amyloid aggregation and may possess cytoprotective effects. This research aimed to investigate the effect of vanillin on the AlCl3 mouse model of AD and thus its possible usefulness in managing AD. Methodology: Male albino mice were divided into 8 groups of 10. Each group received different treatments of AlCl3, AlCl3+vanillin (30, 60 or 120 mg kg–1 day–1), vanillin alone and control. These experiments lasted for 30 days after which animals were subjected to behavioral and neurochemical assessment. Analysis was carried out using one way analysis of variance (ANOVA) with Dunnett’s post test. Results: Lower doses of vanillin led to improvements in behavioral and neurochemical deficits induced by AlCl3. Conversely, a high dose of vanillin caused an exaggeration in the behavioral and neurochemical deficits induced by AlCl3. Furthermore, highest dose vanillin treatment, on its own, inflicted behavioral and neurochemical deficits comparable to those caused by AlCl3. Conclusion: It is concluded that vanillin negatively impacted cholinergic neuronal survival. Any marked benefits with lower doses of vanillin were attributed to its ability to indirectly increase synaptic acetylcholine abundance through blocking its degradation.
PDF Abstract XML References Citation
Received: April 06, 2017;
Accepted: May 29, 2017;
Published: July 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Sawsan Abuhamdah, Duaa Thalji, Naji Abuirmeileh, Anas Bahnassi, Izzedin Salahat and Amjad Abuirmeileh, 2017. Behavioral and Neurochemical Alterations Induced by Vanillin in a Mouse Model of Alzheimer’s Disease. International Journal of Pharmacology, 13: 573-582.
DOI: 10.3923/ijp.2017.573.582
URL: https://scialert.net/abstract/?doi=ijp.2017.573.582
DOI: 10.3923/ijp.2017.573.582
URL: https://scialert.net/abstract/?doi=ijp.2017.573.582
INTRODUCTION
Alzheimer’s disease is the most common neurodegenerative disease, causing problems with memory, thinking and behavior. Symptoms usually develop slowly and get worse over time, becoming severe enough to impede daily tasks1. The AD is not a normal part of aging, although the greatest known risk factor is increasing age. The AD is characterized by loss of mainly cholinergic neurons and their synapses in the cerebral cortex and certain subcortical regions of the brain namely, the hippocampus. The disease is progressive and unfortunately symptoms become apparent after a significant number of these neurons arelost. Although the disease itself will not cause death, after eventual loss of all involved neurons, quality of life is also lost2.
Although the pathology of the disease is well understood, the cause of AD is not yet known. Like other common chronic diseases, AD develops as a result of multiple factors rather than one single cause where both environmental and genetic factors contribute to the pathogenesis of AD. This is one reason why till now there are no available treatments capable of reversing or even stopping the progression of the disease. All current treatments only improve symptoms and at times may delay progression of the disease3.
Vanillin (4-hydroxy-3-methoxybenzaldehyde) is one of the most widely used flavoring agents in the world, it is extracted from the orchid Vanilla planifolia and Vanilla pompona family (Orchidaceae). It is a white needle-likes crystalline powder with an intensely sweet and very distinct pleasant odor4. In addition to its use as a flavoring agent, vanillin has been reported to exhibit multifunctional effect such as antimicrobial5, antioxidant6-8, anti-mutagenic9, antiangiogenetic10-12 and analgesic effects13,14. Furthermore, vanillin showed anti-depressive effect in tail suspension test as well as forced swim test15. Vanillin also demonstrated neuroprotection in an experimental model Huntington’s disease16. In another study, Kim et al.17 found that following global ischemia in gerbils, vanillin demonstrated a neuroprotective effect to hippocampal neurons. Other studies also demonstrated the cytoprotective effects of vanillin18,19. Also, in a new finding, vanillin ameliorated both motor and non-motor symptoms of Parkinson disease20. Recent evidence has indicated that vanillin reduces β-amyloid aggregation, one of the hallmark features of AD21. This phenomenon may be of value to patients suffering AD. Although this data has demonstrated these effects in vitro, they still have not been confirmed in vivo.
The present study was designed to investigate the behavioral and neurochemical effect of vanillin on the AlCl3 mouse model of AD, examining severity of the induced disease in the absence and presence of vanillin. Several studies indicated a positive effects of vanillin on neuronal survival16-21. This study aimed to explore these effects for the first time in an in vivo model of AD because it’s extensive human consumption, the results of this study implicated vanillin as a potential accomplice in the etiology of AD.
MATERIALS AND METHODS
Animals: Adult male albino mice (20-25 g) were purchased from Applied Science University and housed at Isra university animal care facility. The animal unit was maintained at a temperature of 21-22°C, with constant humidity and fixed dark:light cycles. The animals were fed with standard pellet diet and water ad libitum. All procedures concerning animals were carried out in accordance with Jordanian regulations for animal experimentation and care and were approved by the committee of institutional animal care and use (Protocol and Ethical approval memo number IU/FP/120 dated 7th of January, 2014). The study commenced on the (21st of March, 2014 and lasted for duration of 11 months). All experimentation was carried out in the Pharmacology Research Laboratory at the Faculty of Pharmacy at Isra University, Amman, Jordan.
Chemicals and reagents: 1-octane sulfonic acid sodium salt was purchased from (Santa Cruz Biotechnology, Inc, USA), di-sodium hydrogen phosphate anhydrous, aluminum chloride hexahydrate, vanillin and Phosphate Buffer Saline (PBS) were obtained from AZ Chem, acetylcholine chloride was from (Sigma Chemical Co., USA), perchloric acid was from fischer and HPLC water was provided from (LabChemInc, USA).
Experimental design
Administration of AlCl3 and vanillin: The AlCl3 treated groups were subjected to (300 mg kg–1 day–1 of AlCl3) in their drinking water for 30 days according to Walton22. Vanillin treated group were subjected to (30, 60, or 120 mg kg–1 day–1) once daily by oral gavage. Vanillin doses were chosen according to preliminary data which indicated that (30 mg kg–1 day–1) vanillin did improve behavioral deficits in AlCl3 treated mice. Vanillin doses of 60 and 120 mg kg–1 day–1 were used to evaluate any further behavioral improvement. Dose for preliminary trials was chosen according to unpublished data which indicate that vanillin decreased β-amyloid aggregation21.
Eighty animals were randomized and divided into the following 8 groups (n = 10). Group 1 was the control group and received only vehicle, group 2 received only (AlCl3 300 mg kg–1 day–1) for 30 days, groups 3, 4 and 5 received vanillin alone in doses of either 30, 60 or 120 mg kg–1 day–1 for 30 days. Groups 6, 7 and 8 received AlCl3 (300 mg kg–1 day–1) in addition to one of the doses of vanillin (30, 60 or 120 mg kg–1 day–1) for 30 days.
Tissue handling (Brains): After completion of animal treatments and behavioral assessment all animals were sacrificed by decapitation and the brains were removed immediately from the skull and placed on filter paper saturated with (0.2 M reagent grade sucrose) solution in a petri dish over crushed ice. Brains were then halved sagittally and the olfactory bulbs and cerebellum were removed. After this the hippocampi and cortices were dissected out and individually homogenized in 1 mL 0.1 M PBS using a pestle homogenizer from (Genomixbiotec, USA). Homogenates were centrifuged to remove insoluble material (10,000 rpm for 10 min at 5°C). About 40 μL of the supernatant was mixed with 10 μL of 0.2 M perchloric acid to precipitate proteins, then centrifuged again at (10,000 rpm for 10 min at 5°C). Supernatants were placed in sample vials and whole tissue Ach level were estimated using high performance liquid chromatography with cooled auto sampler and coupled to a photodiode array detector (HPLC-PDA, Thermo Scientific™ Dionex™ UltiMate™ 3000 VWD, USA).
HPLC analysis of Ach: Brain tissue levels of Ach were determined by HPLC-PDA at a flow rate of 0.5 mL min–1 by the method of Van Zoonen et al.23. In this method, acetylcholine is converted to choline by acetyl cholineesterase. Choline is then oxidized by choline oxidase, which is packed in to a column immobilized enzyme reactor (IMER), into hydrogen peroxide (H2O2). The H2O2 is then quantified by means of PDA at 240 nm23. Results were expressed in ng mg–1 weight of brain tissue.
Assessment of treatment groups: All experiment mice were assessed both behaviorally and chemically.
Behavioral assessment
Assessment of Spatial Reference Memory (SRM): Morris Water Maze (MWM) was employed to study mechanisms of spatial learning and memory24. The maze consisted of a circular pool (1.2 m in diameter and 0.47 m high) made of black plastic25,26. The pool was filled to depth of 20 cm with water (24-25° C). An escape platform (10 cm diameter), made of black plastic with a grooved surface for a better grip, was submerged 0.5 cm under the water where mice ought to swim until they found the hidden platform where mice used cues outside the maze to develop a spatial map of the environment and guide its performance. The pool was divided into four equal quadrants that were labeled and randomized daily. The time each mouse needed to find the hidden platform (latency) was recorded. During SRM training, the platform was kept in the same spatial location of the pool and a trial was started by placing the mouse in to the pool close to the rim, facing the wall of the pool and at different start positions across trials. The mice were given 4 days of training with four 60 sec training trials per day. At day 5 the platform was removed, then the time the mouse needed to reach the hidden platform and the time spent in the required quadrant after platform was removed were recorded27. After each session the apparatus was thoroughly cleaned with alcohol and dried.
Assessment of Spatial Working Memory (SWM): During spatial working memory training, the escape platform was placed near the edge of the pool in one of the four possible locations (designated N, S, E and W). This test consisted of 3 sessions with a 1 h intersession interval. In each session the platform location was changed from one quadrant to another. In each session the mice were trained twice (1 min for each training) to find the hidden platform. After the training a third test run was carried out and time required to locate platform was recorded26,27.
Neurochemical assessment: Extent of neuronal damage can be assessed neurochemically through measurement of available neurotransmitters, where their abundance can be considered representative of the number of surviving neurons. After behavioral analysis, the hippocampal and cortical regions of the brain for the control and experimental groups were assessed for the availability of Ach and its break down product choline through HPLC-PDA.
Assessment of total protein: Total protein content was determined spectrophotometrically at 550 nm (UV-Shimadzu, Japan) according to the Biuret assay described by George and O’Neil28 using Bovine Serum Albumin (BSA) as a standard.
Statistical analysis: The significance of differences between groups was determined using one-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparison test29, where p<0.05 was considered statistically significant. All the statistical analysis was performed using GraphPad Prism version 5.00 for Windows.
RESULTS
Behavioral assessment of spatial reference Memory and Working Memory using MWM: The MWM is a test of spatial learning that relies on distal cues to navigate from start locations around the perimeter of an open swimming arena to locate a submerged escape platform. Spatial learning is assessed across repeated trials and reference memory is determined by measuring latency in finding a hidden platform and by preference for the platform area when the platform is missing. Spatial learning was assessed in term of SRM as a function of time spent in a designated quadrant (Fig. 1a) and time required to reach the platform (Fig. 1b). The results clearly indicated that (AlCl3 alone, vanillin alone (120 mg kg–1 day–1) and AlCl3+vanillin 120 mg kg–1 day–1) treatment groups exhibited significant depletion (p<0.05) in SRM as seen in the reduced time spent in designated quadrant (Fig. 1a) and demonstrated significant increase (p<0.05) in time required to reach platform (Fig. 1b) while groups treated with AlCl3 and vanillin doses of (30 and 60 mg kg–1 day–1) demonstrated no significant depletion in SRM.
To assess working memory, the platform was relocated every day and the animal was given two trials (or more) per day where the animal must learn the new location of the platform by trial-and-error. As the platform is moved daily, no learning of platform position from the previous day can be transferred to the next day’s problem; hence, recall on each day during trial dependent on that day’s sample trial and measures only temporary or working memory.
The results clearly indicate that (AlCl3 alone, vanillin alone (120 mg kg–1 day–1) and AlCl3+vanillin 120 mg kg–1 day–1) treatment groups suffered impairment of spatial working memory as compared with control, vanillin alone (30 and 60 mg kg–1 day–1 doses) and AlCl3+vanillin (30 and 60 mg kg–1 day–1 doses) treated groups, where mice illustrated a significant increase (p<0.05) in average time needed to reach the desired quadrant for each mice group), where the place of the platform was changed in each session (Fig. 2).
Neurochemical assessment (measurement of tissue Ach): When examining Ach levels in relevant brain structures (hippocampus and cortex), it became evident that AlCl3 brought about significant reduction (p<0.05) in Ach and its degradation byproduct choline.
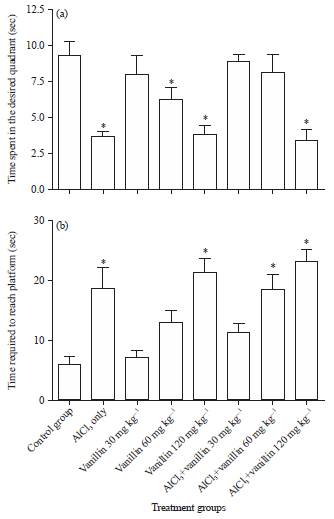 | |
| Fig. 1(a-b): | (a) Difference in SRM between groups as a function of the time spent in the designated quadrant after the platform was removed and (b) Difference in SRM between groups as a function of the time required to find hidden platform |
Control group was only treated with vehicle and all AlCl3 treated groups were subjected to 300 mg kg–1 day–1 for 30 days. Vanillin was administered in 3 different doses (30, 60 and 120 mg kg–1 day–1) from the beginning of the experiment and for the duration of the study either alone or with AlCl3. Data were expressed as Mean±SEM. *Indicates a significant difference from control (p<0.05) | |
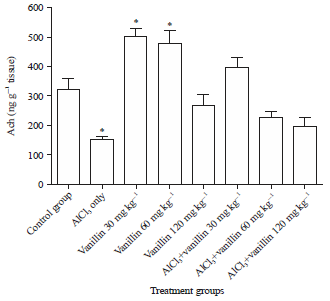
Interestingly, higher doses of vanillin (60 and 120 mg kg–1 day–1) also demonstrated significant depletion (p<0.05) in total Ach levels (Fig. 3-5 and 6-8 in the hippocampus and cortex respectively), this depletion was further exaggerated when these doses were used concomitantly with AlCl3. Conversely, 30 mg kg–1 day–1 of vanillin did not seem to present any significant reduction (p<0.05) in total Ach levels.
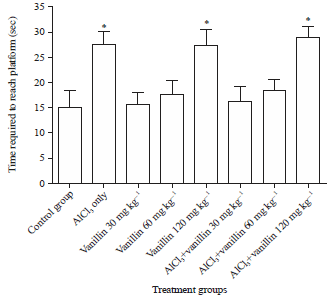 | |
| Fig. 2: | Difference in spatial working memory between groups as a function of the time required to reach platform |
The time represented in seconds on the y axis indicates the average time required to reach the platform from the 3 sessions. Control group was only treated with vehicle and all AlCl3 treated groups were subjected to 300 mg kg–1 day–1 for 30 days. Control group was only treated with vehicle and all AlCl3 treated groups were subjected to 300 mg kg–1 day–1 for 30 days. Vanillin was administered in 3 different doses (30, 60 and 120 mg kg–1 day–1) from the beginning of the experiment and for the duration of the study either alone or with AlCl3. Data were expressed as Mean±SEM. *Indicates a significant difference from control (p<0.05) | |
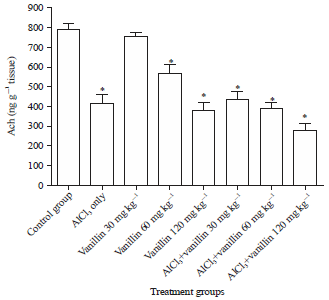 | |
| Fig. 3: | Difference in total hippocampal Ach levels between treatments groups |
Total Ach was reflected by adding measured intact Ach levels to measured choline levels. Control group was only treated with vehicle and all AlCl3 treated groups were subjected to 300 mg kg–1 day–1 for 30 days. Vanillin was administered in 3 different doses (30, 60 and 120 mg kg–1 day–1) from the beginning of the experiment and for the duration of the study either alone or with AlCl3. Data were expressed as Mean±SEM. *Indicates a significance from control difference (p<0.05) | |
 | |
| Fig. 4: | Difference in hippocampal Ach levels between treatment groups |
Control group was only treated with vehicle and all AlCl3 treated groups were subjected to 300 mg kg–1 day–1 for 30 days. Vanillin was administered in 3 different doses (30, 60 and 120 mg kg–1 day–1) from the beginning of the experiment and for the duration of the study either alone or with AlCl3. Data were expressed as Mean±SEM. *Indicates a significance from control difference (p<0.05) | |
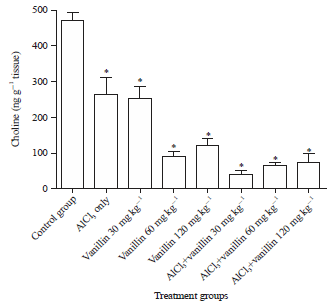 | |
| Fig. 5: | Difference in hippocampal choline levels between treatment groups |
Control group was only treated with vehicle and all AlCl3 treated groups were subjected to 300 mg kg–1 day–1 for 30 days. Vanillin was administered in 3 different doses (30, 60 and 120 mg kg–1 day–1) from the beginning of the experiment and for the duration of the study either alone or with AlCl3. Data were expressed as Mean±SEM. *Indicates a significance from control difference (p<0.05) | |
Further examination of total Ach levels indicated that intact Ach and choline levels were not constantly proportional to total Ach levels, where results (Fig. 3-8) indicated that vanillin preserved intact Ach and consequently decreased its degradation product choline.
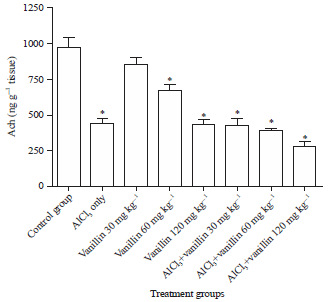 | |
| Fig. 6: | Difference in total cortical Ach levels between treatments groups |
Total Ach was reflected by adding measured intact Ach levels to measured choline levels. Control group was only treated with vehicle and all AlCl3 treated groups were subjected to 300 mg kg–1 day‾1 for 30 days. Vanillin was administered in 3 different doses (30, 60 and 120 mg kg–1 day–1) from the beginning of the experiment and for the duration of the study either alone or with AlCl3. Data were expressed as Mean±SEM. *Indicates a significance from control difference (p<0.05) | |
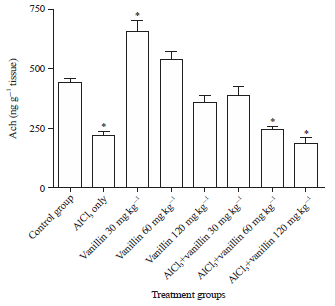 | |
| Fig. 7: | Difference in cortical Ach levels between treatment groups |
Control group was only treated with vehicle and all AlCl3 treated groups were subjected to 300 mg kg–1 day–1 for 30 days. Vanillin was administered in 3 different doses (30, 60 and 120 mg kg–1 day–1) from the beginning of the experiment and for the duration of the study either alone or with AlCl3. Data were expressed as Mean±SEM. *Indicates a significance from control difference (p<0.05) | |
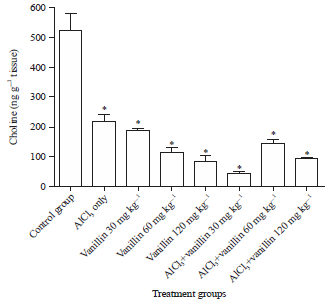 | |
| Fig. 8: | Difference in cortical choline levels between treatment groups |
Control group was only treated with vehicle and all AlCl3 treated groups were subjected to 300 mg kg–1 day–1 for 30 days. Vanillin was administered in 3 different doses (30, 60 and 120 mg kg–1 day–1) from the beginning of the experiment and for the duration of the study either alone or with AlCl3. Data were expressed as Mean±SEM. *Indicates a significance from control difference (p<0.05) | |
This preservation was apparent with every vanillin dose used and at times was capable of masking any deficit in total Ach levels.
DISCUSSION
This research demonstrated the effect of vanillin on the AlCl3 mouse model of AD. These effects were evaluated in term of vanillin’s ability to alter behavioral and neurochemical deficits induced by AlCl3. Although the effect of vanillin had never been evaluated in such a model, previous study demonstrated its possible usefulness in the management of AD19. Therefore, in this study there was an initial inclination to view results in terms of whether vanillin demonstrated any significant improvement (p<0.05) to this AD model or not. However as was demonstrated from the results, vanillin displayed behaviors that were contrary to earlier notions.
Preliminary behavioral data indicated that vanillin significantly improved (p<0.05) spatial reference memory where doses of (30 mg kg–1 day–1) for the duration of the trial seemed to counteract the negative effects inflicted by AlCl3 treatment. Long-term administration of AlCl3 to mice causes memory deficits and worsens their learning ability as demonstrated from this study and previous study30 where AlCl3 treatment inflicted depletion in SRM and SWM31. The ability of vanillin to reverse such effects could have been attributed to either a pharmacokinetic or a pharmacodynamics interaction with AlCl3. Because the nature of AlCl3 treatment requires it to be administered in the drinking water and vanillin was administered orally once daily, a pharmacokinetic interaction hindering AlCl3 absorption into the systemic circulation seems unlikely. Other pharmacokinetic interactions involving distribution, metabolism, or excretion also seem improbable due to the different physicochemical properties of the AlCl3 and vanillin.
When considering a possible pharmacodynamic interaction on membrane receptor level, again this would seem unlikely as AlCl3 possesses intracellular targets rather than extracellular ones. Still, possible biochemical interference in the cellular toxicity of AlCl3 by vanillin might be taking place. Potential mechanisms of interference may include vanillin’s ability to reduce Aβ aggregation19,32 caused by chronic AlCl3 treatment33. Another potential mechanism is vanillin’s ability to modulate Ach transmission by inhibiting the response of gamma-aminobutyric acid (GABA) receptors34. Also, Gupta and Sharma16 demonstrated that vanillin was capable of improving learning and memory impairment attributable to cholinergic dysfunction induced chemically through inhibition of mitochondrial respiratory complex II.
It has been demonstrated that AlCl3causes genomic instability and cell death. This was caused by the inhibition of DNA repair enzymes, enhancement of ROS production, antioxidant enzyme de-activation and decreasing RNA polymerase activity35. Any of these mentioned causes of AlCl3 effects may also pose as a possible target for vanillin.
When choosing to increase the doses of vanillin to (60 and 120 mg kg–1 day–1), behavioral and neurochemical data confirmed and conflicted results obtained from preliminary experiments. Where (60 mg kg–1 day–1) of vanillin seemed to improve AlCl3 induced deficits in spatial working memory and also increased cortical and hippocampal Ach levels that were reduced by AlCl3. On the other hand, the higher dose (120 mg kg–1 day–1) of vanillin clearly demonstrated the opposite behavioral and neurochemical effects. Here, this highest dose of vanillin inflicted the greatest AlCl3 induced deficits in terms of working memory and tissue Ach levels. Interestingly, when vanillin was used alone in this high dose, it inflicted behavioral and neurochemical deficits that were comparable to those induced by AlCl3. These conflicting results indicate that vanillin possesses a dose dependent effect where doses of (30 and 60 mg kg–1 day–1) showed improvements and doses of (120 mg kg–1 day–1) showed deterioration.
When taking a closer look at hippocampal and cortical levels of Ach, it was clearly evident that (60 mg kg–1 day–1) of vanillin was causing an increase in both cortical and hippocampal Ach levels, a decrease in hippocampal and cortical choline levels and a decrease in total Ach levels. This strongly suggested that vanillin actually induces Ach impairment at doses of (60 and 120 mg kg–1 day–1) but that the Ach impairment caused by 60 mg kg–1 day–1 vanillin was masked by this dose’s ability to prevent Ach breakdown. This notion is supported by Kundu et al.36 where vanillin was reported to have AchE inhibitory activity. About 120 mg kg–1 day–1 of vanillin, however, did not present such effects. At this juncture, it should be emphasized that when evaluating total Ach tissue levels, any deficits from control are presumed to reflect a reduction in cholinergic neuron numbers. On the other hand, choline levels were representative of natural Ach breakdown as can be seen in control groups. And where (60 mg kg–1 day–1) of vanillin reduced this natural breakdown enough to mask vanillin induced cell loss, the (120 mg kg–1 day–1) dose of vanillin did not achieve this. This could be attributed to a more extensive cell loss caused by the highest employed dose of vanillin which couldn’t be masked by any AchE inhibition taking place.
Possible reasons behind vanillin’s destructive effects could be through the inhibition of DNA repair enzymes. Here, vanillin has been demonstrated to impair the repair of DNA damage, chromosomal aberrations and chromosomal mutations induced by alkylating agents19,37.
Jayant et al. 38 also presented vanillin’s dose dependant effect, where lower doses of vanillin conferred neuroprotection while higher doses conveyed a destructive effect. Here, neuroprotection was attributed to vanillin’s ability to selectively activate the TRPV1 channel38. Conversely, prolonged activation of this channel negatively impacted neurons through inducing apoptosis39.
Other research also indicated vanillin’s ability to induce neurogenesis and promote cell proliferation and neuroblast differentiation in hippocampal subregions40. Moreover, Murakami et al.41 demonstrated the ability of vanillin to inhibit cyclooxygenase-2 and thus rendering an anti inflammatory effect. This anti inflammatory effect was also displayed by vanillin through inhibiting ERK1/2, P38 and NF-KB signaling42. These anti inflammatory effects would support vanillin’s favorable effect on neuronal survival, where neuroinflammation is a significant contributor to neurodegenerative disease.
In contrast, recent research failed to support vanillin’s beneficial effects on the pathology of AD, where it showed no ability to inhibit Aβ aggregation, a key feature of the disease43.
Although the results obtained from this research did not match initial expectations for vanillin’s ability to improve cholinergic-neurondeficits, they did however demonstrate vanillin’s negative impact on neuronal survival. The results clearly demonstrated a dose dependent effect for vanillin, with an apparent beneficial effect in lower doses. Although intermediate doses indicate that vanillin possesses favorable effects through AchE inhibition, the overall product is a destructive one on cholinergic neurons.
In an attempt to relate the outcomes of this study to real life, it must be indicated that the amount of vanillin present in beverages or food stuff as a flavoring agent varies widely and the doses investigated in this study represent really high daily exposure levels. Accordingly, any daily dosage overlap between experimentally used vanillin and actual life consumption, although possible, seems unlikely over a long duration of time.
CONCLUSION AND FUTURE RECOMMENDATIONS
In conclusion, this study indicated the negative effect of vanillin on cholinergic neuronal survival. The observed benefits with lower doses of vanillin were attributed to vanillin ability to indirectly increase synaptic Ach abundance through inhibiting AchE. The potential benefit of lower vanillin doses requires further evaluation to decide its limited usefulness in improving symptoms of AD. The long-term consequences of such low doses, whether as potential treatment for AD or from dietary consumption, would also require assessment for any negative effect on neuronal survival.
SIGNIFICANCE STATEMENT
This study will help the researcher to recognize vanillin as a substance with potential neuronal degenerative effects that may play a role in the etiology of Alzheimer’s disease.
ACKNOWLEDGMENTS
This study was funded by Isra University and the University of Jordan (IU/RG/PH/2/2014). Thanks to Mr. Mohammad Abushkedim (Department of Applied Pharmaceutical Sciences, Faculty of Pharmacy, Isra University) for all of his help with the animals used in this research. This study has been carried out during a sabbatical leave granted to Dr. Sawsan Abuhamdah from the Department of Biopharmaceutics and Clinical Pharmacy at the University of Jordan during the academic year 2016-2017.
REFERENCES
- Brookmeyer, R., E. Johnson, K. Ziegler-Graham and H.M. Arrighi, 2007. Forecasting the global burden of Alzheimer’s disease. Alzheimer's Dementia, 3: 186-191.
CrossRefDirect Link - Alzheimer's Association, 2013. Alzheimer's disease facts and figures. Alzheimer’s Dementia, 9: 208-245.
CrossRefDirect Link - Kim, H.G. and M.S. Oh, 2012. Herbal medicines for the prevention and treatment of Alzheimer's disease. Curr. Pharm. Des., 18: 57-75.
CrossRefDirect Link - Gallage, N.J. and B.L. Moller, 2015. Vanillin-bioconversion and bioengineering of the most popular plant flavor and its de novo biosynthesis in the vanilla orchid. Mol. Plant, 8: 40-57.
CrossRefDirect Link - Fitzgerald, D.J., M. Stratford, M.J. Gasson, J. Ueckert, A. Bos and A. Narbad, 2004. Mode of antimicrobial action of vanillin against Escherichia coli, Lactobacillus plantarum and Listeria innocua. J. Applied Microbiol., 97: 104-113.
CrossRefDirect Link - Tai, A., T. Sawano, F. Yazama and H. Ito, 2011. Evaluation of antioxidant activity of vanillin by using multiple antioxidant assays Biochim. Biophys. Acta-Gen. Subj., 1810: 170-177.
CrossRefDirect Link - Kumar, S.S., K.I. Priyadharsini and K.B. Sainis, 2002. Free radical scavenging activity of vanillin and o-vanillin using 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical. Redox Rep., 7: 35-40.
CrossRefPubMedDirect Link - Shyamala, B.N., M.M. Naidu, G. Sulochanamma and P. Srinivas, 2007. Studies on the antioxidant activities of natural vanilla extract and its constituent compounds through in vitro models. J. Agric. Food Chem., 55: 7738-7743.
CrossRefDirect Link - Ohta, T., M. Watanabe, K. Watanabe, Y. Shirasu and T. Kada, 1986. Inhibitory effects of flavourings on mutagenesis induced by chemicals in bacteria. Food Chem. Toxicol., 24: 51-54.
CrossRefDirect Link - Imanishi, H., Y. Sasaki, K. Matsumoto, M. Watanabe, T. Ohta, Y. Shirasu and K. Tutikawa, 1990. Suppression of 6-TG-resistant mutations in V79 cells and recessive spot formations in mice by vanillin. Mutat. Res. Lett., 243: 151-158.
CrossRefDirect Link - Sasaki, Y., H. Imanishi, M. Watanabe, T. Ohta and Y. Shirasu, 1990. Suppressing effect of antimutagenic flavorings on chromosome aberrations induced by UV-light or X-rays in cultured Chinese hamster cells. Mutat. Res./Fund. Mol. Mech. Mutagen., 229: 1-10.
CrossRefDirect Link - Sasaki, Y.F., T. Ohta, H. Imanishi, M. Watanabe, K. Matsumoto, T. Kato and Y. Shirasu, 1990. Suppressing effects of vanillin, cinnamaldehyde and anisaldehyde on chromosome aberrations induced by X-rays in mice. Mutat. Res. Lett., 243: 299-302.
CrossRefDirect Link - Park, S.H., Y.B. Sim, S.M. Choi, Y.J. Seo, M.S. Kwon, J.K. Lee and H.W. Suh, 2009. Antinociceptive profiles and mechanisms of orally administered vanillin in the mice. Arch. Pharm. Res., 32: 1643-1649.
CrossRefDirect Link - Beaudry, F., A. Ross, P.P. Lema and P. Vachon, 2010. Pharmacokinetics of vanillin and its effects on mechanical hypersensitivity in a rat model of neuropathic pain. Phytother. Res., 24: 525-530.
CrossRefDirect Link - Xu, J., H. Xu, Y. Liu, H. He and G. Li, 2015. Vanillin-induced amelioration of depression-like behaviors in rats by modulating monoamine neurotransmitters in the brain. Psychiatry Res., 225: 509-514.
CrossRefDirect Link - Gupta, S. and B. Sharma, 2014. Pharmacological benefits of agomelatine and vanillin in experimental model of huntington's disease. Pharmacol. Biochem. Behav., 122: 122-135.
CrossRefPubMedDirect Link - Kim, H.J., I.K. Hwang and M.H. Won, 2007. Vanillin, 4-hydroxybenzyl aldehyde and 4-hydroxybenzyl alcohol prevent hippocampal CA1 cell death following global ischemia. Brain Res., 1181: 130-141.
CrossRefDirect Link - Dhanalakshmi, C., T. Manivasagam, J. Nataraj, A.J. Thenmozhi and M.M. Essa, 2015. Neurosupportive role of Vanillin, a natural phenolic compound, on rotenone induced neurotoxicity in SH-SY5Y neuroblastoma cells. Evid. Based Complement. Alternat. Med.
CrossRefDirect Link - Keshava, C., N. Keshava, T.M. Ong and J. Nath, 1998. Protective effect of vanillin on radiation-induced micronuclei and chromosomal aberrations in V79 cells. Mutat. Res./Fund. Mol. Mech. Mutagen., 397: 149-159.
CrossRefDirect Link - Dhanalakshmi, C., U. Janakiraman, T. Manivasagam, A.J. Thenmozhi and M.M. Essa et al., 2016. Vanillin attenuated behavioural impairments, neurochemical deficts, oxidative stress and apoptosis against rotenone induced rat model of Parkinson’s disease. Neurochem. Res., 41: 1899-1910.
CrossRefDirect Link - Walton, J.R., 2006. Aluminum in hippocampal neurons from humans with Alzheimer's disease. Neurotoxicology, 27: 385-394.
CrossRefDirect Link - Van Zoonen, P., C. Gooijer, N.H. Velthorst, R.W. Frei, J.H. Wolf, J. Gerrits and F. Flentge, 1987. HPLC detection of choline and acetylcholine in serum and urine by an immobilized enzyme reactor followed by chemiluminescence detection. J. Pharm. Biomed. Anal., 5: 485-492.
CrossRefDirect Link - Brandeis, R. and S. Yehuda, 1989. The use of the morris water maze in the study of memory and learning. Int. J. Neurosci., 48: 29-69.
CrossRefDirect Link - Morris, R., 1984. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods, 11: 47-60.
CrossRefPubMedDirect Link - Van Der Staay, F.J. and M. de Jonge, 1993. Effects of age on water escape behavior and on repeated acquisition in rats. Behav. Neural Biol., 60: 33-41.
CrossRefDirect Link - George, J.W. and S.L. O’Neil, 2001. Comparison of refractometer and biuret methods for total protein measurement in body cavity fluids. Vet. Clin. Pathol., 30: 16-18.
CrossRefDirect Link - Dunnett, C.W., 1955. A multiple comparison procedure for comparing several treatments with a control. J. Am. Stat. Assoc., 50: 1096-1121.
CrossRefDirect Link - Berlyne, G.M., J.B. Ari, E. Knopf, R. Yagil, G. Weinberger and G.M. Danovitch, 1972. Aluminium toxicity in rats. Lancet, 299: 564-568.
CrossRefDirect Link - Song, S., X. Ma, Y. Zhou, M. Xu, S. Shuang and C. Dong, 2016. Studies on the interaction between vanillin and β-Amyloid protein via fluorescence spectroscopy and atomic force microscopy. Chem. Res. Chinese Univ., 32: 172-177.
CrossRefDirect Link - Aly, H.F., F.M. Metwally and H.H. Ahmed, 2011. Neuroprotective effects of dehydroepiandrosterone (DHEA) in rat model of Alzheimer's disease. Acta Biochem. Polinica, 58: 513-520.
PubMedDirect Link - Pajouhesh, H. and G.R. Lenz, 2005. Medicinal chemical properties of successful central nervous system drugs. NeuroRX, 2: 541-553.
CrossRef - Bharathi, P. Vasudevaraju, M. Govindaraju, A.P. Palanisamy, K. Sambamurti and K.S.J. Rao, 2008. Molecular toxicity of aluminium in relation to neurodegeneration. Indian J. Med. Res., 128: 545-556.
Direct Link - Kundu, A. and A. Mitra, 2013. Flavoring extracts of Hemidesmus indicus roots and Vanilla planifolia pods exhibit in vitro acetylcholinesterase inhibitory activities. Plant Foods Hum. Nutr., 68: 247-253.
CrossRefDirect Link - Aoshima, H. and Y. Tenpaku, 1997. Modulation of GABA receptors expressed in Xenopus oocytes by 13-l-hydoxylinoleic acid and food additives. Biosci. Biotechnol. Biochem., 61: 2051-2057.
CrossRefDirect Link - Jayant, S., B.M. Sharma and B. Sharma, 2016. Protective effect of transient receptor potential vanilloid subtype 1 (TRPV1) modulator, against behavioral, biochemical and structural damage in experimental models of Alzheimer's disease. Brain Res., 1642: 397-408.
CrossRefDirect Link - Song, J., J.H. Lee, S.H. Lee, K.A. Park, W.T. Lee and J.E. Lee, 2013. TRPV1 activation in primary cortical neurons induces calcium-dependent programmed cell death. Exp. Neurobiol., 22: 51-57.
CrossRefDirect Link - Cho, J.H., J.H. Park, J.H. Ahn, J.C. Lee and I.K. Hwang et al., 2016. Vanillin and 4-hydroxybenzyl alcohol promotes cell proliferation and neuroblast differentiation in the dentate gyrus of mice via the increase of brain-derived neurotrophic factor and tropomyosin-related kinase B. Mol. Med. Rep., 13: 2949-2956.
CrossRefDirect Link - Murakami, Y., A. Hirata, S. Ito, M. Shoji and S. Tanaka et al., 2007. Re-evaluation of cyclooxygenase-2-inhibiting activity of vanillin and guaiacol in macrophages stimulated with lipopolysaccharide. Anticancer Res., 27: 801-807.
Direct Link - Yan, X., D.F. Liu, X.Y. Zhang, D. Liu and S.Y. Xu et al., 2017. Vanillin protects dopaminergic neurons against inflammation-mediated cell death by inhibiting ERK1/2, P38 and the NF-κB signaling pathway. Int. J. Mol. Sci., Vol. 18.
CrossRefDirect Link - Espargaro, A., T. Ginex, M.D.M. Vadell, M.A. Busquets and J. Estelrich et al., 2017. Combined in vitro cell-based/in silico screening of naturally occurring flavonoids and phenolic compounds as potential anti-alzheimer drugs. J. Nat. Prod., 80: 278-289.
CrossRefDirect Link