
Amira Badr
Department of Pharmacology and Toxicology, College of Pharmacy, King Saud University, P.O. Box 22452, 11459 Riyadh, Saudi Arabia
Dalia Fouad
Department of Zoology and Entomology, Faculty of Science, Helwan University, EinHelwan, Cairo, Egypt
LiveDNA: 20.4569
International Journal of Pharmacology
Year: 2016 | Volume: 12 | Issue: 7 | Page No.: 675-688







ABSTRACT
Background and Objective: Cisplatin is an effective chemotherapeutic agent, while its usage is limited due to its nephrotoxicity. Olive Leaf Extract (OLE) possesses anti-inflammatory and antioxidant effects. This study aimed to investigate if OLE can protect against cisplatin-nephrotoxicity. Materials and Methods: Six groups were used, group 1: Control, group 2: Received 1000 mg kg–1 b.wt., OLE daily for 10 days via oral tubes, group 3: Received a single dose of cisplatin (7 mg kg–1 b.wt.) intraperitoneally on the 5th day and group 4-6: Received 250, 500 and 1000 mg kg–1 b.wt., OLE daily for 10 days, respectively and a cisplatin intraperitoneal injection (7 mg kg–1 b.wt.) on the 5th day. The ratio of kidney weight to body weight was calculated. Renal function tests, histopathology, level of renal malondialdehyde (MDA) and glutathione, immunohistochemical detection of NF-κB and cytochrome c, the renal level of TNF-α, caspase-9 and DNA fragmentation were determined. Results: Cisplatin induced a significant elevation of kidney weight to body weight ratio, serum Blood Urea Nitrogen (BUN) and creatinine, MDA, TNF-α and caspase-9 in kidney. Cisplatin increased the NF-κB activation, cytochrome C expression and DNA fragmentation were also detected. However, cisplatin decreased serum albumin and renal glutathione content. The OLE pre-treatment significantly ameliorated cisplatin nephrotoxicity with the highest dose nearly normalized all measured parameters. Conclusion: The OLE is a promising nephroprotective agent against cisplatin nephrotoxicity possibly through antioxidant, anti-inflammatory and anti-apoptotic activity.
PDF Abstract XML References Citation
How to cite this article
Amira Badr and Dalia Fouad, 2016. Anti-apoptotic and Anti-inflammatory Effects of Olive Leaf Extract Against Cisplatin-induced Nephrotoxicity in Male Rats. International Journal of Pharmacology, 12: 675-688.
DOI: 10.3923/ijp.2016.675.688
URL: https://scialert.net/abstract/?doi=ijp.2016.675.688
DOI: 10.3923/ijp.2016.675.688
URL: https://scialert.net/abstract/?doi=ijp.2016.675.688
INTRODUCTION
Human being suffer from several diseases, most of them have a treatment. Unfortunately, some medicals have side effects that limit the clinical application of these drugs. Recently, several studies have been done to explore the role and the possible use of natural products and dietary components as potential chemopreventive agents, to minimize the side effects of chemotherapeutic such as, curcumine1, ginseng2, thymoquinone3 and berberine4.
Cisplatin (CP) is one of the most widely used and most potent chemotherapeutic drugs. It is considered as one of the most remarkable successes in the war on cancer. Since the accidental discovery over four decades ago, CP has been widely used for chemotherapy5. It is used for the treatment of a variety of human cancers types, such as head, neck, colon, testis, breast, ovary, cervix, lung, bladder and many other tissues6,7. It most probably acts by binding to DNA, leading to the formation of inter and intrastrand cross-links resulting in defective DNA templates and arrest of DNA synthesis and replication8.
The cytotoxic activity of CP targets not only tumor cells, but also targets normal somatic cells, particularly in kidneys. Kidneys are the main route of CP excretion and proximal tubule cells serve as a primarily site of CP accumulation9. Thus, it is not surprising that nephrotoxicity is the most serious dose-limiting side effect in CP chemotherapy today, with a severe renal dysfunction that occurs in 20-30% of patients4. Oxidative stress, inflammation, apoptosis, necrosis, activate several signal transduction pathways involving in cell apoptosis, mitochondrial damage, decreased ATPase activity and impaired cellular transport mechanisms are suggested as the major mechanisms of CP-induced nephrotoxicity10,11.
The CP accumulates in the human kidney cells, resulting in the enhanced production of Reactive Oxygen Species (ROS) and the decrease in the antioxidant enzymes12. During CP nephrotoxicity, oxidative stress via ROS has been implicated in renal injury under various pathological conditions13. The ROS are produced during normal metabolism or as a consequence of the response to abnormal stress. It has been implicated in the pathogenesis of ageing and diseases including cancer, atherosclerosis, liver, lungs and kidney injury14,15. The ROS initiate inflammation and mediate the activation of NF-κB which upregulates the expression of several genes involved in the inflammatory response16. It may be one of the early signals that are at least partially responsible for the activation of various signaling pathways that culminate in renal cell injury and death during CP nephrotoxicity. Thus, the antioxidant activity or the inhibition of the generation of free radicals is important. Some of the antioxidants are naturally present in the body, while others have to be provided as supplements. Hence, their administration in the form of supplements may augment the function of endogenous free radical scavengers and consequently decrease the deleterious effects of the reactive metabolites12. Several antioxidants have shown its protective effect against renal dysfunction and tissue damage in CP-treated mice or rats17.
Moreover, previous studies showed that CP treatment stimulated the expression of proinflammatory chemokines and cytokines such as interleukins (IL-1β, IL-18 and IL-6) and tumor necrosis factor-α (TNF-α)2. The production of these inflammatory agents plays an important role in the pathogenesis of CP-induced renal injury and damage regulator18. The TNF-α appears to be a key upstream. Binding of TNF-α to the TNF-α death receptors (TNFR) 1 and 2 leads to the recruitment and activation of caspase-8, which further activates downstream caspases to induce apoptosis via extrinsic pathway. On the other hand, the intrinsic or mitochondrial pathway has emerged as the major apoptotic pathway in CP nephrotoxicity. The activation of NF-κB upregulates the production of TNF-α and the expression of the apoptotic genes4.
In mediterranean area and some other countries around the world, olives and its industrial derivatives were recommended traditionally as natural plants with health benefits because they are rich with polyphenolic compounds19. Extracts from the fruits and leaves of the olive tree (Olea europaea) are known to have antioxidant, anti-inflammatory, antiapoptotic properties and a cancer-preventive activities20-22. The majority of these properties are attributed to the presence of varying concentrations of phenolic compounds including oleuropein, verbascoside and ligstroside, which change gradually during maturation of the fruit to hydroxytyrosol and other simple phenolics23.
Olive Leaf Extract (OLE) and its active constituent, oleuropein has shown protective activity against toxicity of several compounds. It showed hepatoprotective activity in CCl4-treated mice24, cardioprotective activity in doxorubicin-treated cardiomyocytes25, neuroprotective and nephroprotective activity against gentamycin26 and cyclosporin induced nephrotoxicity27. This study aimed to evaluate the role of OLE as a promising protective remedy against the CP-induced nephrotoxicity treatment.
MATERIALS AND METHODS
Fresh olive tree leaves were collected from Jordan. Plant material was identified and authenticated by botanical survey.
Preparation of olive leaves extract: Five hundred grams of olive tree leaves obtained from Jordan; washed, air-dried and ground with a mixer to a fine powder. Olive leaves were then consecutively macerated for three days in 2 L of methanol with shaking. The resulting extract was filtered daily and 2 L fresh methanol were added. The methanolic extract was found to be rich in terms of chemical constituents and therefore was selected for the experiment. Collected fractions were put together. The methanolic solution was evaporated by using rotary evaporator (Rotavapor) to obtain a semisolid mass of methanolic olive leaves extract. The extract was then stored in -20°C until used28.
Chemicals: The CP was obtained as commercially used vials; product of Hospira (USA). Ether, methanol, n-butanol, 1,1’,3,3’-tetramethoxypropane, potassium dihydrogen phosphate, reduced glutathione (GSH), Ellman’s reagent (5,5’-dithio-bis-(2-nitrobenzoic acid)), trichloroacetic acid (TCA) and thiobarbituric acid (TBA) were purchased from Sigma-Aldrich (St. Louis, MO). Urea, creatinine and albumin kit were obtained from Sigma-Aldrich (St. Louis, MO). Rat Tumor Necrosis Factor-alpha (TNF-α) and caspase-9 ELISA kits were purchased from Raybioech (Norcross, USA) and Cloud-Clone corp (Houston, USA), respectively. Immunohistochemistry (IHC) antibodies for NF-κB and cytochrome C were obtained from Thermo Scientisic (USA). The DNA extraction kit was obtained from Qiagene (Hilden, Germany). Other chemicals were of high analytical reagents or grade.
Experimental design: Adult healthy male Wistar rats (Sprague Dawley) weighing (150-200 g) were obtained from the animal farm at King Saud University, Saudi Arabia, Riyadh. Animals were housed as four per cage in standard polypropylene cages. The rats were acclimatized for 1 week before beginning the experiments. The animals were provided with water and standard basal diet ad libitum and were maintained under controlled conditions of temperature, humidity and light (12:12 h light:dark cycle). Handling of animals was in compliance with the guidelines for the care and use of animals for scientific purposes with ethical approval in 12-1-15 for animal trial according to King Saud University instructions.
Forty eight adult rats were randomly divided into 6 groups, 8 rats per group. The experiment was carried out for 10 days. Group 1 (Control) served as untreated control, received vehicle daily via oral tube. Group 2 (OLE) received an oral administration of 1000 mg kg–1 b.wt., OLE daily for 10 days via oral tube. Group 3 (CP) rats received vehicle daily and a cisplatin was injected intraperitonealy (7 mg kg–1 b.wt.) at 5th day. Group 4-6 OLE (250, 500 and 1000 mg kg–1 b.wt.)+CP received orally 250, 500 and 1000 mg kg–1 OLE daily for 10 days, respectively and were also treated with a single dose of CP intraperitoneal injection (7 mg kg–1 b.wt.) on the 5th day of the experiments 1 h prior to OLE dose.
The animals of all groups were sacrificed by fast decapitation on the 11th day, animals were anesthetized with ether and sacrificed on the 6th day after cisplatin administration. Blood samples were collected, allowed to stand for half an hour and then centrifuged at 3000 rpm for 15 min at 4°C to separate serum and stored at -80°C for the different biochemical assays. The kidney was immediately removed, dried and weighed. Part of the kidney tissue was transferred immediately in 10% phosphate buffered formaldehyde for histological and immunohistochemical studies. Second part was weighed and homogenized immediately to give 50% (w/v) homogenate in ice-cold medium containing 50 mM tris-HCl, pH 7.4, to be used for the various biochemical determinations. Third part stored at -80°C for DNA extraction.
Estimation of kidney weight to body weight ratio: Rats were weighed on last day of experiment, kidney of each rat was dried and weighed to estimate kidney weight to body weight ratio:
Kidney function tests: Serum levels of Blood Urea Nitrogen (BUN), creatinine and albumin were estimated using diagnostic spectrophotometric kits; according to the standard procedure to assess the renal failure.
Histopathological examination: The kidney samples were fixed with 10% phosphate buffered formaldehyde for 24 h and then washed in tap water. Serial dilutions of alcohol (methyl, ethyl and absolute ethyl) were used for dehydration. Specimens were cleared in xylene and embedded in paraffin at 56°C in a hot air oven for 24 h. Paraffin bees wax tissue blocks were prepared for sections of 5-6 μm thickness were cut using a slidge microtome. The tissue sections obtained were collected on glass slides, deparaffinized and stained with hematoxylin and eosin stains for histopathological examinations with a light microscope29.
Determination of oxidative stress markers: Twenty percent kidney homogenate was used for determination of lipid peroxidation as a marker of oxidative stress. Lipid peroxides were determined spectrophotometrically as thiobarbituric acid-reactive substances (TBARs) according to the method of Uchiyama and Mihara30. The colorimetric determination of TBARs based on the reaction of malondialdehyde (MDA) with thiobarbituric acid at low pH and at high temperature. The resultant pink colored product was extracted by n-butanol and the absorbance is determined spectrophotometrically at 535 nm.
Twenty percent kidney homogenate was used for determination of reduced glutathione (GSH). The assay procedure involves the estimation of GSH using spectrophotometer according to Ellman’s method31. An aliquot of 0.5 mL of tissue homogenate was used. Precipitation of protein thiols by TCA was carried out and samples were centrifuged at 3000 rpm for 10 min. The resulting supernatant was used for determination of GSH using Ellman’s reagent. The absorbance was measured at 412 nm.
Detection of TNF-α and caspase 9 using ELISA kits: Tumor Necrosis Factor-α (TNF-α) and caspase 9 (Raybioech and Cloud-Clone corp) were detected in kidney tissue homogenate samples using standard sandwich enzyme-linked immunosorbent assay (ELISA) kit according to the manufacturer’s instruction.
Total DNA preparation extraction and fragmentation: Thirty milligrams of each tissue was homogenized in RTL lysis buffer (Qiagen) containing 1% 2-mercaptoethanol. Total DNA was extracted using All Prep DNA/RNA Mini kit (Qiagen, Cat# 80204), following the manufacturers manual and the DNA was eluted with 50 μL elution buffer. Quantification of the extracted DNA was determined using NanoDrop-8000 and its integrity was assessed by agarose gel (1.5%) electrophoresis. Gels were illuminated with 300 nm UV light and a photographic record was made32.
Immunohistochemical detection of NF-κB and cytochrome c: Formalin-fixed tissues were embedded in paraffin and cut into 5 μm sections. The kidney sections were fixed overnight, deparaffinised in xylene, rehydrated in a graded series of ethanol and boiled in antigen unmasking solution (Vector Laboratories, Burlingame, CA, USA) for 5 min. Sections were immersed in peroxidase-blocking reagent (Dako, Botany Bay, NSW, Australia) for 10 min and incubated in a humidified chamber with blocking goat serum (Dako) for 30 min. Sections were incubated with anti-NF-κB antibody (rabbit polyclonal) (1:500) in blocking solution for 12 h at 4°C for NF-κb detection or cytochrome c antibody (Clone 7H8.2C1) (mouse monoclonal) for cytochrome c detection. They were then re-equilibrated to room temperature and washed with PBS, incubated with Horse Radish Peroxidase (HRP) antibody conjugates (1:2500) in blocking solution without tween-20 for 2 h at room temperature.
Specimens were washed with PBS and incubated with 0.2% solution of 3,3-diaminobenzidine (DAB) until desired stain intensity develops followed by washing in distilled water. Sections were counterstained with hematoxylin, dehydrated in a graded series of ethanol and mounted with di-n-butylphthalate-polystyrenexylene (DPX)33. Immunoreactivity was quantitated in a blinded manner by three independent observers and the total number of positively stained cells was quantitated.
Statistical analysis: Results are reported as mean±standard error of the mean (SEM). Statistical analysis was performed using one-way analysis of variance (ANOVA). If the overall p-value was found statistically significant (p<0.05), further comparisons among groups were made according to post hoc Tukey’s test. All statistical analyses were performed using Graph Pad In Stat 3 (Graph Pad Software, Inc. La Jolla, CA, USA) software. Graphs were sketched using Graph Pad Prism version 4 software (Graph Pad Software, Inc. La Jolla, CA, USA).
RESULTS
Kidney weight/body weight ratio: Kidney weight/body weight ratio of CP treated group significantly increased (p<0.01) compared to control group. The OLE pre-treatment with different concentrations administered with CP showed a significant decrease (p<0.05, p<0.01 and p<0.01, respectively) in the ratio as compared to CP administered to animals alone (Fig. 1a).
Levels of serum BUN, creatinine and albumin: The levels of serum BUN and creatinine were significantly higher in the CP-treated animals when compared to control group. Treatment with OLE+CP significantly decreased levels of BUN and creatinine at all different doses except at OLE (250 mg kg–1 b.wt.) regarding creatinine level (Fig. 1b, c). While, the levels of serum albumin was significantly lowered in the CP-treated animals when compared to control group, animals treated with OLE+CP maintained normal albumin level at all doses (Fig. 1d).
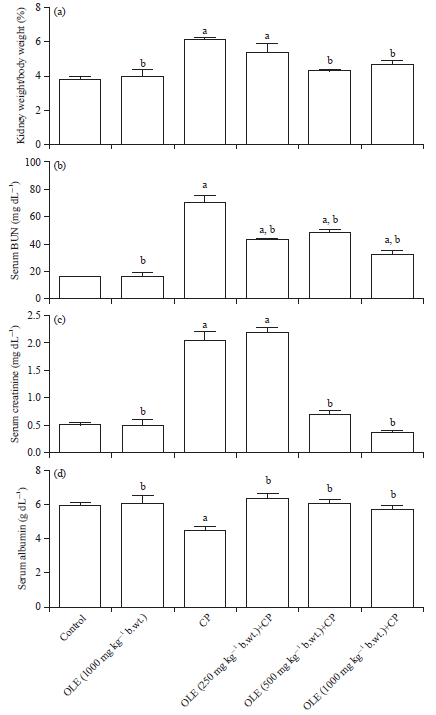 | |
| Fig. 1(a-d): | (a) Kidney weight/body weight ratio (%) in different animal groups, (b) Serum BUN (mg dL–1), (c) Serum creatinine (mg dL–1) and (d) Albumin (g dL–1) in different animal groups. Each value represents the mean of 8 records+SE, ap<0.05 significant change with respect to control group, bp<0.05 significant change with respect to CP group |
Treatment with OLE (1000 mg kg–1 b.wt.) alone did not reveal any changes in the levels of the tested parameters; as compared to control group (Fig. 1a-d).
Kidney histopathology: Control group (Fig. 2a) and rats treated with OLE (Fig. 2b) showed normal glomeruli and tubules and no significant differences were found between the two groups. The CP administration resulted in histopathologic changes of kidneys including cellular degeneration in lining epithelium of renal tubules and desquamation of degenerated cells present in the lumen of the tubules, vacuolization, apoptosis, sever necrosis, loss of brush border, blood congestion and debris of the intratubular and flattening of the tubular epithelium. It also caused tubular dilatation in the corticomedullary junction of rats kidneys. In necrotic tubules, eosinophilic amorphous material as well as pyknotic and karyorrhectic debris was observed (Fig. 2c).
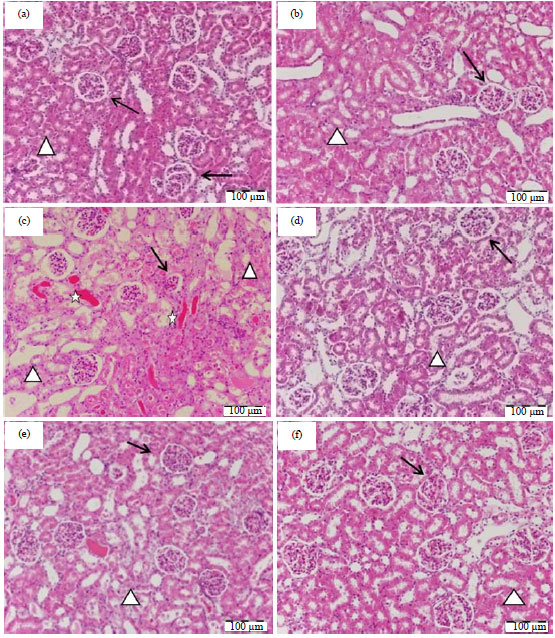 | |
| Fig. 2(a-f): | (a) Histological examinations by hematoxylin and eosin staining demonstrating the effect of OLE on CP induced renal damage in rats. Light micrographs showing group treated with vehicle, (b) OLE 1000 mg kg–1 b.wt., showing the normal glomruli (↑) and renal tubules lined by tubular cells with vesicular nuclei (Δ), (c) CP showing the necrotic glomeruli (↑), the renal tubules with marked vacuolation of its lining epithelium and dens nuclei (Δ) and (d-f) Congested blood vessels (✰), OLE (250, 500 and 1000 mg kg–1 b.wt.)+CP showing decreased renal damaged in a dose-dependent manner; evidenced by normal glomruli (↑) and renal tubules (Δ), that in rats receiving the highest dose of OLE was relatively resembling control group. Representative results from 8 similarly treated rats. Original magnification is 40X |
Treatment with OLE dose-dependently reduced pathological changes in the kidneys including extensive glomerular necrosis and degenerated cells (Fig. 2d, e), with visible mitosis and regeneration of tubular epithelium in group treated with OLE at the dose of 1000 mg kg–1 b.wt. (Fig. 2f). These results show that OLE administration protected kidney from renal injury after cisplatin injection. A significant improvement in kidney morphology and overall renal injury score was noticed (Fig. 2d-f).
Oxidative stress markers
Effect of CP and OLE treatment on LPO level: The CP treatment significantly enhanced (p<0.01) the LPO in kidney tissues as compared to control group, as evidenced by increased TBARS. Groups pre-treated with different concentrations of OLE with CP showed a significant decline (p<0.05, p<0.01 and p<0.01, respectively) in LPO level as compared to CP alone (Fig. 3a).
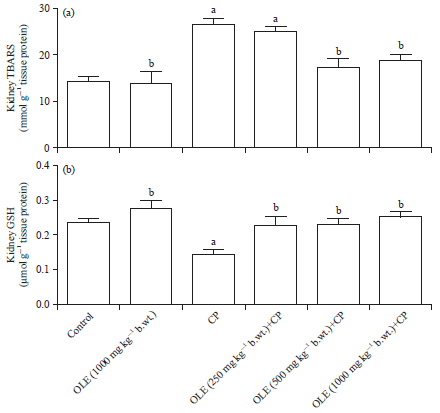 | |
| Fig. 3(a-b): | TBARS (nmol g–1 tissue protein) and kidney GSH (μmole g–1 tissue protein) in different animal groups. Each value represents the mean of 8 records+SE, ap<0.05 significant change with respect to control group, bp<0.05 significant change with respect to CP group |
Effect of CP and OLE treatment on GSH level: The CP treatment significantly (p<0.01) reduced the level of GSH as compared to control group (Fig. 3b). The OLE pretreated animals supplemented with CP showed a significant (p<0.01) maintenance of GSH level comparable to that of control group.
The OLE (1000 mg kg–1 b.wt.) alone did not reveal any changes in the levels of LPO and GSH as compared to control group (Fig. 3a, b).
Inflammatory response markers
Nuclear factor kappa B: Immunohistochemical analysis of NF-κB expression in the kidneys of control group (Fig. 4a) and rats treated with OLE (Fig. 4b) showed low NF-κB immunoreactivity. In contrast, strong NF-κB immunopositivity was found in renal tubular cells of CP-treated animals, with numerous positive nuclei (Fig. 4c). Treatment of CP-intoxicated rats with OLE reduced NF-κB immunopositivity in a dose-dependent manner (Fig. 4d-f).
Renal expression of NF-κB in rats receiving the highest dose of OLE was similar to control group (Fig. 4f), without nuclear immunopositivity.
Tumor necrosis factor-α: Figure 5 shows the level of cytokine TNF-α quantitation by ELISA were markedly increased (223% of control) in kidney of rats treated with cisplatin. On the other hand, OLE alone slightly decreased the levels of TNF-α compared to control. Rats treated with OLE at different doses prior to cisplatin exhibited a significant (p<0.01) suppression in the level of TNF-α compared to rats that treated with cisplatin alone (Fig. 5).
Apoptosis
Genomic DNA fragmentation: Figure 6 shows the qualitative changes in the integrity of the renal genomic DNA which indicated the apoptosis. Agarose gel electrophoresis shows that cisplatin treatment resulted in a dramatic oligonucleosome-length damage of DNA, characterized by smearing of DNA fragments. On the other hand, DNA isolated from control samples and OLE (1000 mg kg–1 b.wt.) alone exposed tissues showed no DNA smearing, suggesting no DNA fragmentation. The DNA of rats treated with OLE (1000 mg kg–1) prior to cisplatin showed improvement in the DNA damage with mild smearing.
Caspase-9 activity: The apoptosis induced in the cisplatin group correlated with the elevation in caspase-9 activity, which also known as index of apoptosis. While, the group treated with OLE prior cisplatin significantly reduced the caspase-9 activity (Fig. 7).
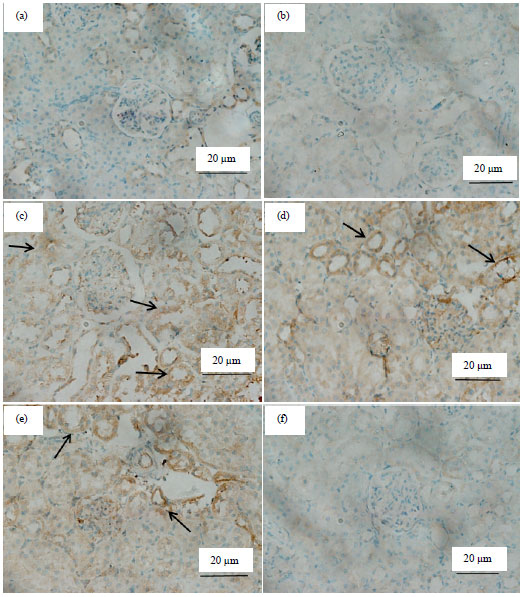 | |
| Fig. 4(a-f): | Immunohistochemistry was performed for NF-κB expression in rat kidneys in different animal groups. (a) Rats treated with vehicle, (b) OLE 1000 mg kg–1 b.wt., (c) CP and (d-f) OLE (250, 500 and 1000 mg kg–1 b.wt.)+CP. Arrows show NF-κB immunopositive nuclei. Representative results from 8 similarly treated rats. Immunohistochemistry staining, original magnification 40X |
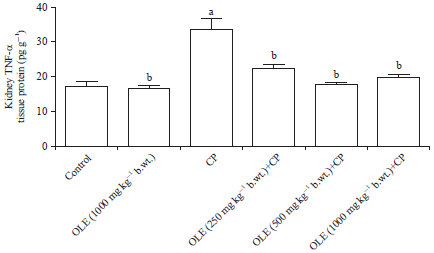 | |
| Fig. 5: | Kidney TNF-α ELISA (tissue protein pg g–1) in different animal groups. Each value represents the mean of 8 records+SD, ap<0.05 significant change with respect to control group, bp<0.05 significant change with respect to CP group |
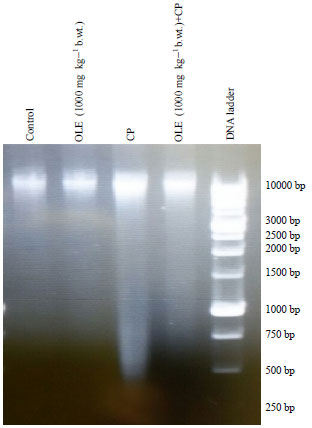 | |
| Fig. 6: | DNA fragmentation in control and experimental rats, Lanes, 1: Control group, 2: Group treated with OLE (1000 mg kg–1 b.wt.), 3: Group treated with CP and 4: Treated with OLE (OLE 1000 mg kg–1 b.wt.)+CP |
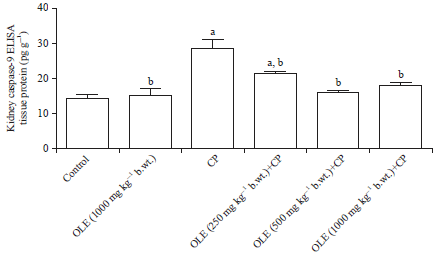 | |
| Fig. 7: | Caspase 9 activity ELISA (tissue protein pg g–1) in different animal groups. Each value represents the mean of 8 records+SD, ap<0.05 significant change with respect to control group, bp<0.05 significant change with respect to CP group |
DISCUSSION
The CP is an effectively important chemotherapeutic anticancer agent10. However, several studies have shown that its administration is associated with nephrotoxicity, which occurs through overproduction of Reactive Oxygen Species (ROS), tubule interstitial inflammation and apoptosis12,34. This study investigates the possible protective effect of OLE against cisplatin-induced nephrotoxicity in rats.
In the current study, the administration of a single dose of CP (7 mg kg–1 b.wt.) resulted in significant changes in kidney weight/body weight ratio and renal function tests. The blood urea nitrogen and serum creatinine were increased in the CP treated group which serum albumin was decreased due to loss of albumin in urine compared to control group (Fig. 1). Changes in kidney function tests may occur due to glomerular damage and decreased glomerular filtration rate35,36.
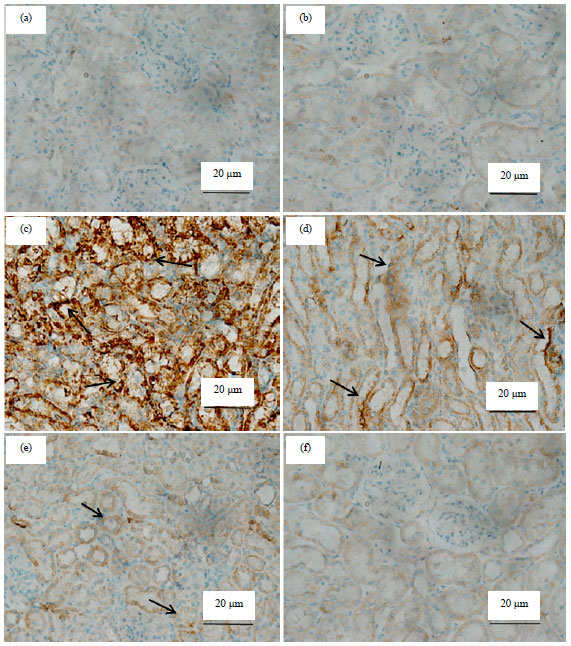 | |
| Fig. 8(a-f): | Immunohistochemistry was performed for expression of cytochrome c in the rat kidneys. (a) Rats treated with vehicle, (b) OLE 1000 mg kg–1 b.wt., (c) CP and (d-f) OLE (250, 500 and 1000 mg kg–1 b.wt.)+CP. Arrows show cytochrome c immunopositive expression. Representative results from 8 similarly treated rats. Immunohistochemistry staining, original magnification 40X |
Nephrotoxicity was also evident by these histopathological examination (Fig. 2c) that showed obvious tissue damage in kidney, including vacuolization, severe necrosis, degenerative changes in lining epithelium of renal tubules and desquamation of degenerated cells present in the lumen of the tubules. Pretreatment with different doses of (OLE) maintained the ratio of kidney weight/body weight and protected the kidney against cisplatin-induced nephrotoxicity, as evidenced by maintaining normal kidney function tests and histopathological examination showing preserved renal tissue integrity in a dose-dependent manner (Fig. 2d-f). No significant differences were found between the control and OLE treated group whose showed normal renal structure (Fig. 2a, b).
The degradation of cellular DNA by endonucleases is an important component of renal tubular epithelial cell death induced by ischemia or nephrotoxins37. Cisplatin binds to DNA to form covalent platinum DNA adducts and also acts as a DNA alkylator. In addition to the adduct-forming direct DNA interaction, cisplatin appears to induce ROS generation that leads to oxidative stress in several types of cells38. The excess of free radicals are directly involved in kidney injury39, the antioxidant system act as powerful protecting agents against oxidative stress which leads to acute renal failure. All these findings were consistent with the results obtained from the present study. The CP induced oxidative stress and affected antioxidant defense system as it increased the level of kidney malondialdehyde; the end product of the lipid peroxidation (Fig. 3a) and decreased the level of GSH (Fig. 3b) in rats treated with CP. The decrease of GSH may occur due to the formation of cisplatin-GSH conjugation products40. These results are in agreement with Chirino et al.41.
Pretreatment with OLE protected cells against CP-induced oxidative stress as it maintained GSH level comparable to that of control and decreased MDA levels significantly as compared to CP group. This can be attributed to antioxidant activity of OLE (Fig. 3b). Many researchers have demonstrated the ability of the OLE to scavenge free radicals due to the presence of several phenolic antioxidant compounds; including p-coumaric, ferulic and sinapic acids, flavonoids and procyanidins42,43.
Acute kidney injury is associated with a robust inflammatory response. It was found that inflammation plays a master role in the pathogenesis of cisplatin-induced nephrotoxicity5. The inflammatory process is a key contributor to acute kidney damage. In order to study the anti-inflammatory effect of OLE in this experimental model, the expression of NF-κB and the TNF-α level in renal tissues was determined. Both were significantly increased in cisplatin treated rats suggesting the implication of inflammation in mediating nephrotoxicity (Fig. 4, 5). These findings are consistent with14,44,45.
The TNF-α appears to be a key upstream regulator in the inflammatory response triggered by cisplatin. The TNF-α is a known proinflammatory cytokine involved in the inflammatory responses such as differentiation, maturation and activation of inflammatory cells such as neutrophils, T cells, macrophages and natural killer cells45. The TNF-α as a cytokine is increased in the kidney after CP intoxication which induces the inflammation and directs cellular toxicity10. Therefore, the reduction of TNF-α production results in lower CP-mediated nephrotoxicity44. The production of TNF-α after CP administration is considered as the direct result of ROS formation and activation of NF-κB. The TNF-α itself can activate IκB kinase, leading to NF-κB activity46. The NF-κB activation is pivotal in the expression of proinflammatory cytokines like TNF-α and other mediators involved in acute inflammatory responses and other conditions associated with increased ROS generation.
The toxicity and inflammation of CP in rat tissues were significantly ameliorated by OLE through the inhibition of NF-κB expression and TNF-α production (Fig. 4, 5). The ability of OLE to suppress the NF-κB pathway demonstrates its anti-inflammatory activity, which is in agreement with previous studies in different experimental models47.
Apoptosis plays an important role in the development and maintenance of homeostasis in most of the multicellular organisms. This type of cell death can be triggered by the activation of ROS, pro-inflammatory cytokines that ultimately culminate in the activation of caspase family of proteases. Apoptotic process induced by CP is through the release of cytochrome C and then generates superoxide48. Apoptosis play important role in the pathophysiology of high doses of CP-induced renal injury18,49. The mechanism of CP chemotherapeutic activity depends on the high doses of CP which induce apoptosis in response to cellular stresses and DNA damage50. The CP results in the apoptosis via the activation of p53 which induces the transcription of apoptotic genes, including caspase-dependent or independent pathways51,52.
In the present study, the increase in DNA fragmentation, caspase-9 activation and cytochrome c expression were observed in CP-treated rat kidneys (Fig. 6-8). It has been hypothesized that oxidative damage can occur in DNA during the peroxidative breakdown of membrane polyunsaturated fatty acids. The DNA damage affects homeostasis of various cells leading to induced signal transductions associated with apoptosis and cell proliferation53,54. Administration of OLE extracts prior to CP treated rats protected the renal tissues and markedly decreased the percentage of fragmented DNA (Fig. 6), through reduced all the previous apoptotic factors (Fig. 7, 8). These results agree with the most recent findings on the antiapoptotic effect of OLE55,56.
These results suggest that antioxidant activity and the decrease in inflammatory mediators and apoptotic factors expression could be the key mechanism for nephroprotective activity of OLE against CP-induced nephrotoxicity. These findings are in accordance with growing evidence on free radical scavenging properties of natural phenolic compounds57,58.
CONCLUSION
In conclusion, to the best of our knowledge, this is the first study to demonstrate the nephroprotective effect of OLE against kidney injury induced by CP.
ACKNOWLEDGMENTS
This research project was supported by a grant from the "Research Center of the Female Scientific and Medical Colleges", Deanship of Scientific Research, King Saud University.
REFERENCES
- Waseem, M., P. Pandey, B. Tomar, S. Raisuddin and S. Parvez, 2014. Ameliorative action of curcumin in cisplatin-mediated hepatotoxicity: An in vivo study in wistar rats. Arch. Med. Res., 45: 462-468.
CrossRefDirect Link - Yousef, M.I. and H.M. Hussien, 2015. Cisplatin-induced renal toxicity via tumor necrosis factor-α, interleukin 6, tumor suppressor P53, DNA damage, xanthine oxidase, histological changes, oxidative stress and nitric oxide in rats: Protective effect of ginseng. Food Chem. Toxicol., 78: 17-25.
CrossRefDirect Link - Al-Malki, A.L. and A.A.R. Sayed, 2014. Thymoquinone attenuates cisplatin-induced hepatotoxicity via nuclear factor kappa-β. BMC Complementary Altern. Med., Vol. 14.
CrossRefDirect Link - Domitrovic, R., O. Cvijanovic, E. Pernjak-Pugel, M. Skoda, L. Mikelic and Z. Crncevic-Orlic, 2013. Berberine exerts nephroprotective effect against cisplatin-induced kidney damage through inhibition of oxidative/nitrosative stress, inflammation, autophagy and apoptosis. Food Chem. Toxicol., 62: 397-406.
CrossRefDirect Link - Pabla, N. and Z. Dong, 2008. Cisplatin nephrotoxicity: Mechanisms and renoprotective strategies. Kidney Int., 73: 994-1007.
CrossRefDirect Link - Cepeda, V., M.A. Fuertes, J. Castilla, C. Alonso, C. Quevedo and J.M. Perez, 2007. Biochemical mechanisms of cisplatin cytotoxicity. Anticancer Agents Med. Chem., 7: 3-18.
PubMedDirect Link - Wang, D. and S.J. Lippard, 2005. Cellular processing of platinum anticancer drugs. Nat. Rev. Drug Discov., 4: 307-320.
CrossRefPubMedDirect Link - Hah, S.S., K.M. Stivers, R.W. de Vere White and P.T. Henderson, 2006. Kinetics of carboplatin-DNA binding in genomic DNA and bladder cancer cells as determined by accelerator mass spectrometry. Chem. Res. Toxicol., 19: 622-626.
CrossRefDirect Link - Yao, X., K. Ichpisal, N, Kurtzman and K. Nugent, 2007. Cisplatin nephrotoxicity: A review. Am. J. Med. Sci., 334: 115-124.
CrossRefPubMedDirect Link - Miller, R.P., R.K. Tadagavadi, G. Ramesh and W.B. Reeves, 2010. Mechanisms of cisplatin nephrotoxicity. Toxins, 2: 2490-2518.
CrossRefDirect Link - Podratz, J.L., A.M. Knight, L.E. Ta, N.P. Staff and J.M. Gass et al., 2011. Cisplatin induced mitochondrial DNA damage in dorsal root ganglion neurons. Neurobiol. Dis., 41: 661-668.
CrossRefDirect Link - Saad, A.A., M.I. Youssef and L.K. El-Shennawy, 2009. Cisplatin induced damage in kidney genomic DNA and nephrotoxicity in male rats: The protective effect of grape seed proanthocyanidin extract. Food Chem. Toxicol., 47: 1499-1506.
PubMed - Taguchi, T., A. Nazneen, M.R. Abid and M.D. Razzaque, 2005. Cisplatin-associated nephrotoxicity and pathological events. Contrib. Nephrol., 148: 107-121.
PubMedDirect Link - Liang, N. and D.D. Kitts, 2014. Antioxidant property of coffee components: Assessment of methods that define mechanisms of action. Molecules, 19: 19180-19208.
CrossRefDirect Link - Antosiewicz, J., W. Ziolkowski, S. Kar, A. A. Powolny and S.V. Singh, 2008. Role of reactive oxygen intermediates in cellular responses to dietary cancer chemopreventive agents. Planta Med., 74: 1570-1579.
CrossRefDirect Link - Singh, N., V. Kamath, K. Narasimhamurthy and P.S. Rajini, 2008. Protective effect of potato peel extract against carbon tetrachloride-induced liver injury in rats. Environ. Toxicol. Pharmacol., 26: 241-246.
CrossRefDirect Link - Takai, N., K. Abe, M. Tonomura, N. Imamoto and K. Fukumoto et al., 2015. Imaging of reactive oxygen species using [3H]hydromethidine in mice with cisplatin-induced nephrotoxicity. EJNMMI Res., Vol. 5.
CrossRefDirect Link - Faubel, S., E.C. Lewis, L. Reznikov, D. Ljubanovic and T.S. Hoke et al., 2007. Cisplatin-induced acute renal failure is associated with an increase in the cytokines interleukin (IL)-1β, IL-18, IL-6 and neutrophil infiltration in the kidney. J. Pharmacol. Exp. Ther., 322: 8-15.
CrossRefDirect Link - Dub, A.M. and A.M. Dugani, 2013. Antithrombotic effect of repeated doses of the ethanolic extract of local olive (Olea europaea L.) leaves in rabbits. Libyan J. Med., Vol. 8.
Direct Link - Masoko, P. and D.M. Makgapeetja, 2015. Antibacterial, antifungal and antioxidant activity of Olea africana against pathogenic yeast and nosocomial pathogens. BMC Complement. Altern. Med., Vol. 15.
CrossRefDirect Link - Andrikopoulos, N.K., A.C. Kaliora, A.N. Assimopoulou and V.P. Papageorgiou, 2002. Inhibitory activity of minor polyphenolic and nonpolyphenolic constituents of olive oil against in vitro low-density lipoprotein oxidation. J. Med. Food, 5: 1-7.
CrossRefDirect Link - Kaeidi, A., S. Esmaeili-Mahani, V. Sheibani, M. Abbasnejad, B. Rasoulian, Z. Hajializadeh and S. Afrazi, 2011. Olive (Olea europaea L.) leaf extract attenuates early diabetic neuropathic pain through prevention of high glucose-induced apoptosis: In vitro and in vivo studies. J. Ethnopharmacol., 136: 188-196.
CrossRefDirect Link - Boskou, G., F.N. Salta, S. Chrysostomou, A. Mylona, A. Chiou and N.K. Andrikopoulos, 2006. Antioxidant capacity and phenolic profile of table olives from the Greek market. Food Chem., 94: 558-564.
CrossRefDirect Link - Domitrović, R., H. Jakovac, V.V. Marchesi, I. Šain, Ž. Romić and D. Rahelić, 2012. Preventive and therapeutic effects of oleuropein against carbon tetrachloride-induced liver damage in mice. Pharmacol. Res., 65: 451-464.
CrossRefDirect Link - Andreadou, I., D. Benaki, P. Efentakis, S.I. Bibli and A.I. Milioni et al., 2015. The natural olive constituent oleuropein induces nutritional cardioprotection in normal and cholesterol-fed rabbits: Comparison with preconditioning. Planta Med., 81: 655-663.
CrossRefPubMedDirect Link - Tavafi, M., H. Ahmadvand and P. Toolabi, 2012. Inhibitory effect of olive leaf extract on gentamicin-induced nephrotoxicity in rats. Iran. J. Kidney Dis., 6: 25-32.
PubMedDirect Link - Mostafa-Hedeab, G., L.M. Sati, H.M. Elnaggar, Z.O. Elgatlawey, A.A. Eltwab, W.A. Elsaghayer and H. Ali, 2015. Ameliorating effect of olive leaf extract on cyclosporineinduced nephrotoxicity in rats. Iran. J. Kidney Dis., 9: 361-368.
Direct Link - Ojo, O.O., A.O. Ajayi and Anibijuwon, 2007. Antibacterial potency of methanol extracts of lower plants. J. Zhejiang Univ. Sci. B, 8: 189-191.
CrossRefDirect Link - Domitrovic, R., H. Jakovac and G. Blagojevic, 2011. Hepatoprotective activity of berberine is mediated by inhibition of TNF-α, COX-2 and iNOS expression in CCl4-intoxicated mice. Toxicology, 280: 33-43.
CrossRefDirect Link - Uchiyama, M. and M. Mihara, 1978. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem., 86: 271-278.
CrossRefDirect Link - Ellman, G.L., 1959. Tissue sulfhydryl groups. Arch. Biochem. Biophys., 82: 70-77.
CrossRefPubMedDirect Link - Sellins, K.S. and J.J. Cohen, 1995. Nuclear changes in the cytotoxic T lymphocyte-induced model of apoptosis. Immunol. Rev., 146: 241-266.
CrossRefDirect Link - Liu, Y., X. Lu, S. Nguyen, J.L. Olson, H.K. Webb and D.L. Kroetz, 2013. Epoxyeicosatrienoic acids prevent cisplatin-induced renal apoptosis through a p38 mitogen-activated protein kinase-regulated mitochondrial pathway. Mol. Pharmacol., 84: 925-934.
CrossRefDirect Link - Sahu, B.D., K.K.R. Rentam, U.K. Putcha, M. Kuncha, G.M.N. Vegi and R. Sistla, 2011. Carnosic acid attenuates renal injury in an experimental model of rat cisplatin-induced nephrotoxicity. Food Chem. Toxicol., 49: 3090-3097.
CrossRefDirect Link - Noori, S. and T. Mahboob, 2010. Antioxidant effect of carnosine pretreatment on cisplatin-induced renal oxidative stress in rats. Indian J. Clin. Biochem., 25: 86-91.
CrossRefDirect Link - Liu, Y., A.M. Sun and L.D. Dworkin, 1998. Hepatocyte growth factor protects renal epithelial cells from apoptotic cell death. Biochem. Biophys. Res. Commun., 246: 821-826.
CrossRefDirect Link - Basnakian, A.G., E.O. Apostolov, X. Yin, M. Napirei, H.G. Mannherz and S.V. Shah, 2005. Cisplatin nephrotoxicity is mediated by deoxyribonuclease I. J. Am. Soc. Nephrol., 16: 697-702.
CrossRefDirect Link - Chtourou, Y., B. Aouey, M. Kebieche and H. Fetoui, 2015. Protective role of naringin against cisplatin induced oxidative stress, inflammatory response and apoptosis in rat striatum via suppressing ROS-mediated NF-κB and P53 signaling pathways. Chem.-Biol. Interactions, 239: 76-86.
CrossRefDirect Link - Santos, N.A.G., C.S. Catao, N.M. Martins, C. Curti, M.L.P. Bianchi and A.C. Santos, 2007. Cisplatin-induced nephrotoxicity is associated with oxidative stress, redox state unbalance, impairment of energetic metabolism and apoptosis in rat kidney mitochondria. Arch. Toxicol., 81: 495-504.
CrossRefPubMedDirect Link - Hanigan, M.H. and P. Devarajan, 2003. Cisplatin nephrotoxicity: Molecular mechanisms. Cancer Ther., 1: 47-61.
Direct Link - Chirino, Y.I., J. Trujillo, D.J. Sanchez-Gonzalez, C.M. Martinez-Martinez, C. Cruz, N.A. Bobadilla and J. Pedraza-Chaverri, 2008. Selective iNOS inhibition reduces renal damage induced by cisplatin. Toxicol. Lett., 176: 48-57.
CrossRefDirect Link - Yuan, J.J., C.Z. Wang, J.Z. Ye, R. Tao and Y.S. Zhang, 2015. Enzymatic hydrolysis of oleuropein from Olea europea (olive) leaf extract and antioxidant activities. Molecules, 20: 2903-2921.
CrossRefDirect Link - De Marino, S., C. Festa, F. Zollo, A. Nini, L. Antenucci, G. Raimo and M. Iorizzi, 2014. Antioxidant activity and chemical components as potential anticancer agents in the olive leaf (Olea europaea L. cv Leccino.) decoction. Anti-Cancer Agents Med. Chem., 14: 1376-1385.
Direct Link - Ramesh, G. and W.B. Reeves, 2004. Inflammatory cytokines in acute renal failure. Kidney Int., 66: S56-S61.
CrossRefPubMedDirect Link - Li, S., N. Gokden, M.D. Okusa, R. Bhatt and D. Portilla, 2005. Anti-inflammatory effect of fibrate protects from cisplatin-induced ARF. Am. J. Physiol. Renal Physiol., 289: F469-F480.
CrossRefDirect Link - Kelliher, M.A., S. Grimm, Y. Ishida, F. Kuo, B.Z. Stanger and P. Leder, 1998. The death domain kinase RIP mediates the TNF-induced NF-κB signal. Immunity, 8: 297-303.
CrossRefDirect Link - Gong, D., C. Geng, L. Jiang, L. Wang, H. Yoshimura and L. Zhong, 2012. Mechanisms of olive leaf extract‐ameliorated rat arthritis caused by kaolin and carrageenan. Phytother. Res., 26: 397-402.
CrossRefDirect Link - Prasad, K.N., 2004. Multiple dietary antioxidants enhance the efficacy of standard and experimental cancer therapies and decrease their toxicity. Integr. Cancer Therapies, 3: 310-322.
CrossRefDirect Link - Kaushal, G.P., V. Kaushal, X. Hong and S.V. Shah, 2001. Role and regulation of activation of caspases in cisplatin-induced injury to renal tubular epithelial cells. Kidney Int., 60: 1726-1736.
CrossRefPubMedDirect Link - Offer, H., N. Erez, I. Zurer, X. Tang, M. Milyavsky, N. Goldfinger and V. Rotter, 2002. The onset of p53-dependent DNA repair or apoptosis is determined by the level of accumulated damaged DNA. Carcinogenesis, 23: 1025-1032.
CrossRefDirect Link - Cummings, B.S. and R.G. Schnellmann, 2002. Cisplatin-induced renal cell apoptosis: Caspase 3-Dependent and-independent pathways. J. Pharmacol. Exp. Ther., 302: 8-17.
PubMedDirect Link - Jiang, M., Q. Wei, J. Wang, Q. Du, J. Yu, L. Zhang and Z. Dong, 2006. Regulation of PUMA-α by p53 in cisplatin-induced renal cell apoptosis. Oncogene, 25: 4056-4066.
CrossRefDirect Link - Jia, G., C. Tohyama and H. Sone, 2002. DNA damage triggers imbalance of proliferation and apoptosis during development of preneoplastic foci in the liver of Long-Evans Cinnamon rats. Int. J. Oncol., 21: 755-761.
Direct Link - Bonventre, J.V., 2003. Dedifferentiation and proliferation of surviving epithelial cells in acute renal failure. J. Am. Soc. Nephrol., 14: S55-S61.
PubMedDirect Link - Wang, Y., S. Wang, W. Cui, J. He, Z. Wang and X. Yang, 2013. Olive leaf extract inhibits lead poisoning-induced brain injury. Neural Regeneration Res., 8: 2021-2029.
CrossRefDirect Link - Bali, E.B., V. Ergin, L. Rackova, O. Bayraktar, N. Kucukboyaci and C. Karasu, 2014. Olive leaf extracts protect cardiomyocytes against 4-hydroxynonenal-induced toxicity in vitro: Comparison with oleuropein, hydroxytyrosol and quercetin. Planta Med., 80: 984-992.
CrossRefPubMedDirect Link - Kusirisin, W., C. Jaikang, C. Chaiyasut and P. Narongchai, 2009. Effect of polyphenolic compounds from Solanum torvum on plasma lipid peroxidation, superoxide anion and cytochrome P450 2E1 in human liver microsomes. Med. Chem., 5: 583-588.
CrossRefDirect Link - Wang, K.J., Y.J. Zhang and C.R. Yang, 2006. New phenolic constituents from Balanophora polyandra with radical‐scavenging activity. Chem. Biodiversity, 3: 1317-1324.
CrossRefDirect Link