
Elif Kaya-Tilki
Department of Pharmacology, Faculty of Pharmacy, Anadolu University, Tepebasi 26470, Eskişehir, Turkey
Miris Dikmen
Department of Pharmacology, Faculty of Pharmacy, Anadolu University, Tepebasi 26470, Eskişehir, Turkey
Yusuf Ozturk
Department of Pharmacology, Faculty of Pharmacy, Anadolu University, Tepebasi 26470, Eskişehir, Turkey
LiveDNA: 90.2044
International Journal of Pharmacology
Year: 2016 | Volume: 12 | Issue: 4 | Page No.: 351-360







ABSTRACT
Neuropathological characteristics of neurodegenerative disorders are atrophy or loss of specific neurons in the specific brain areas. Small-molecule products like epigenetic drugs, which can activate silenced genes, regulate chromatin remodelling and transcription, is presently been focused as an alternative approach for eliciting neuritogenic activity. Trichostatin A (TSA) has been effectively investigated for cancer treatment but it also has been shown to possess neuritogenic potential. In this study, it was aimed to compare the neuritogenic effects of two epigenetic drugs on PC-12 Adh cell line, which are the histone deacetylase inhibitor (HDACi) trichostatin A and a novel non-covalent DNA methyltransferase inhibitor (DNMTi) RG108. Cytotoxic effects were determined by MTT assay. Cell differentiation and migration analysis were evaluated by xCELLigence Real Time Cell Analyzer Dual Plate (RTCA DP) system according to the changing in Cell Index (CI) values. Cell migration analysis was also corroborated with morphological migration analysis. Matrix metalloproteinase-2 (MMP-2) levels related with neurite outgrowth were measured by ELISA. Finally, neurite outgrowth was observed with immunofluorescence staining and determined by neurite outgrowth analysis on PC-12 Adh cell line. The RG108 was found nearly as effective as TSA on neuronal differentiation, neurite outgrowth and cellular migration in PC-12 Adh cell line in a combination with nerve growth factor. According to the results, DNA methyltransferase inhibitors also hold promise in neurodegenerative disorders.
PDF Abstract XML References Citation
How to cite this article
Elif Kaya-Tilki, Miris Dikmen and Yusuf Ozturk, 2016. Effects of DNMT and HDAC Inhibitors (RG108 and Trichostatin A) on NGF-induced Neurite Outgrowth and Cellular Migration. International Journal of Pharmacology, 12: 351-360.
DOI: 10.3923/ijp.2016.351.360
URL: https://scialert.net/abstract/?doi=ijp.2016.351.360
DOI: 10.3923/ijp.2016.351.360
URL: https://scialert.net/abstract/?doi=ijp.2016.351.360
INTRODUCTION
The Central Nervous System (CNS) is characterized by a weak spontaneous regenerative capacity and has limited capacity of neurons to regrow injured axons (Oberbauera et al., 2013). The insufficiency of regeneration is based on a combination of different factors including reduced capacity of regeneration, lack of necessary trophic molecules to support growth and the presence of an environment hostile for any growth line (Woolf, 2003).
Neuroregeneration in the adult CNS is a multistep process that contains new neurons, generated through proliferation of endogenous stem/progenitor cells that will differentiate or the survival of injured neurons and both of them integrate into existing neural networks. Neurite outgrowth is the process in which neurons elongate their neurites to reach their targets (pathfinding), leading to the establishment of neuronal connections during development (Meur et al., 2009). The initiation and guidance of a neurite originate by several signal inputs from the extracellular space. The extracellular matrix (ECM) is a complex structure that contains chondroitin sulfate proteoglycans (CSPGs), which are produced by glial cells both during CNS development and injury (Kiryushko et al., 2004). The MMP-2 is one of the enzymes which is involved in the regulation of cell motility and neurite outgrowth via the degradation of inhibitory CSPGs for rendering the nerve more permissive to axon regeneration (Ferguson and Muir, 2000). Nerve Growth Factor (NGF) is also a extracellular matrix-associated neurotrophic factor (neurite promoter) that is essential for survival and differentiation of neuronal cells (Kiryushko et al., 2004; Huang and Reichardt, 2001; Segal, 2003). The NGF signaling is initiated when it binds to TrkA receptor at the cell surface, leading to transient activation of the Ras/MAPK pathway and phosphoinositide 3-kinase that promotes cell survival (Zhang et al., 2000). However, the delivery of exogenous neurotrophic factors is not effective enough for neurodegenerative diseases treatment. Neurotrophic factors are large polypeptide molecules that do not penetrate the blood brain barrier. In addition, they are metabolized by peptidases when administered peripherally (Maruoka et al., 2011). Therefore, potentiating the actions of NGF with small molecular weight compounds can be an alternative therapeutic approach.
Cellular and molecular changes can interact with genes and environmental factors to determine which cells age successfully and which go towards neurodegeneration (Hindle, 2010). Epigenetics refers to the reversible regulation of various genomic function that occur without a change in the DNA sequence and mediated through changes in DNA methylation and chromatin structure which can be an alternative approach in the neurodegenerative diseases treatment (Chouliaras et al., 2010).
DNA methylation and histone acetylation are interdependent and maintain equilibrium, allowing temporal expression of genes in normal cells, whereas in neurodegenerative disorders this balance might be disrupted. As HDACi, DNMTi are also low molecular weight compounds which are currently being developed for cancer therapy. Besides of anticarcinogenic effects, neuritogenic effects of HDACi were shown earlier, however the effects of DNMTi on neurite outgrowth and neuronal migration of PC-12 Adh cells, which respond to NGF with a dramatic change in phenotype and acquire a number of properties characteristic of sympathetic neurons (Greene and Tischler, 1976) are still not clear. In order to investigate the neuritogenic potential of DNMTi, the effects of RG108, a novel non-covalent DNMT1 inhibitor with lower cytotoxic properties according to the other agents in the group, was compared with a HDACi trichostatin A (TSA) on PC-12 Adh neurite outgrowth and cell migration.
MATERIALS AND METHODS
Cell culture: The PC-12 Adh (CRL-1721.1™) cell line was obtained from American Type Culture Collection (ATCC, USA) and maintained undifferentiated in DMEM medium containing 10% horse serum, 5% fetal bovine serum and 1% penicillin/streptomycin at 37°C in a humidified incubator with 5% CO2.
To induce differentiation, 1% fetal bovine serum and 1% penicillin/streptomycin containing DMEM differentiation medium was used. Neuronal Growth Factor (NGF) was used as a co-stimulator in 100 nM concentration (Bang et al., 2001; Parker et al., 2000). TSA and RG108 was dissolved in DMSO and diluted to working concentrations with fresh medium. The highest concentration of DMSO was used in 200 μM which was less than 0.01%. Control group was prepared with medium containing 0.1% DMSO. The growth curve of each cell line was assessed.
Cell viability assay: In order to obtain non-cytotoxic concentrations of the compounds, neuronal viability was determined by MTT assay as described previously (Hansen et al., 1989). In short, cells were grown in 96-well plates at a density of 3×103 cells per well and subjected to different RG108 and TSA (10 and 100 nM; 1, 10, 100, 200 μM) concentrations. After 24 or 48 h incubation, MTT solution was added to reach a final concentration of 0.5 mg mL–1. The cells] were incubated for another 4 h. Then, current medium was removed and 100 μL of DMSO solution was added. The absorbance was measured at 540 nm using a Cytation 3 Cell Imaging Multi-Mode Reader (Bio-Tek). Cell survival rates were expressed as the percentage of the DMSO (0.1%) solvent control.
Cell differentiation assay with Real-Time Cell Analysis system (RTCA DP): To investigate the differentiation of PC-12 Adh cells and determine the changing over time, a real-time cell analysis was performed with xCELLigence DP instrument by using E-plates that contain micro-electrodes and measuring electrical impedance which indicated as Cell Index (CI) value (a dimensionless parameter to represent cell status). This method is a modification of cell proliferation assay based on a characteristic feature of PC-12 cells, they stop proliferating with NGF signaling (Mustafa et al., 2015, 2016; Dwane et al., 2013). Concentrations were used as 10 and 100 nM, which showed lower cytotoxic effects according to the results of the MTT assay, alone or in combination with 100 nM NGF. Background of the E-plates were measured in 100 μL differentiation medium in the Real-Time Cell Analyzer (RTCA DP) station. Afterwards, cells were seeded in a 2×104 density per well on to 96-well E-plates (Roche Applied Sciences, Indianapolis, IN) in 100 μM differentiation medium. Cells were incubated up to 24 h and the impedance of each well was monitored using the RTCA device (xCELLigence, ACEA Biosciences, San Diego, CA) at 1 h intervals. After incubation the instrument was paused, 100 μL of the current medium was removed, twice of the concentrations were added into the wells and diluted to the final concentrations in differentiation medium. Assays were performed in octet and cell-free and concentration-free (with 0.1% DMSO) controls were run in parallel. Cells were monitored for up to 7 days at 1 h intervals in order to determine the cell differentiation comparatively, according to the CI values of the instrument. Neuronal differentiation percentage graph was drawn according to these CI values with GraphPad Prism V6 (http://www.graphpad.com/scientific-software/prism/).
Cell migration assay with Real-Time Cell Analysis system (RTCA DP): The RTCA DP instrument uses the CIM-Plate 16 for migration analysis, which contains microelectronic sensors connected on to the underside of the plate (Limame et al., 2012). For migration assay, the wells of the bottom chamber were filled with 160 μL of 10% serum containing DMEM medium and the top and bottom parts of the CIM-16 plates were assembled together. After the addition of 20 μL serum-free DMEM medium to the top chamber wells, the assembled CIM-plate 16 was allowed to stabilize for 1 h at 37°C, 5% CO2. After the incubation, cells were seeded at a 2×104 density per well on to the top chambers of CIM-16 plates in 80 μL of serum-free media, then 100 μL of 100 nM NGF alone or combined with 100 nM RG108 and TSA concentrations were added. CIM-plate 16 was placed into the RTCA DP for data collection after 30 min incubation at room temperature. The RTCA DP software was set to monitor CI values once every 10 min for 72 h.
Morphological analysis of cell migration: Morphological migration analysis was performed with The Oris™ Cell Migration Assay kit (Platypus Technologies, LLC., Madison, WI) following the manufacturer’s instructions. Briefly, cells were collected in differentiation medium at 5×104 cells mL–1 and 100 μL of the cell suspension was added to each well of the Oris plate. After 6 h incubation at 37°C in a 5% CO2 incubator, the stoppers and then current medium were removed from the Oris plate. About 100 μL of 100 nM NGF alone or combined with 100 nM RG108 and TSA concentrations were added to wells. Attached cells were incubated for 72 h and every 24 h 3 wells from all concentrations were stained with 10 μL Hoechst 33258 for 5 min and photographed with an Oris detection mask in Cytation 3 Cell Imaging Multi-Mode Reader (Bio-Tek) using the blue filter cube.
Neurite outgrowth analysis: PC-12 Adh cells were plated on to the collagen IV coated 96-well culture plate (BD BioCoat™ Collagen IV Cellware) at a density of 2×103 cells per well in differentiation medium. Six hours after plating, medium was replaced with 100 μL of 100 nM NGF alone or combined with 10 and 100 nM RG108 and TSA concentrations. Seven days incubation period was determined according to the results of the differentiation analysis in RTCA DP system. On 7th day, morphometric analysis was performed on digitized images of live cells taken under a Leica DM 300 inverted microscope. Neurite outgrowth analysis was performed as described by Tamplenizza et al. (2013) with some modifications. Briefly, neurite growth was determined by manually tracing the length of the longest neurite and branch (if present) on images by using Leica LAS Image Analysis programme in pixels then converted to μm by using ImageJ (NIH). Totally 100 neurites were measured for each concentration group. Total length was divided to 100 to find the average neurite length per group. Experiments were repeated at least three times independently.
Quantifying MMP-2 levels: For quantifying the MMP-2 levels related with neurite outgrowth, an ELISA assay was performed. Cells were seeded on a collagen IV coated 6-well plate at a 4×104 density per well in differentiation medium. After 6 h, medium was replaced with 100 nM NGF alone or combined with 10 and 100 nM RG108 and TSA concentrations containing 2 mL differentiation medium. On 7th day, supernatants of the cells were collected from wells for each group and stored at -80°C until the day of the assay was performed. A MMP 2 Rat ELISA Kit (USCN Life Science, Cat# SEA100Ra) was used and the analysis was performed according to the kit procedure.
Immunoflorescence staining: After neurite outgrowth analysis, cells were fixated with 80% methanol for 5 min. Cells were rinsed twice with 1×PBS then permeabilized with 0.1% PBS-Tween 20 for 20 min. After two washes in PBS, the cells were incubated in 1% BSA/5% fetal bovine 5% horse serum/0.3M glycine in 0.1% PBS-Tween solution for 1 h to permeabilise the cells and block non-specific protein-protein interactions. The anti-beta III tubulin antibody [2G10] (Abcam, Cat# ab78078, RRID: AB-2256751) was diluted 1:250 in the same solution and then the cells were incubated with 100 μL of antibody overnight at +4°C. After the incubation period, cells were rinsed twice with PBS and secondary antibody goat anti-mouse IgG H and L (Alexa Fluor®488), (Abcam, Cat# ab150113) at a 1:500 dilution was added to the wells and incubated for 1 h in room temperature. Hoechst 33258 (10 μg mL–1) was used to stain the cell nuclei (blue) 10 min before the imaging with Cytation 3 Cell Imaging Multi-Mode Reader (Bio-Tek, USA).
Statistical analysis: Data were analyzed by one way ANOVA with Tukey’s post-hoc, expressed as Mean±Standard error, p>0.05 not significant, p<0.05*, p<0.01** and p<0.001***.
RESULTS
Effects of trichostatin A and RG108 on cell viability: According to the MTT assay results, the cell viability significantly decreased with 200, 100 and 10 μM TSA concentrations for 24 and 48 h. On the other hand, 200 and 100 μM RG108 concentrations slightly decreased the cell viability (p>0.05 not significant) (Fig. 1). Non-cytotoxic concentrations were determined as 10 and 100 nM for further experiments.
Effects of trichostatin A and RG108 on cell differentiation: The CI values of concentrations without NGF and the control group obtained from RTCA DP instrument were like a plateau; however, the concentration groups containing NGF have showed a rapid increase after 4th day (Fig. 2a). Neuronal differentiation percentage graph was drawn according to the CI values on 4th day (Fig. 2b). According to the graphics, around 4th day, differentiation has been started and the experiment was terminated around 7th day because of the decrease in the CI values, that may be arising from the insufficient medium in the wells. The rapid increase in the CI values were accepted as the initial stages of differentiation (Dwane et al., 2013). Because during the initiation, cell body becomes larger which cause an increase in the impedance and then, the cell begins to extend neurites and cell body shrinks which cause a drop in the impedance.
Effects of trichostatin A and RG108 on cell migration: The most effective concentration on PC-12 Adh cell differentiation, 100 nM TSA and RG108 in a combination with 100 nM NGF, were choosen for migration analysis according to the results of the previous cell differentiation assay.
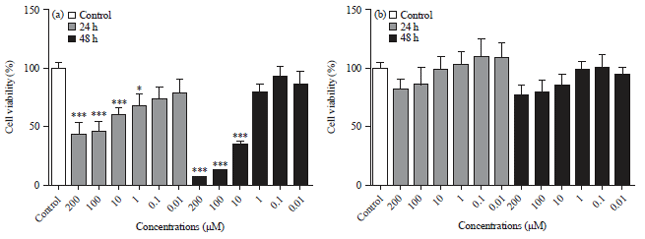 | |
| Fig. 1(a-b): | Effects of (a) TSA and (b) RG108 on PC-12 Adh cell viability. The results are the means of 3 independent experiments. The error bars represent the standard deviations ((n = 8), p>0.05 not significant, *p<0.05 and ***p<0.001, compared to control) |
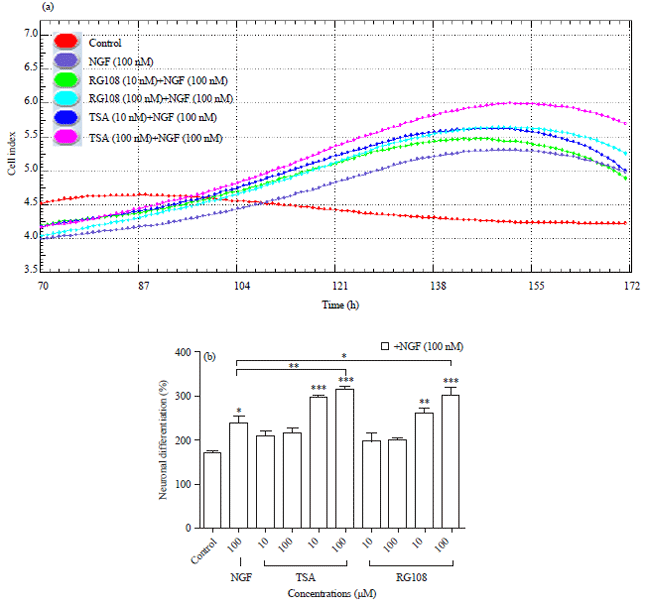 | |
| Fig. 2(a-b): | Real-time monitoring of differentiative effects of (a) TSA and RG108 on PC-12 Adh cell line using RTCA-DP System (n = 8) and (b) Neuronal differentiation percentage graph was drawn according to these CI values p>0.05 not significant, *p<0.05, **p<0.01 and ***p<0.001, compared to control |
The level of cell migration was assessed by increases in the slope of the curve during active migration. Within the first 24 h, both TSA and RG108 combinations showed migration kinetics similar to that of NGF. In keeping with the results of the endpoint measurement, after 72 h, RG108 and TSA combination with NGF were caused higher migration compared with the positive control NGF (Fig. 3a). In addition to the results of migration analysis conducted with RTCA DP, according to the morphological migration analysis images shown in Fig. 3b, the effects of TSA combination was slightly higher than RG108 combination.
Effects of TSA and RG108 on NGF-induced neurite outgrowth: According to the results, 100 nM TSA in a combination with 100 nM NGF showed the highest neuritogenic effect (1430.7 μm) on PC-12 Adh cells. Average neurite length of the cells treated with 100 nM RG108 in a combination with 100 nM NGF was also showed slightly higher neuritogenic effect (1185.6 μm) than the cells treated with 10 nM combination groups. This data supports our neuronal differentiation and migration analysis results and taking together all this data may point a dose-dependent increase in neuritogenic effects (Fig. 4 and 5a).
Effects of TSA and RG108 on MMP-2 levels: After injury or degeneration, MMP-2 is involved in the regulation of neurite outgrowth via the processing of inhibitory CSPGs that are upregulated in the CNS and modulates cell migration through the release or inactivation of chemotactic signals (Rodriguez et al., 2010; Zhang et al., 2007).
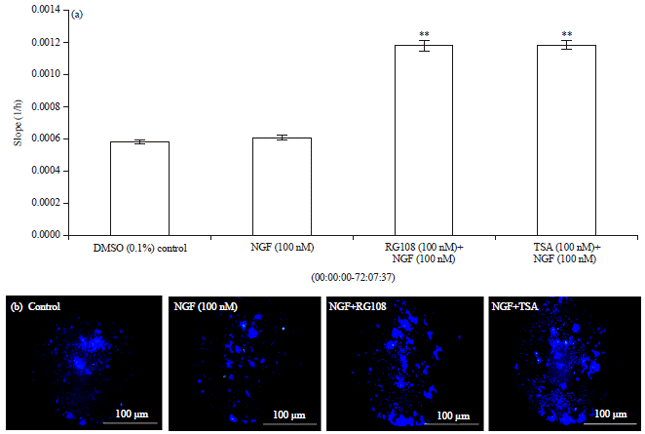 | |
| Fig. 3(a-b): | Migration assay results were obtained with the xCELLigence RTCA DP system. (a) The slope of the migration curve was calculated using the RTCA 1.2.1 Software and (b) Morphological migration analysis images after 72 h, cells were stained with Hoechst 33258 and images were taken by Cytation 3 Cell Imaging Multi-Mode Reader with an Oris detection mask (n = 4), p>0.05 not significant and **p<0.01 and compared to control) |
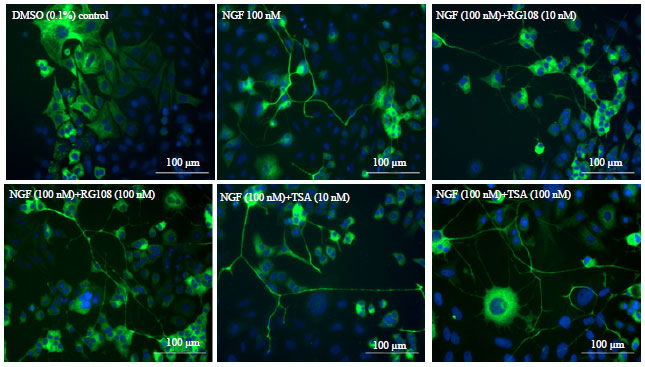 | |
| Fig. 4: | Images of PC-12 Adh cells treated with various NGF, TSA and RG108 concentrations on 7th day. Cell nuclei were stained with Hoechst 33258 (blue) and tubulins were stained with AlexaFluor®488 (green). Images were taken with Cytation 3 Cell Imaging Multi-Mode Reader with 20X objective |
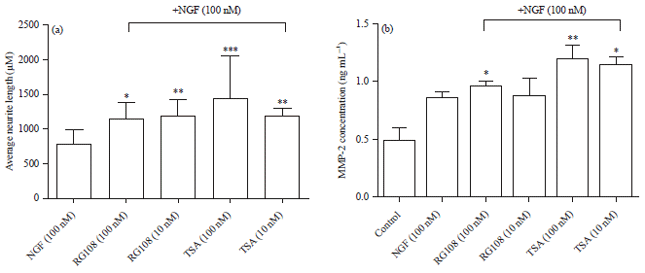 | |
| Fig. 5(a-b): | Comparison of (a) Average neurite lengths and (b) Changes in MMP-2 levels. Data are Mean±SD values, p>0.05 not significant, *p<0.05, **p<0.01 and ***p<0.001 |
MMP-2 levels of control, NGF (100 nM), NGF (100 nM) combination with TSA (100 and 10 nM) and RG108 (100 and 10 nM) were 0.48, 0.86, 0.96, 0.87, 1.20 and 1.14 ng mL–1, respectively (Fig. 5b). According to the results, the MMP-2 levels of RG108 (100 nM) and TSA (10 and 100 nM)+NGF (100 nM) combinations were significantly increased compared with the control and NGF (100 nM) groups.
DISCUSSION
In this study, the epigenetic molecular mechanisms’ relevance to neuronal differentiation were tried to understand. Histone deacetylase inhibitors (Sun et al., 2014; Cheng et al., 2012; Hamner et al., 2008; Choi et al., 2001) and DNMT inhibitors (Gnyszka et al., 2013; Savickiene et al., 2012a, b) are currently under development for cancer treatment. In terms of CNS applications, similar to the cancer therapy, where a disease treatment requires an extensive reorganization in epigenome, the drugs that can broadly affect the epigenome are desirable. Therefore, epigenetic drugs hold a great promise for the therapeutic efficacy in neurodegenerative disorders (Day et al., 2015). For this purpose, comparison of the effects of DNMTi RG108 and a HDACi TSA on PC-12 Adh neuronal differentiation and neurite outgrowth was done which is a very popular model for studying cell differentiation and have capacity to grow neurite-like processes in response to NGF (Greene and Tischler, 1976). Significant induction of neurite outgrowth and cellular migration with both RG108 and TSA synergistically with NGF were defined. This result was also supported with immunofluorescence imaging, morphologic and quantitative cell migration assay and measurement of MMP-2 levels related with neurite outgrowth which is an important process during neurite outgrowth. According to the results, RG108 is nearly as effective as TSA at 10 and 100 nM concentrations with lower cytotoxicity.
Neuronal injury and degeneration are responsible for various neurodegenerative disorders that are associated with loss of brain cells and axons resulting in functional deficits. Due to limited regeneration capacity of damaged neurons, the regulation of neurite outgrowth is crucial in developing strategies to promote axon and dendrite regeneration after nerve injury and in degenerative diseases (Wu et al., 2012). Cell migration establishes the formation of a highly specific pattern of connections between nerve cells and it is required for axonal guidance and neurite outgrowth and involves a number of coordinated signalling pathways (Dwane et al., 2013). Previous studies have indicated that neurite outgrowth and neuronal cell migration are important steps of neuroregeneration (Holtmaat et al., 1998; Struzyna et al., 2015).
Many different environmental factors can modulate epigenetic markers during development and in adult organism. In turn, epigenetics can cause and is associated with several neurodegenerative diseases. One potential method for preventing cell death and stimulating regeneration is to alter the intracellular signals which a cell uses to transduce responses to injury or growth-inhibiting substrates. For example, in apoptotic cell death, several transcription factors, including fas, p53, c-Jun, bax and bcl-2 are involved and may represent common pathways to intervene and prevent cell loss (Bredesen, 1995). Epigenetic modifications modulate gene expression, mediated by altering the transcription factor activities, during several physiological processes, mainly during embryonic development and cellular differentiation (Nicolia et al., 2015).
Increased histone acetylation and transcription factor activity have been shown to protect from apoptosis in animal models of neurodegeneration (Saha and Pahan, 2006) and to promote neuronal differentiation (Balasubramaniyan et al., 2006; Schwechter et al., 2007; Maruoka et al., 2011). The TSA, directly inhibits histone deacetylases and allows histone acetyltransferases to hyperacetylate H3 at K9 and p53 at their C-terminus. The p53 forms transcriptional complexes with CBP/p300 and P/CAF. Then, in concert with CBP/p300 and P/CAF transcription co-factors, TSA causes to occupy specific promoters within the newly formed permissive chromatin state (Kiryushko et al., 2004; Tedeschi et al., 2009; Di Giovanni et al., 2006; Gaub et al., 2010). This situation results with the increased expression of neuronal outgrowth related genes like coronin 1b and GAP-43 (Kiryushko et al., 2004). Growth-associated proteins such as tubulin, actin and GAP-43 are thought to act as precursors of critical growth periods after injury and during development (Bisby and Tetzlaff, 1992; Hopker et al., 1999).
DNA methylation is also related with gene transcription which is mediated by DNMTs. In theory, a single cytosine methylation can completely silence the associated gene (Day et al., 2015). The RG108 is a novel DNMT1 enzyme inhibitor with low cytotoxicity and genotoxicity (Stresemann et al., 2006), this is the first paper that reports the neuritogenic effects of RG108 synergistically with NGF. The role of DNMT inhibitors on neurite outgrowth is still not clear but a study about the role of epigenetic reprogramming in cell differentiation which was investigated the NGF-induced differentiation of PC-12 cells was reported that the mRNA and protein levels of DNMT1 and DNMT3a decreased whereas, de novo methyltransferase DNMT3b increased during neurite outgrowth (Bai et al., 2005). Another study showed that upregulation of DNMT involved in motor neuron apoptosis and DNMT inhibitors, like RG108, protected motor neurons from apoptosis in vivo and in vitro (Chestnut et al., 2011). In addition, an interesting study was performed by Miller and Sweatt (2007) revealed that following contextual fear conditioning, DNMT gene expression is upregulated in the adult rat hippocampus and that DNMT inhibition blocks memory formation and also it activates the synaptic plasticity transcriptional gene reelin (Miller and Sweatt, 2007). Reelin is a highly expressed ECM glycoprotein gene during neuronal migration and neuroplasticity and responsible from cell to cell interactions (D’Arcangelo, 2005; Weeber et al., 2002). It regulates cell migration and positioning in the developing brain and also its function depends upon its proteolytic cleavage by MMPs and inhibition of MMPs causes a significant decrease in migration of neuron cells (Maurya et al., 2014; Tinnes et al., 2011; Ayoub et al., 2005). Therefore, according to the concentration groups in which higher MMP-2 levels were obtained, were also showed higher neuritogenic effects, this data might point a relation between reelin pathway.
CONCLUSION
In conclusion, promotion of an environment conducive to regrowth of the degenerated neurons by potentiating the effects of NGF with low molecular weight compounds, like RG108 and TSA, that can potantiate the action of neurotrophic factors might hold promise for counteracting neurodegeneration and with further investigations can be regarded as potential drug candidates. As another epigenetic modifier drug group, DNMTi also deserves interest in terms of these effects. Taking together all this data points that DNA methylation is a dynamic process which needs further investigations to understand in this field.
ACKNOWLEDGMENT
This study was performed at Anadolu University, Faculty of Pharmacy, Pharmacology Cell Culture Laboratory in Eskisehir, Turkey.
REFERENCES
- Ayoub, A.E., T.Q. Cai, R.A. Kaplan and J. Luo, 2005. Developmental expression of matrix metalloproteinases 2 and 9 and their potential role in the histogenesis of the cerebellar cortex. J. Comp. Neurol., 481: 403-415.
CrossRefDirect Link - Balasubramaniyan, V., E. Boddeke, R. Bakels, B. Kust, S. Kooistra, A. Veneman and S. Copray, 2006. Effects of histone deacetylation inhibition on neuronal differentiation of embryonic mouse neural stem cells. Neuroscience, 143: 939-951.
CrossRefDirect Link - Bang, O.S., E.K. Park, S.I. Yang, S.R. Lee, T.F. Franke and S.S. Kang, 2001. Overexpression of Akt inhibits NGF-induced growth arrest and neuronal differentiation of PC12 cells. J. Cell Sci., 114: 81-88.
Direct Link - Bisby, M.A. and W. Tetzlaff, 1992. Changes in cytoskeletal protein synthesis following axon injury and during axon regeneration. Mol. Neurobiol., 6: 107-123.
PubMedDirect Link - Chestnut, B.A., Q. Chang, A. Price, C. Lesuisse, M. Wong and L.J. Martin, 2011. Epigenetic regulation of motor neuron cell death through DNA methylation. J. Neurosci., 31: 16619-16636.
CrossRefPubMedDirect Link - Choi, J.H., H.J. Kwon, B.I. Yoon, J.H. Kim, S.U. Han, H.J. Joo and D.Y. Kim, 2001. Expression profile of histone deacetylase 1 in gastric cancer tissues. Jpn. J. Cancer Res., 92: 1300-1304.
CrossRefDirect Link - Chouliaras, L., B.P.F. Rutten, G. Kenis, O. Peerbooms and P.J. Visser et al., 2010. Epigenetic regulation in the pathophysiology of Alzheimer's disease. Prog. Neurobiol., 90: 498-510.
CrossRefDirect Link - D'Arcangelo, G., 2005. Apoer2: A reelin receptor to remember. Neuron, 47: 471-473.
CrossRefDirect Link - Day, J.J., A.J. Kennedy and J.D. Sweatt, 2015. DNA methylation and its implications and accessibility for neuropsychiatric therapeutics. Annu. Rev. Pharmacol. Toxicol., 55: 591-611.
CrossRefDirect Link - Di Giovanni, S., C.D. Knights, M. Rao, A. Yakovlev and J. Beers et al., 2006. The tumor suppressor protein p53 is required for neurite outgrowth and axon regeneration. EMBO J., 25: 4084-4096.
CrossRefDirect Link - Cheng, D.D., Q.C. Yang, Z.C. Zhang, C.X. Yang and Y.W. Liu, 2012. Antitumor activity of histone deacetylase inhibitor trichostatin A in osteosarcoma cells. Asian Pac. J. Cancer Prev., 13: 1395-1399.
CrossRefDirect Link - Dwane, S., E. Durack and P.A. Kiely, 2013. Optimising parameters for the differentiation of SH-SY5Y cells to study cell adhesion and cell migration. BMC Res. Notes, Vol. 6.
CrossRefDirect Link - Ferguson, T.A. and D. Muir, 2000. MMP-2 and MMP-9 increase the neurite-promoting potential of schwann cell basal laminae and are upregulated in degenerated nerve. Mol. Cell. Neurosci., 16: 157-167.
CrossRefDirect Link - Gaub, P., A. Tedeschi, R. Puttagunta, T. Nguyen, A. Schmandke and S. di Giovanni, 2010. HDAC inhibition promotes neuronal outgrowth and counteracts growth cone collapse through CBP/p300 and P/CAF-dependent p53 acetylation. Cell Death Differ., 17: 1392-1408.
CrossRefDirect Link - Gnyszka, A., Z. Jastrzebski and S. Flis, 2013. DNA methyltransferase inhibitors and their emerging role in epigenetic therapy of cancer. Anticancer Res., 33: 2989-2996.
Direct Link - Greene, L.A. and A.S. Tischler, 1976. Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc. Natl. Acad. Sci. USA., 73: 2424-2428.
Direct Link - Hamner, J.B., T.L. Sims, A. Cutshaw, P.V. Dickson and S. Rosati et al., 2008. The efficacy of combination therapy using adeno-associated virus-interferon β and trichostatin a in vitro and in a murine model of neuroblastoma. J. Pediatr. Surg., 43: 177-183.
CrossRefDirect Link - Hansen, M.B., S.E. Nielsen and K. Berg, 1989. Re-examination and further development of a precise and rapid dye method for measuring cell growth/cell kill. J. Immunol. Methods, 119: 203-210.
CrossRefPubMedDirect Link - Hindle, J.V., 2010. Ageing, neurodegeneration and Parkinson's disease. Age Ageing, 39: 156-161.
CrossRefDirect Link - Holtmaat, A.J.G.D., A.B. Oestreicher, W.H. Gispen and J. Verhaagen, 1998. Manipulation of gene expression in the mammalian nervous system: Application in the study of neurite outgrowth and neuroregeneration-related proteins. Brain Res. Rev., 26: 43-71.
CrossRefDirect Link - Hopker, V.H., D. Shewan, M. Tessier-Lavigne, M. Poo and C. Holt, 1999. Growth-cone attraction to netrin-1 is converted to repulsion by laminin-1. Nature, 401: 69-73.
CrossRefDirect Link - Huang, E.J. and L.F. Reichardt, 2001. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci., 24: 677-736.
CrossRefPubMedDirect Link - Kiryushko, D., V. Berezin and E. Bock, 2004. Regulators of neurite outgrowth: Role of cell adhesion molecules. Ann. N. Y. Acad. Sci., 1014: 140-154.
CrossRefDirect Link - Limame, R., A. Wouters, B. Pauwels, E. Fransen and M. Peeters et al., 2012. Comparative analysis of dynamic cell viability, migration and invasion assessments by novel real-time technology and classic endpoint assays. PLoS One, Vol. 7.
CrossRefDirect Link - Maruoka, H., H. Sasaya, K. Sugihara, K. Shimoke and T. Ikeuchi, 2011. Low-molecular-weight compounds having neurotrophic activity in cultured PC12 cells and neurons. J. Biochem., 150: 473-475.
CrossRefDirect Link - Maurya, S.K., J. Mishra, V.K. Tripathi, R. Sharma and M.H. Siddiqui, 2014. Cypermethrin induces astrocyte damage: Role of aberrant Ca2+, ROS, JNK, P38, matrix metalloproteinase 2 and migration related reelin protein. Pestic. Biochem. Physiol., 111: 51-59.
CrossRefDirect Link - Miller, C.A. and J.D. Sweatt, 2007. Covalent modification of DNA regulates memory formation. Neuron, 53: 857-869.
CrossRefDirect Link - Meur, A.M.L., A.N. Mohebiany and T.P. O'Connor, 2009. Varicones and growth cones: Two neurite terminals in PC12 cells. PLoS One, Vol. 4.
CrossRefDirect Link - Mustafa, A.M., G. Caprioli, M. Dikmen, E. Kaya and F. Maggi et al., 2015. Evaluation of neuritogenic activity of cultivated, wild and commercial roots of Gentiana lutea L. J. Funct. Foods, 19: 164-173.
CrossRefDirect Link - Mustafa, A.M., F. Maggi, F. Papa, E. Kaya, M. Dikmen and Y. Ozturk, 2016. Isofuranodiene: A neuritogenic compound isolated from wild celery (Smyrnium olusatrum L., Apiaceae). Food Chem., 192: 782-787.
CrossRefDirect Link - Nicolia, V., M. Lucarellib and A. Fuso, 2015. Environment, epigenetics and neurodegeneration: Focus on nutrition in Alzheimer's disease. Exp. Gerontol., 68: 8-12.
CrossRefDirect Link - Oberbauera, E., C. Urmannc, C. Steffenhagena, L. Bielera and D. Brunnerc et al., 2013. Chroman-like cyclic prenylflavonoids promote neuronal differentiation and neurite outgrowth and are neuroprotective. J. Nutr. Biochem., 24: 1953-1962.
CrossRefDirect Link - Parker, E.M., A. Monopoli, E. Ongini, G. Lozza and C.M. Babij, 2000. Rapamycin, but not FK506 and GPI-1046, increases neurite outgrowth in PC12 cells by inhibiting cell cycle progression. Neuropharmacology, 39: 1913-1919.
CrossRefDirect Link - Rodriguez, D., C.J. Morrison and C.M. Overall, 2010. Matrix metalloproteinases: What do they not do? New substrates and biological roles identified by murine models and proteomics. Biochimica Biophysica Acta (BBA)-Mol. Cell Res., 1803: 39-54.
CrossRefDirect Link - Saha, R.N. and K. Pahan, 2006. HATs and HDACs in neurodegeneration: A tale of disconcerted acetylation homeostasis. Cell Death Different., 13: 539-550.
CrossRefPubMedDirect Link - Savickiene, J., G. Treigyte, V.V. Borutinskaite and R. Navakauskiene, 2012. Antileukemic activity of combined epigenetic agents, DNMT inhibitors zebularine and RG108 with HDAC inhibitors, against promyelocytic leukemia HL-60 cells. Cell. Mol. Biol. Lett., 17: 501-525.
CrossRefDirect Link - Savickiene, J., G. Treigyte, A. Jazdauskaite, V.V. Borutinskaite and R. Navakauskiene, 2012. DNA methyltransferase inhibitor RG108 and histone deacetylase inhibitors cooperate to enhance NB4 cell differentiation and E-cadherin re-expression by chromatin remodelling. Cell Biol. Int., 36: 1067-1078.
CrossRefDirect Link - Schwechter, B.R., L.E. Millet and L.A. Levin, 2007. Histone deacetylase inhibition-mediated differentiation of RGC-5 cells and interaction with survival. Investigat. Ophthalmol. Visual Sci., 48: 2845-2857.
CrossRefDirect Link - Segal, R.A., 2003. Selectivity in neurotrophin signaling: Theme and variations. Ann. Rev. Neurosci., 26: 299-330.
CrossRefDirect Link - Bai, S., K. Ghoshal, J. Datta, S. Majumder, S.O. Yoon and S.T. Jacob, 2005. DNA methyltransferase 3b regulates nerve growth factor-induced differentiation of PC12 cells by recruiting histone deacetylase 2. Mol. Cell. Biol., 25: 751-766.
CrossRefDirect Link - Stresemann, C., B. Brueckner, T. Musch, H. Stopper and F. Lyko, 2006. Functional diversity of DNA methyltransferase inhibitors in human cancer cell lines. Cancer Res., 66: 2794-2800.
CrossRefPubMedDirect Link - Struzyna, L.A., J.P. Harris, K.S. Katiyar, H.I. Chen and D.K. Cullen, 2015. Restoring nervous system structure and function using tissue engineered living scaffolds. Neural Regeneration Res., 10: 679-685.
CrossRefDirect Link - Sun, S., Y. Han, J. Liu, Y. Fang and Y. Tian et al., 2014. Trichostatin A targets the mitochondrial respiratory chain, increasing mitochondrial reactive oxygen species production to trigger apoptosis in human breast cancer cells. PloS One, Vol. 9.
CrossRef - Tamplenizza, M., C. Lenardi, E. Maffioli, S. Nonnis and A. Negri et al., 2013. Nitric oxide synthase mediates PC12 differentiation induced by the surface topography of nanostructured TiO2. J. Nanobiotechnol., Vol. 11.
Direct Link - Tedeschi, A., T. Nguyen, R. Puttagunta, P. Gaub and S. Di Giovanni, 2009. A p53-CBP/p300 transcription module is required for GAP-43 expression, axon outgrowth and regeneration. Cell Death Differentiation, 16: 543-554.
CrossRefDirect Link - Tinnes, S., M.K.E. Schafer, A. Flubacher, G. Munzner, M. Frotscher and C.A. Haas, 2011. Epileptiform activity interferes with proteolytic processing of Reelin required for dentate granule cell positioning. FASEB J., 25: 1002-1013.
CrossRefPubMedDirect Link - Weeber, E.J., U. Beffert, C. Jones, J.M. Christian, E. Forster, J.D. Sweatt and J. Herz, 2002. Reelin and ApoE receptors cooperate to enhance hippocampal synaptic plasticity and learning. J. Biol. Chem., 277: 39944-39952.
CrossRefDirect Link - Wu, C.L., Y.H. Chou, Y.J. Chang, N.Y. Teng, H.L. Hsu and L. Chen, 2012. Interplay between cell migration and neurite outgrowth determines SH2B1 β-enhanced neurite regeneration of differentiated PC12 cells. PloS One, Vol. 7.
CrossRef - Zhang, Y., H.J. Klassen, B.A. Tucker, M.T.R. Perez and M.J. Young, 2007. CNS progenitor cells promote a permissive environment for neurite outgrowth via a matrix metalloproteinase-2-dependent mechanism. J. Neurosci., 27: 4499-4506.
CrossRefDirect Link - Zhang, Y., D.B. Moheban, B.R. Conway, A. Bhattacharyya and R.A. Segal, 2000. Cell surface Trk receptors mediate NGF-induced survival while internalized receptors regulate NGF-induced differentiation. J. Neurosci., 20: 5671-5678.
PubMedDirect Link