
Jutti Levita
Faculty of Pharmacy, Universitas Padjadjaran, Jatinangor, West Java, Indonesia
As ari Nawawi
School of Pharmacy, Institut Teknologi Bandung, West Java, Indonesia
Abdul Mutalib
Center for Radioisotope and Radiopharmaca, National Nuclear Agency, Serpong, Indonesia
Slamet Ibrahim
School of Pharmacy, Institut Teknologi Bandung, West Java, Indonesia
International Journal of Pharmacology
Year: 2010 | Volume: 6 | Issue: 5 | Page No.: 569-576







ABSTRACT
This study focuses on the anti-inflammatory activity of andrographolide, a diterpenoid compound from Andrographis paniculata, that have been observed in animals and in vitro in different cells of human and animals. Studies included activities of andrographolide and the features of the target, especially limited to transcription factors NF-kappaB. The active site of NF-kappaB, called DNA binding region is found to have mainly positive potential. In order to have an electrostatic complementarity specific inhibitor should have negative potential. Andrographolide has three hydrogen bond donors (H atoms in hydroxyl groups attached to C-3, C-19 and C-14), five hydrogen bond acceptors (O atoms in hydroxyl groups attached to C-3, C-19 and C-14, carbonyl and lactone) and log P-value 2.9. It fulfills Lipinski’s rules of five criteria of drug properties. This compound has different mechanisms of anti-inflammatory activity. It can inhibit the activation of NF-kappaB, suppresses inducible nitric oxide synthase (iNOS) expression. It also prevents oxygen radical production by human neutrophils and inhibits COX-2 expression in human fibroblast cells. This compound also exerts anticancer and antitumour activities, hepatoprotective against various inducers, immunomodulator, antioxidant, antidiabetic, antimicrobe and antivirus activities. Synthetic analogues of the compound which have been created and analyzed also showed similar activities.
PDF Abstract XML References Citation
Received: March 08, 2010;
Accepted: May 12, 2010;
Published: July 14, 2010
How to cite this article
Jutti Levita, As ari Nawawi, Abdul Mutalib and Slamet Ibrahim, 2010. Andrographolide: A Review of its Anti-inflammatory Activity via Inhibition of NF-kappaB Activation from Computational Chemistry Aspects. International Journal of Pharmacology, 6: 569-576.
DOI: 10.3923/ijp.2010.569.576
URL: https://scialert.net/abstract/?doi=ijp.2010.569.576
DOI: 10.3923/ijp.2010.569.576
URL: https://scialert.net/abstract/?doi=ijp.2010.569.576
INTRODUCTION
Andrographolide, a bioactive compound of Andrographis paniculata (Burm.F) Nees, is the major labdane diterpenoidal constituent in this plant. This plant has been used traditionally in the South East Asia countries, India and China to treat various diseases. Andrographolide which is a very bitter compound was isolated in its pure form and characterized for the first time by Gorter at 1911. The structure of andrographolide has been analyzed by X-ray crystallographic method. Its systematic name is 3-[2-[decahydro-6-hydroxy-5-(hydroxymethyl)-5,8a-dimethyl-2-methylene-1-naphthalenyl] ethylidine]dihydro-4-hydroxy-2(3H)-furanone (Smith et al., 1982).
Andrographolide (Fig. 1a) has an α-alkylidene γ-butyrolactone, two olefin bonds at C-8 and C-12 and three hydroxyls at C-3, C-19 and C-14 (Nanduri et al., 2004). Its molecular formula is C20H30O5. This compound has many bioactivities including anti-inflammatory via different mechanisms (Chiou et al., 2000; Shen et al., 2002; Satyanarayana et al., 2004; Wang et al., 2004; Xia et al., 2004; Hidalgo et al., 2005; Sheeja et al., 2006; Abu-Ghefreh et al., 2009; Bao et al., 2009; Li et al., 2009; Suebsasana et al., 2009; Chandrasekaran et al., 2010; Chao et al., 2010; Levita et al., 2010), anticancer and antitumour (Rajagopal et al., 2003; Satyanarayana et al., 2004; Shen et al., 2009; Varma et al., 2009; Lee et al., 2010; Tan et al., 2010; Zhou et al., 2010), hepatoprotective against various inducers (Handa and Sharma, 1990a, b; Visen et al., 1993; Koul and Kapil, 1994), immunomodulator (Wang et al., 2010), antioxidant (Sheeja et al., 2006; Akowuah et al., 2008; Lin et al., 2009), antidiabetic (Zhang et al., 2009), antimicrobe (Xu et al., 2006) and antivirus (Calabrese et al., 2000; Chen et al., 2009). Andrographolide also activates human bitter taste receptor hTAS2R50 (Behrens et al., 2009).
Andrographolide is soluble in methanol, ethanol, pyridine, acetic acid and acetone, but it is slightly dissolved in ether and water. Its melting point is 228-230°C and its ultraviolet λmax in ethanol is 223 nm. This compound can be extracted from the leaves of Andrographis paniculata (Burm.F) Nees by employing methanol as solvent using standard soxhlet method as well as supercritical carbon dioxide extraction. The best extraction condition occurred at 10 MPa, 40°C and a flow rate of 2 mL min-1 for a 3 g sample of Andrographis paniculata ground-dried leaves. The measured extraction rate was found to be about 0.0174 g of andrographolide gram-1 of andrographolide present in the leaves h-1 of operation (Kumoro and Hasan, 2007).
Other constituents contained in the plant, known as andrographolide’s analogues were 14-deoxyandrographolide, 14-deoxy-11,12-didehydroandrographolide, 14-deoxy-11-oxoandrographolide and neoandrographolide. A few years later Fujita and colleagues reexamined the constituents of the plant and isolated andrographolide, neoandrographolide, 14-deoxyandrographolide and three new diterpenoids, which were andrograpanin andropanoside and 14-deoxy-12-methoxy-andrographolide (Fujita et al., 1984).
Andrographolide andrographiside and neoandrographolide were investigated on the hepatocellular antioxidant defense system in carbon tetrachloride-treated mice and resulted that intraperitonial administration of the three diterpenes at a dose of 100 mg kg-1 b.wt. for seven days caused an increasing of cellular antioxidant defense and a decreasing in lipid peroxidation. The data indicated that the antioxidant activity might play an important role in their antihepatotoxic activity (Koul and Kapil, 1994).
Structure of andrographolide: The structure of andrographolide (Fig. 1a) has been analyzed by X-ray crystallographic method and the proposed systematic name is 3-[2-[decahydro-6-hydroxy-5-(hydroxymethyl)- 5,8a-dimethyl-2-methylene-1-naphthalenyl] ethylidine] dihydro-4-hydroxy-2(3H)-furanone (Smith et al., 1982). The Carbon-13 NMR spectrum of andrographolide in C5D5N solvent showed the chemical shifts (δ) of C-1 (δ 37.6 ppm), C-2 (δ 29.1 ppm), C-3 (δ 80.1 ppm), C-4 (δ 43.4 ppm), C-5 (δ 55.6 ppm), C-6 (δ 24.5 ppm), C-7 (δ 38.3 ppm), C-8 (δ 148.1 ppm), C-9 (δ 56.6 ppm), C-10 (δ 39.4 ppm), C-11 (δ 25.1 ppm), C-12 (δ 146.8 ppm), C-13 (δ 130.2 ppm), C-14 (δ 66.2 ppm), C-15 (δ 75.2 ppm), C-16 (δ 170.5 ppm), C-17 (δ 108.7 ppm), C-18 (δ 23.7 ppm), C-19 (δ 64.2 ppm) and C-20 (δ 15.3 ppm). The values were recorded at 50 MHz and tetramethylsilane was used as the standard (Fujita et al., 1984).
Based on computational study using AutoDock and MOE softwares andrographolide is concluded as a flexible ligand which has eleven conformers and six rotatable bonds (Levita et al., 2009b). This flexibility allows andrographolide to be able to interact with different targets.
Andrographolide’s three dimensional structure (Fig. 1b) has been examined, calculated and stored in PubChem database. This compound which has molecular weight of 350.4492 g mol-1, three hydrogen bond donors (H atoms in hydroxyl groups attached to C-3, C-19 and C-14), five hydrogen bond acceptors (O atoms in hydroxyl groups attached to C-3, C-19 and C-14, carbonyl and lactone) and log P value 2.9, fulfills Lipinski’s drug properties (Daisy et al., 2009).
Understanding the interactions between proteins and ligands is crucial for the pharmaceutical industries. The experimental structures of these protein-ligand complexes are usually obtained by techniques such as X-ray crystallography or NMR.
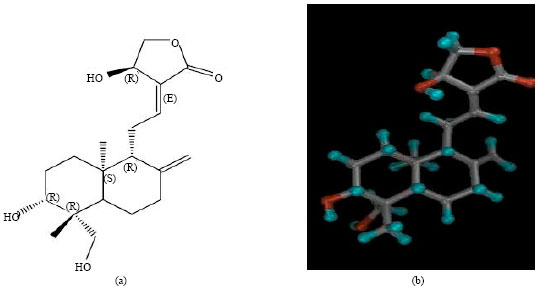 | |
| Fig. 1: | (a, b) 2D and 3D structure of andrographolide (2D structure was sketched using ChemOffice 2004 and visualized using AutoDock http://www.autodock.scripps.edu/) |
To overcome the limitation of predicting the interactions between thousands of ligands and one receptor, the use of computational algorithms (i.e., docking algorithms) have been developed (Daisy et al., 2009).
Andrographolide’s anti-inflammatory activity: Andrographolide has different mechanisms of anti-inflammatory activity. It can inhibit the activation of NF-kappaB, suppresses inducible nitric oxide synthase (iNOS) expression, inhibits COX-2 expression in human fibroblast cells and also prevents oxygen radical production by human neutrophils (Chiou et al., 2000; Shen et al., 2002; Satyanarayana et al., 2004; Wang et al., 2004; Xia et al., 2004; Hidalgo et al., 2005; Sheeja et al., 2006; Abu-Ghefreh et al., 2009; Bao et al., 2009; Li et al., 2009; Suebsasana et al., 2009; Chandrasekaran et al., 2010; Chao et al., 2010; Levita et al., 2010). Andrographolide is also able to modulate T cell activation both in vitro as well as in vivo. The exact mechanism by which andrographolide exhibits its beneficial effect on EAE is still not fully elucidated, but the researchers provide evidence it could prevent initial T cell priming by interfering with DC maturation and antigen presentation capacity. Therefore, andrographolide may have utility as a therapeutic agent for the treatment of autoimmune diseases, such as multiple sclerosis (Iruretagoyena et al., 2005). Recent study shows that andrographolide interacts with Arg513 and His90 in the cyclooxygenase site of COX-2 and inhibits PGE2 production in human fibroblast cells (IC50 = 4 μM) (Levita et al., 2010).
Inhibition of NF-kappaB: NF-kappaB comprises a family of inducible transcription factors that serve as important regulators of the host immune and inflammatory response. The NF-kappaB transcription factor regulates expression of various components of the immune system including proinflammatory cytokines, chemokines, adhesion molecules and inducible enzymes such as cycloxygenase-2 and inducible nitric oxide synthase, as well as proteins that regulate the specific immune response, such as interleukin (IL)-2, IL-12 and interferon-γ that control lymphocyte proliferation and differentiation. Disregulation of this transcription factor can thus lead to inflammatory and autoimmune diseases (Yamamoto and Gaynor, 2001).
The specific amino acids that are referred to as the DNA Binding Region of NF-kappaB (Fig. 2b) are residues Arg59, Tyr60, Val61, Cys62, Glu63, Gly64, Pro65, Ser66, His67, Gly68, Gly69, Leu70 and Pro71 of the subunit p50. DBR of the p50 subunit is found to have mainly a positive potential (Pande et al., 2003).
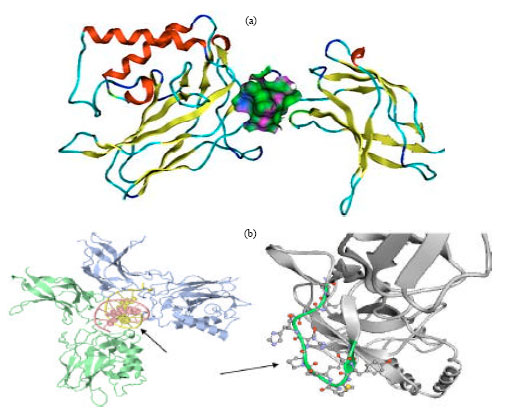 | |
| Fig. 2: | (a) Visualization of sub unit p50 NF-kappaB and (b) DNA binding region of NF-kappaB is showed by black arrows (NF-kappaB pdb file 1NFK was downloaded from http://www.pdb.org and visualized using AutoDock http://www.autodock.scripps.edu/) |
This potential is strongly positive on the external surface which is the first accessible surface for any ligand or the target DNA. Further, the major amino acids including Arg56, His67 and Ser66, are also having a net positive potential around them. In order to have an electrostatic complementarity with the DNA binding region, specific inhibitors should preferably have a negative potential around them (Pande et al., 2003; Levita et al., 2009b).
Sub unit p50 NF-kappaB which has been separated from its homodimeric form, shows molecular surface of DNA Binding Region (DBR) which is located at the middle of the protein (Fig. 2a). Green color visualizes the hydrophobic region of the DBR, while magenta and blue colors represent hydrophilic and polar regions (Levita et al., 2009b).
Molecular modelling study on different classes of p50-DNA binding inhibitors gave a good prediction about their possible structural activity. By making strong hydrogen bonds, active inhibitors may be masking important amino acids like arginine, histidine and serine, which are important in DNA binding. The NF-kappaB dimer interface exhibits hydrophobic interactions and hydrogen bonds. Two unique hydrogen bonds are present in the NF-kappaB p50-p65 heterodimer that distinguish it from the p50 and p65 homodimers. The homologous residues, Asp-254 (from p50) and Asn-200 (from p65), form a hydrogen bond between their O-δ and N-δ atoms. This contact is unique and observed only in the p50-p65 heterodimer but not in the respective homodimers, which may contribute to the stability of the heterodimer. Similarly, a new hydrogen bond is formed when N-∈2 from His-304 contacts S-γ in Cys-197. In p50, Arg-252, Glu-265, Tyr-267, Cys-270, Arg-305 and Phe-307 also interact with the p65 subunit through hydrogen bonds. These residues contact Asp-243, Glu-211, Arg-198, Val-199, Asn-200, His-245, Val-248, Leu-215, Cys-216 and Asp-217 in p65 (Berkowitz et al., 2002). Andrographolide can be placed in the DBR p50 NF-kappaB and makes four hydrogen bondings with Arg51, Asp206, Asn244 and Asp336, with its scoring value is -9.9863 kcal mol-1 which represents its docking energy. The same interactions are also showed by aurine tricarboxylic acid (-12.1801 kcal mol-1) and gallic acid (-12.4534 kcal mol-1), known inhibitors of p50 NF-kappaB (Levita et al., 2009b).
Andrographolide has been proven to attenuate inflammation by inhibition of NF-kappaB activation through covalent modification of reduced Cys62 of p50. Mechanistically, it formed a covalent adduct with reduced cysteine (62) of p50, thus blocking the binding of NF-kappaB oligonucleotide to nuclear proteins.
Andrographolide suppressed the activation of NF-kappaB in stimulated endothelial cells, which reduced the expression of cell adhesion molecule E-selectin and prevented E-selectin mediated leukocyte adhesion under flow. It also abrogated the cytokine-and endotoxin-induced peritoneal deposition of neutrophils, attenuated septic shock and prevented allergic lung inflammation in vivo. Notably, it had no suppressive effect on IkappaBα degradation, p50 and p65 nuclear translocation, or cell growth rates (Xia et al., 2004).
Other researchers analyzed the effect of andrographolide on the activation of NF-kappaB induced by Platelet-Activating Factor (PAF) and N-formyl-methionyl-leucyl-phenylalanine (fMLP) in HL-60 cells differentiated to neutrophils. PAF (100 nM) and fMLP (100 nM) induced activation of NF-kappaB as determined by degradation of inhibitory factor B α (IkBα) using Western blotting in cytosolic extracts and by binding to DNA using Electrophoretic Mobility Shift Assay (EMSA) in nuclear extracts. Andrographolide had been proven inhibited the NF-kappaB luciferase activity induced by PAF. However andrographolide did not reduce phosphorylation of p38 MAPK or ERK1/2 and did not change IkappaBα degradation induced by PAF and fMLP. It also reduced the DNA binding of NF-kappaB in whole cells and in nuclear extracts induced by PAF and fMLP. It is concluded that andrographolide exerts its anti-inflammatory effects by inhibiting NF-kappaB binding to DNA and thus reducing the expression of proinflammatory proteins, such as COX-2 (Hidalgo et al., 2005).
To understand further about the molecular recognition of inhibitors by NF-kappaB, another researchers analyzed 15-deoxy-D12,14-prostaglandin J2 (15d-PGJ2), an electrophilic prostaglandin, which is a dehydration product of prostaglandin D2 (PGD2) by using docking studies. 15d-PGJ2 was reported to inhibit multiple steps in the NF-kappaB signaling pathway along with specifically inhibiting the NF-kappaB subunits (p50 and p65), directly by a covalent modification of the subunits (Straus et al., 2000). The study indicated that 15d-PGJ2 forms a covalent adduct with the aid of its electrophilic carbon with single critical cysteine residues, which are Cys62 in the p50 subunit and Cys38 in the p65 subunit. The docking studies in combination with comparative electrostatic potentials reveal that 15d-PGJ2 has complementary interactions with its target proteins, governed mainly by fit of shape (hydrophobic and van der Waals interactions) in all the cases and also complementarity of electrostatics (Pande and Ramos, 2005).
The effects of nuclear factor kappa B (NF-kappaB) inhibition on the secretion of macrophage migration inhibitory factor (MIF) in human CD4(+) T cells was also examined. The results indicated that pharmacological inhibition of NF-kappaB causes the release of MIF through de novo synthesis of MIF and the secretion of preformed MIF in CD4(+) T cells through the production of reactive oxygen species (Cho et al., 2009).
This compound also reduced NFAT luciferase activity and interfered with its nuclear distribution, with these effects being linked to an increase in c-jun-N-terminal kinase (JNK) phosphorylation. Additionally, reduction of NF-kappaB activity in Jurkat cells treated with andrographolide was observed. Using Western blotting, Carretta, M.D and colleagues demonstrated that andrographolide decreased ERK1 and ERK5 phosphorylation induced by anti-CD3 or PMA/Ionomycin. Andrographolide did not affect cell viability at concentration of 10 and 50 muM; however, their results suggested that andrographolide increased early apoptosis at 100 muM. They concluded that andrographolide could exert immunomodulatory effects by interfering with NFAT activation and ERK1 and ERK5 phosphorylation in T-cells (Carretta et al., 2009).
Suppression of inducible nitric oxide synthase (iNOS) expression: In pathological conditions, macrophages produce both NO and superoxide anion simultaneously, resulting in the formation of ONOO¯, which, through further reaction, can exert even stronger oxidant effects (Ischiropoulos et al., 1992). Therefore, high amounts of NO potentially cause cytotoxicity and capable of injuring the surrounding cells and tissues indiscriminately either by itself or by formation of ONOO¯.
Andrographolide has been reported to exhibit Nitric Oxide (NO) inhibitory property in endotoxin-stimulated macrophages, however, the detailed mechanisms remain unclear. This compound inhibits NO synthesis in RAW 264.7 cells by reducing the expression of iNOS protein and the reduction could occur through two additional mechanisms which are: prevention of the de novo protein synthesis and decreasing the protein stability via a post-transcriptional mechanism. It is also possible that inhibition of iNOS protein expression and NO production under immune stimulation and/or bacteria infection may explain, in part, the beneficial effects of andrographolide as an anti-inflammatory agent. It is worth noting that the inhibitory activity of andrographolide lasted for 18 h after LPS stimulation. These data suggest that andrographolide acts more like a protein synthesis inhibitor (Chiou et al., 2000).
Prevention of oxygen radical production: The anti-inflammatory effect of andrographolide could be explained by its ability to inhibit neutrophil adhesion/transmigration through suppression of Mac-1 upregulation. The inhibitory effect of andrographolide on Mac-1 expression could be mediated by down regulation of ROS production via a PKC-dependent but calcium independent mechanism (Shen et al., 2002).
Inhibition on PAF-induced platelet aggregation: Andrographolide was investigated for its suggested influence on the biosynthesis of eicosanoids and the Platelet-Activating Factor (PAF). Whereas in isolated human Polymorph-Nuclear Leukocytes (PMNL) no influence on the biosynthesis was found, it could be shown that andrographolide inhibits PAF-induced human blood platelet aggregation in a dose dependent manner (IC50 = 5 μM). These results indicate that andrographolide has a mechanism of action different from that of Non-Steroidal Antiinflammatory Drugs (NSAID) and most likely associated with the cardiovascular and antithrombotic activity described of Andrographis paniculata (Amroyan et al., 1999).
Inhibition of COX-2 expression: Andrographolide and its analogue, neoandrographolide a minor diterpenoid compound contained in Andrographis paniculata (Burm.F) Nees, had been proven interacted with Arg513 and His90 in the cyclooxygenase site of COX-2. The energy needed for the interactions are relatively small andrographolide -11.7963 kcal mol-1 and neoandrographolide -7.4339 kcal mol-1 (calculated using AutoDockTools 3.0.5). These results indicates that the interactions are quite favourable and will happen spontaneously (Levita et al., 2009a, b, 2010).
The inhibitory activity of andrographolide to COX-2 enzyme which was determined by measuring PGE2 production in human fibroblast cells stimulated with LPS resulted an IC50 = 4 μM which was 0.7 times of acetosal’s. These data confirmed that andrographolide’s anti-inflammatory activity also occured via inhibition of COX-2 expression (Levita et al., 2010).
REFERENCES
- Abu-Ghefreh, A.A., H. Canatan and C.I. Ezeamuzie, 2009. In vitro and in vivo anti-inflammatory effects of andrographolide. Int. Immunopharmacol., 9: 313-318.
CrossRefDirect Link - Akowuah, G.A., I. Zharik and A. Mariam, 2008. Analysis of urinary andrographolides and antioxidant status after oral administration of Andrographis paniculata leaf extract in rats. Food Chem. Toxicol., 46: 3616-3620.
PubMed - Amroyan, E., E. Gabrielian, A. Panossian, G. Wikman and H. Wagner, 1999. Inhibitory effect of andrographolide from Andrographis paniculata on PAF-induced platelet aggregation. Phytomedicine, 6: 27-31.
PubMed - Bao, Z., S. Guan, C. Cheng, S. Wu and S.H. Wong et al., 2009. A novel antiinflammatory role for andrographolide in asthma via inhibition of the nuclear factor-kB pathway. Am. J. Respir. Crit. Care Med., 179: 657-665.
PubMed - Behrens, M., A. Brockhoff, C. Batram, C. Kuhn, G. Appendino and W. Meyerhof, 2009. The human bitter taste receptor hTAS2R50 is activated by the two natural bitter terpenoids andrographolide and amarogentin. J. Agric. Food Chem., 57: 9860-9866.
Direct Link - Berkowitz, B., D.B. Huang, F.E. Chen-Park, P.B. Sigler and G. Ghosh, 2002. The X-ray crystal structure of the NF-_B p50_p65 heterodimer bound to the interferon beta-kappa B site. J. Biol. Chem., 277: 24694-24700.
PubMed - Calabrese, C., S.H. Berman, J.G. Babish, X. Ma and L. Shinto et al., 2000. A phase I trial of andrographolide in HIV positive patients and normal volunteers. Phytother. Res., 14: 333-338.
PubMedDirect Link - Carretta, M.D., P. Alarcon, E. Jara, L. Solis and J.L. Hancke et al., 2009. Andrographolide reduces IL-2 production in T-cells by interfering with NFAT and MAPK activation. Eur. J. Pharmacol., 602: 413-421.
PubMed - Chandrasekaran, C.V., A. Guptaa and A. Agarwal, 2010. Effect of an extract of Andrographis paniculata leaves on inflammatory and allergic mediators in vitro. J. Etnopharmacol., 129: 203-207.
PubMed - Chao, W.W., Y.H. Kuo and B.F. Lin, 2010. Anti-inflammatory activity of new compounds from Andrographis paniculata by NF-kappaB transactivation inhibition. J. Agric. Food Chem., 58: 2505-2512.
Direct Link - Chen, J.X., H.J. Xue, W.C. Ye, B.H. Fang and Y.H. Liu et al., 2009. Activity of andrographolide and its derivatives against influenza virus in vivo and in vitro. Biol. Pharm. Bull., 32: 1385-1391.
PubMed - Chiou, W.F., C.F. Chen and J.J. Lin, 2000. Mechanisms of suppression of inducible nitric oxide synthase (iNOS) expression in RAW 264.7 cells by andrographolide. Br. J. Pharmacol., 129: 1553-1560.
CrossRef - Cho, M.L., Y.M. Moon, Y.J. Heo, Y.J. Woo and J.H. Ju et al., 2009. NF-kappa B inhibition leads to increased synthesis and secretion of MIF in human CD4+ T cells. Immunol. Lett., 123: 21-30.
PubMed - Daisy, P., R. Sasikala and A. Ambika, 2009. Analysis of binding properties of phosphoinositide 3-kinase through in silico molecular docking. J. Proteomics Bioinform., 2: 274-284.
Direct Link - Fujita, T., R. Fujitani, Y. Takeda, Y. Takaishi, T. Yamada, M. Kido and I. Miura, 1984. On the diterpenoids of Andrographis paniculata: X-ray crystallographic analysis of andrographolide and structure determination of new minor diterpenoids. Chem. Pham. Bull., 32: 2117-2125.
Direct Link - Handa, S.S. and A. Sharma, 1990. Hepatoprotective activity of andrographolide from Andrographis paniculata against carbon tetrachloride. Indian J. Med. Res., 92: 276-283.
PubMedDirect Link - Handa, S.S. and A. Sharma, 1990. Hepatoprotective activity of andrographolide against galactosamine and paracetamol intoxication in rats. Ind. J. Med. Res., 92: 284-292.
PubMed - Hidalgo, M.A., A. Romero, J. Figueroa, P. Cortes, I.I. Concha, J.L. Hancke and R.A. Burgos, 2005. Andrographolide interferes with binding of nuclear factor-kB to DNA in HL-60-derived neutrophilic cells. Br. J. Pharmacol., 144: 680-685.
CrossRef - Iruretagoyena, M.I., J.A. Tobar, P.A. Gonzalez, S.E. Sepulveda and C.A. Figueroa et al., 2005. Andrographolide interferes with T-Cell activation and reduces experimental autoimmune encephalomyelitis in the mouse. J. Pharmacol. Exp. Therapeutics, 312: 366-372.
Direct Link - Koul, I.B. and A. Kapil, 1994. Effect of diterpenes from Andrographis paniculata on antioxidant defense system and lipid peroxidation. Ind. J. Pharmacol., 26: 296-300.
Direct Link - Kumoro, A.C. and M. Hasan, 2007. Supercritical carbon dioxide extraction of andrographolide from Andrographis paniculata: Effect of the solvent flow rate, pressure, and temperature. Chin. J. Chem. Eng., 15: 877-883.
CrossRefDirect Link - Lee, Y.C., H.H. Lin, C.H. Hsu, C.J. Wang, T.A. Chiang and J.H. Chen, 2010. Inhibitory effects of andrographolide on migration and invasion in human non-small cell lung cancer A549 cells via down-regulation of PI3K/Akt signaling pathway. Eur. J. Pharmacol., 632: 23-32.
CrossRef - Levita, J., E.P. Istyastono, A. Nawawi, A. Mutholib, I.J.P. de Esch and S. Ibrahim, 2009. Analyzing the interaction of andrographolide and neoandrographolide, diterpenoid compounds from Andrographis paniculata (Burm.F) nees, to cyclooxygenase-2 enzyme by docking simulation. ITB J. Sci., 41: 110-119.
CrossRefDirect Link - Levita, J., A. Nawawi, A. Mutholib and S. Ibrahim, 2010. Andrographolide inhibits COX-2 expression in human fibroblast cells due to its interaction with arginine and histidine in cyclooxygenase site. J. Applied Sci., 10: 1481-1484.
CrossRefDirect Link - Lin, F.L., S.J. Wu, S.C. Lee and L.T. Ng, 2009. Antioxidant, antioedema and analgesic activities of andrographis paniculata extracts and their active constituent andrographolide. Phytother. Res., 23: 958-964.
Direct Link - Nanduri, S., V.K. Nyavanandi, S.S.R. Thunuguntla, S. Kasu and M.K. Pallerla et al., 2004. Synthesis and structure-activity relationship of andrographolide analogues as cytotoxic agents. Bioorg. Med. Chem. Lett., 14: 4711-4717.
CrossRef - Pande, V., R.K. Sharma, I. Jun-Ichiro, M. Otsuka and M.J. Ramos, 2003. A molecular modeling study of inhibitors of nuclear factor-kappa B (p50)- DNA binding. J. Comput. Aided Mol. Design, 17: 825-836.
Direct Link - Pande, V. and M.J. Ramos, 2005. Molecular recognition of 15-deoxy-D12,14-prostaglandin J2 by nuclear factor-kappa B and other cellular proteins. Bioorganic Med. Chem. Lett., 15: 4057-4063.
CrossRef - Rajagopal, S., R.A. Kumar, D.S. Deevi, C. Satyanarayana and R. Rajagopalan, 2003. Andrographolide, a potential cancer therapeutic agent isolated from Andrographis paniculata. J. Exp. Ther. Oncol., 3: 147-158.
CrossRef - Satyanarayana, C., D.S. Deevi, R. Rajagopalan, N. Srinivas and S. Rajagopal, 2004. DRF 3188 a novel semi-synthetic analog of andrographolide: Cellular response to MCF7 breast cancer cells. BMC Cancer, 4: 1-8.
Direct Link - Sheeja, K., P.K. Shihab and G. Kuttan, 2006. Antioxidant and anti-inflammatory activities of the plant andrographis paniculata nees. Immunopharmacol. Immunotoxicol., 28: 129-140.
PubMed - Shen, K.K., T.Y. Liu, C. Xu, L.L. Ji and Z.T. Wang, 2009. Andrographolide inhibits hepatoma cells growth and affects the expression of cell cycle related proteins. Yao Xue Xue Bao, 44: 973-979.
PubMed - Shen, Y.C., C.F. Chen and W.F. Chiou, 2002. Andrographolide prevents oxygen radical production by human neutrophils: Possible mechanisms involved in its anti-inflammatory effect. Br. J. Pharmacol., 135: 399-406.
CrossRef - Smith, A.B., B.H. Toder, P.J. Carroll and J. Donohue, 1982. Andrographolide: An x-ray crystallographic analysis. J. Chem. Crystallography, 12: 309-319.
Direct Link - Suebsasana S., P. Pongnaratorn, J. Sattayasai, T. Arkaravichien, S. Tiamkao and C. Aromdee, 2009. Analgesic, antipyretic, anti-inflammatory and toxic effects of andrographolide derivatives in experimental animals. Arch. Pharm. Res., 32: 1191-1200.
Direct Link - Tan, Y., K.H. Chiow, D. Huang and S.H. Wong, 2010. Andrographolide regulates epidermal growth factor receptor and transferrin receptor trafficking in epidermoid carcinoma (A-431) cells. Br. J. Pharmacol., 159: 1497-1510.
Direct Link - Varma, A., H. Padh and N. Shrivastava, 2009. Andrographolide: A new plant-derived antineoplastic entity on horizon. eCAM Adv. Access.
CrossRef - Visen, P.K.S., B. Shukla, G.K. Patnaik and B.N. Dhawan, 1993. Andrographolide protects rat hepatocytes against paracetamol-induced damage. J. Ethnopharmacol., 40: 131-136.
CrossRefDirect Link - Wang, T., B. Liu, W. Zhang, B. Wilson and J.S. Hong, 2004. Andrographolide reduces inflammation-mediated dopaminergic neurodegeneration in mesenphalic neuronglia cultures by inhibiting microglial activation. J. Pharmacol. Exp. Therapeutics, 308: 975-983.
PubMed - Wang, W., J. Wang, S.F. Dong, C.H. Liu and P. Italiani et al., 2010. Immunomodulatory activity of andrographolide on macrophage activation and specific antibody response. Acta Pharmacol. Sin., 31: 191-201.
Direct Link - Xia, Y.F., B.Q. Ye, Y.D. Li, J.G. Wang and X.J. He et al., 2004. Andrographolide attenuates inflammation by inhibition of NF-kB activation through covalent modification of reduced cysteine62 of p50. J. Immunol., 173: 4207-4217.
Direct Link - Xu, Y., R.L. Marshall and T.K.S. Mukkur, 2006. An investigation on the antimicrobial activity of Andrographis paniculata extracts and andrographolide in vitro. Asian J. Plant Sci., 5: 527-530.
CrossRefDirect Link - Yamamoto, Y. and R.B. Gaynor, 2001. Therapeutic potential of inhibition of the NF-kappaB pathway in the treatment of inflammation and cancer. J. Clin. Invest., 107: 135-142.
CrossRefPubMedDirect Link - Zhang, Z., J. Jiang, P. Yu, X. Zeng, J.W. Larrick and Y. Wang, 2009. Hypoglycemic and beta cell protective effects of andrographolide analogue for diabetes treatment. J. Trans. Med., 7: 62-62.
CrossRef - Zhou, J., C.N. Ong, G.M. Hur and H.M. Shen, 2010. Inhibition of the JAK-STAT3 pathway by andrographolide enhances chemosensitivity of cancer cells to doxorubicin. Biochem. Pharmacol., 79: 1242-1250.
CrossRef - Li, J., L. Luo, X. Wang, B. Liao and G. Li, 2009. Inhibition of NF-kappaB expression and allergen-induced airway inflammation in a mouse allergic asthma model by andrographolide. Cell. Mol. Immunol., 6: 381-385.
PubMed