
Stanton G. Belford
Department of Curriculum and Teaching, 5040 Haley Center, Auburn University, Auburn, AL. 36849, USA
Dawn A.T. Phillip
Department of Life Sciences, St. Augustine, The University of the West Indies, Trinidad, West Indies
International Journal of Oceanography and Marine Ecological System
Year: 2012 | Volume: 1 | Issue: 3 | Page No.: 67-75







ABSTRACT
Zoanthids are integral components of coral reef benthic communities in the southern Caribbean. Although reef-building scleractinian corals are important in providing the foundational structure of coral reefs, the dynamic nature of current trends in climate change will definitely affect cnidarian distribution and may promote competitive hardier species to increase in their abundance and distribution on coral reefs. Currently, two common zoanthid sp. (Palythoa caribaeorum and Zoanthus sociatus) are distributed more in some zones than others when compared to their reef-building cnidarian counterparts. The distribution of zoanthids was assessed for 2008 and 2012 in four horizontal intertidal zones using the line and point transect method. Intertidal scleractinian and zoanthid cover was 16.8 and 36.7%, respectively in 2008, compared to scleractinian (17.2%) and zoanthid cover (48.7%) in 2012. Distribution of zoanthids significantly increased in zones 2 and 4 from 2008-2012, as a result of increased coverage for Z. sociatus and P. caribaeorum respectively. Scleractinian coverage change was negligible in all zones, compared to increased zoanthid coverage, especially in zones 2 and 4 in 3.5 years. This study provides the first quantitative zoanthid distribution data over a short time period illustrating the dynamic changes in distribution favoring zoanthids over their more famous scleractinian benthic components of coral reef communities.
PDF Abstract XML References Citation
Received: June 05, 2012;
Accepted: June 11, 2012;
Published: November 08, 2012
How to cite this article
Stanton G. Belford and Dawn A.T. Phillip, 2012. Intertidal Distribution Patterns of Zoanthids Compared to their Scleractinian Counterparts in the Southern Caribbean. International Journal of Oceanography and Marine Ecological System, 1: 67-75.
DOI: 10.3923/ijomes.2012.67.75
URL: https://scialert.net/abstract/?doi=ijomes.2012.67.75
DOI: 10.3923/ijomes.2012.67.75
URL: https://scialert.net/abstract/?doi=ijomes.2012.67.75
INTRODUCTION
One of the features very common to coral reef communities is the countless interactions and associations among various species in their ecosystem. The dynamic nature of these communities is dependent on coral species distribution and abundance, which influences changes in the benthic component of reefs. Coral reef benthic biota plays a fundamental role on the overall reef ecology (Mendonca-Neto et al., 2008). For example, cnidarian distribution on reefs is affected by sediment and this influences overall species diversity on these reef systems (Hoegh-Guldberg, 1999). Rivers are the main source of sediment input to the reef (Wilkinson, 2002; Mallela and Perry, 2007). Additionally, rivers carry nutrient input, which are other types of terrestrial run-off that affect coral distribution (Goreau, 1992; Wilkinson et al., 1994).
Nutrient poor tropical waters contain dynamic coral communities teeming with diversity (Muscatine et al., 2005). This marine ecosystem diversity is only second to tropical rainforests as the most diverse ecosystem in the world (Souter and Linden, 2000). Unfortunately, it has been suggested that there is a global and steady decline in reef habitat due to coral bleaching and ocean acidification which are triggered by current trends in global climate change (Bruno and Selig, 2007; Hoegh-Guldberg, 2011). Furthermore, one-third of Caribbean reefs are affected by coastal development and pollution from inland sources (Wilkinson, 2004). Altogether, a total of 60% of reefs in the Caribbean are affected by overfishing (Wilkinson, 2004; Burke and Maidens, 2004).
Success in these nutrient-poor waters lies in the symbiotic relationship between corals and their endosymbiotic dinoflagellates known as zooxanthellae (genus Symbiodinium; Muscatine and Porter, 1977). Zooxanthellae have been shown to release up to 95% of their photosynthetic products to their hosts in return for the metabolic waste products; nitrogen and phosphate (Muscatine, 1990; Garrison, 2005). Consequently, corals just require ample space to grow because their obligate bond with their symbiotic associates account for their successful survival. Ultimately, corals compete for space to grow and some species have the tendency to out-compete other species. Buss and Jackson (1979) suggested that competition influences marine hard bottom structure and diversity. Competitive coral species can gain advantages in substratum by acquiring open patches by invasion, overgrowing neighbors and moving onto dead surfaces of other organisms (Connell, 1978; Keough, 1985).
There is a plethora of knowledge on reef-building coral distribution throughout the Caribbean. Reef-building corals belong to the order Scleractinia (Class Anthozoa: Phylum Cnidaria). However, a lack of information, particularly quantitative descriptions, exists for non-scleractinian corals. One hexacorallian group that has not been widely investigated even though it is a major component of reef communities in the southern Caribbean are the zoanthids (Order Zoanthidea or Zoantharia: Cnidaria: Hexacorallia).
Zoanthids are small, generally colonial, anemone-like cnidarians (Wood and Wood, 2000; Reimer et al., 2008) that can grow as dense mats or small patches in both shallow and deep parts of coral reefs. They have circular oral disks surrounded by two rings of short tentacles (Humann and Deloach, 2002). Zoanthids are found worldwide, but their species diversity is relatively unknown (Ryland and Muirhead, 1993; Burnett et al., 1997; Reimer et al., 2007). There are two zoanthid species common throughout the intertidal zone on reef systems located on the north eastern coast of Trinidad (Belford and Phillip, 2011): P. caribaeorum and Z. sociatus (Perez et al., 2005).
P. caribaeorum can be found throughout the Atlantic (Haywick and Mueller, 1997) and is known to be common in Brazilian reefs (Perez et al., 2005). Their colonies are highly competitive and show high tolerance to environmental change. Karlson (1983) showed that zoanthids may need continual natural disturbances in order to maintain their dominance on reefs in Discovery Bay, Jamaica. Currently, there is a lack of quantitative data on the distribution of Z. sociatus and the effects of competition for space by P. caribaeorum in the southern Caribbean. Additionally, more long-term quantitative data are needed to monitor reef communities on the northeastern coast of Trinidad, which are affected by nutrients and sediments brought by the South Equatorial current.
Coral reef monitoring in Trinidad and Tobago has thus far been mostly restricted to reefs in Tobago. For example, Buccoo Reef on Tobago is a government protected area and is economically valuable to the tourism industry. The island of Tobago has many famous dive sites and one of these sites has the largest known symmetrical brain coral (Diploria strigosa) in the world. In comparison, there are no protected reefs surrounding the island of Trinidad. Studies adding quantitative baseline data to be used in future proposals to assist in stimulating policy makers to implement protection ordinances that will maintain the sustainability of marine habitats located on the northeastern coasts of Trinidad are desperately needed.
The island of Trinidad is the closest Caribbean island to South America. Reef systems along the northeastern Trinidad coasts are affected by wind, currents, rainfall and sediment discharge from local and South American sources. The South Equatorial current distributes freshwater from the South American mainland to the coastal shores of Trinidad and Tobago. This water contains nutrients, organic matter and sediment from the Amazon and Orinoco Rivers which affects seasonal changes in turbidity and salinity (Nansingh and Jurawan, 1999; Spalding et al., 2001; Mallela and Harrod, 2008).
The main goal of this study was to quantify intertidal zoanthid distribution at sites along the northeastern coast of Trinidad over 2 periods in time, (1) August 2008 and (2) January 2012 and to determine if changes in zoanthid and scleractinian percentage cover over the years favored any species.
MATERIALS AND METHODS
Study area: The two sites were located on the northeastern coast of Trinidad (located between 10°-11°N, 61°-62°W), close to the two largest South American rivers (Amazon and Orinoco). These rivers heavily influence the study reefs through contributions of freshwater, nutrients and sediments. The Caribbean Sea surrounds the study areas. Both sites in the Toco region are on the north coast, within ~4.0 km from Galeota Point which is the extreme northeastern point of Trinidad (Fig. 1).
Both study sites were investigated between two periods in time, (1) August 2008 and (2) January 2012. Toco Bay (TB) and Baptist Bay (BB) (10°50.107`N to 10°50.266`N) are adjacent to each other, located towards the eastern end of the island next to the Toco Fishing Depot. Transect points were previously marked and served as points where coral cover data were collected (Belford and Phillip, 2011).
Field survey: The line and point transect method was used to estimate intertidal benthos distribution and abundance. Transect lines were made from brightly colored yellow polypropylene line (40 m) with colored surveyors tape at 1 m intervals.
 | |
| Fig. 1: | Map of the Caribbean (inset) showing the study sites surveyed in August 2008 and January 2012 along the northeastern coast of Trinidad |
The line was stretched perpendicular to the shore towards the reef and each reef component (biota and non-biota) were tallied under 1 m intervals along the transect line (Belford and Phillip, 2011). A total of 8 transect points, 50 m apart were measured from the west to east at both TB and BB. Transect points were set at 50 m intervals and were marked by spray painting an entry point on terrestrial substrate (trees and walls). A hand-held Global Positioning System (GPS) and photographs were also used to record the position of each transect point to provide location confirmation during subsequent visits.
Four intertidal zones were used to represent distances from the maximum tide imprint left on the shore (hereafter shoreline) during low tide. Zone 1 was 0-10 m from the shoreline and zone 2 (11-20 m), 3 (21-30 m) and 4 (31-40 m) were respectively designated. Coral abundances were determined as percentage coral cover of 4 zones on transect lines. Scleractinian coverage of reef-building Porites porites and Siderastrea radians were calculated for each zone and compared to total zoanthid (P. caribaeorum and Z. sociatus) cover for 2008 and 2012.
Statistical analysis: Data were collected during daytime falling tides (mean low tide depth 0.30-0.35 m) and water surface temperature was also measured using a hand-held digital thermometer. Local tide tables were used to determine the lowest tides of the month. These low tides provided easy access to collect data on benthic reef components using the line and point transect method.
RESULTS
P. caribaeorum forms brown mat-like colonies but can appear as solitary polyps (Fig. 2a, b). Polyps are separated by a thin coenenchyme (P. caribaeorum). Zoanthid species were observed in both solitary and colony forms and in particular P. caribaeorum was seen growing as mats throughout the reef benthos at Toco Bay (TB) and Baptist Bay (BB) and also at a fringing reef located at Salybia Bay approximately 2 km from TB. Although, Z. sociatus often include a diverse array of colors including gray, blue, purple and orange (Fig. 2c, d), they mostly appear as green mats throughout various sites of the northeastern Toco region of Trinidad (Fig. 2e, f).
Percentage cover of reef-building corals such as Porites porites and Siderastrea radians have not changed as much for a period of approximately 3.5 years. There were slight increase in coral cover in zones 2 and 3, compared to similar decreases in zones 1 and 4 (Fig. 3a). Surprisingly, the average hard coral cover for zones 1-4 in August 2008 was 16.8% compared to 17.2% in January 2012.
Zoanthid sp. showed increased coverage in all zones for the 3.5 year time period. However, the most noticeable increase occurred in zones 2 and 4 (17.5% and 14.5% respectively, Fig. 3b). These values were the result of an 11.3% increase in coverage for Z. sociatus in zone 2 (Fig. 3c) and a 16.7% increase for P. caribaeorum in zone 4 (Fig. 3d). Interestingly, there wasn’t that much change in cover for Z. sociatus in all zones, except zone 2, compared to a similar pattern for P. caribaeorum in all zones except zone 4.
Overall, although reef-building corals showed increase in coverage only for zones 2 and 3, zoanthids showed a steady increase in all zones, with higher coverage in zones 2 and 4 compared to their reef-building scleractinian counterparts. Consequently, zoanthid percentage cover was greater than twice that of reef-builders in all zones examined in January 2012, compared to similar examination in August 2008 (Fig. 3a, b).
 | |
| Fig. 2(a-f): | The two common zoanthid species were visibly growing in either solitary polyps or forming extensive mats in all zones as shown in, (a) P. caribaeorum growing as solitary polyps at Toco Bay, (b) Salybia Bay and displayed various color morphotypes, (c) Orange color morphotype of Z. sociatus, (d) Brown morph, (e) In various shades of green at Toco Bay and (f) In the reef within the immediate vicinity of the Toco Lighthouse |
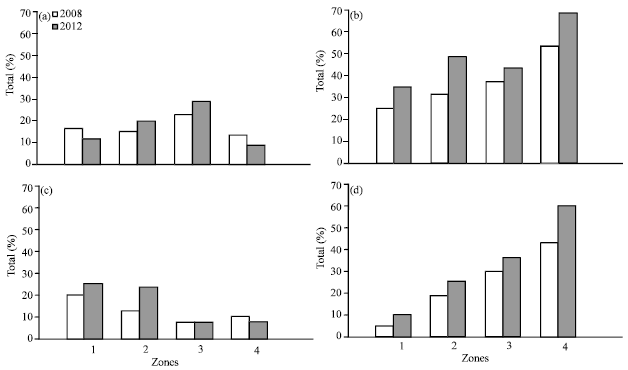 | |
| Fig. 3(a-d): | Intertidal coral cover in zones 1-4 for combined Toco and Baptist Bays during August 2008 and January 2012, (a) Reef-builders, (b) Zoanthids, (c) Z. sociatus and (d) P. caribaeorum |
DISCUSSION
The results of this study showed favorable growth for zoanthids in all 4 intertidal zones compared to their scleractinian counterparts over a short period of time (3.5 years). The noticeable increased cover in zones 2 and 4 correlated with either a decrease or only a slight increase in scleractinian cover in these zones. Interestingly, although the only indication of decreased coverage was reported for Z. sociatus in zone 4, its counterpart, P. caribaeorum had a noticeable increase in this zone and favorable growth in all zones. Past research by Belford and Phillip (2011) has shown initial scleractinian cover at 13.3% and more recently 17.2% (this study) with an overall 6 year increase in coral cover of just 3.9%. Consequently, zoanthid growth is more favorable as shown from its steady increase in cover and it will be interesting to see if this pattern continues in the wake of dynamic climate change.
Additionally, the results of this study add the first quantitative comparison of benthos cover for zoanthid species and reef-building corals growing at Toco and Baptist Bays, on the northeast coast of Trinidad, in the southern Caribbean. Currently, there is a need for long-term data due to the inexistence of an official reef-monitoring program in the area. Both sites are ~2.5 km away from Salybia Bay (the best developed fringing reef in Trinidad) and long-term effects of climate change and human influence may affect all coral ecosystems on the northeastern coast of Trinidad. Ultimately, quantitative baseline data in this study can be used to encourage the implementation of a reef-monitoring program to follow annual patterns in cnidarian abundance and distribution.
The rocky intertidal nature at TB partly restricted the growth of reef-builders in 2008 and 2012. The combination of high energy and turbidity in conjunction with the closeness of the upper surface of the rocks at low tide levels is a limiting growth factor of scleractinians. Ultimately, high energy and shallowness favors zoanthid growth because they are better adapted to strong wave action and exposure. Scleractinian coral cover maintained consistent coverage in all zones, compared to noticeable zoanthid increases in a few zones for the 3.5 year period of time. An increase in P. caribaeorum growth in zone 4 and approximately consistent growth of Z. sociatus in zone 2 provided an illustration of the dynamic nature of zoanthid cover on the coral reef. P. caribaeorum cover in zone 4 correlated with a decrease in scleractinian cover as noted for the time period.
P. caribaeorum was the most abundant cnidarian species at the outer intertidal zone close to the reef crest at all sites including Salybia Reef (Belford and Phillip, 2011), whereas Z. sociatus was more abundant nearer the shoreline in zones 1 and 2. In Venezuela, Bastidas and Bone (1996) found similar high abundances of P. caribaeorum in shallow fore reef zones where this species formed massive brown slippery mats while Z. sociatus dominated nearest to the shore. Bastidas and Bone (1996) further suspected that P. caribaeorum can over-grow Z. sociatus, since P. caribaeorum exhibited faster initial growth after cover removal, which did not favor Z. sociatus. Karlson (1983) noticed a decrease in Z. sociatus cover when he investigated the effects of storm, due to the overturning of rocks and shading. Shading occurs in certain parts of the reef near to the shore at Toco and Baptist Bays, thus may be one reason why many types of Z. sociatus color morphotypes can be seen closer to the shoreline (Belford and Phillip, 2011).
Unlike many corals, zoanthids are common on shallow reef flats in the Caribbean and Bahamas (Wood and Wood, 2000). Intertidal sections of reef flats on the northeastern coast of Trinidad show similar zoanthid distributions, but abundances are different within the intertidal zone. At TB and BB, Z. sociatus mean cover was higher closer to the shore (0-15 m), compared to higher coverage of P. caribaeorum in zones 3 and 4, that is, further away from the shore. Cnidarians such as beaded anemones, Epicystis crucifer, lesser starlet coral, Siderastrea radians, finger coral, Porites porites, fire coral, Millepora alcicornis and algae were less common than Z. sociatus 0-15 m from the shore. Zoanthids at both sites covered rocks, boulders and coral rubble. Both zoanthid species are exposed to the atmosphere at spring tides for at least 3.5 hours for three days each month. This exposure may contribute to the “hardiness” of zoanthid species in this region, due to inconsistent fluctuations in surface and water temperatures but this is entirely speculative.
The low percentage scleractinian cover at the sites in this study is representative of reef benthos seen throughout the Caribbean. Scleractinian (reef-building) corals are the fundamental unit of reef ecosystems (Souter and Linden, 2000). Major Caribbean reef-builders such as Acropora palmata, Acropora cervicornis and Montastraea annularis have declined in abundance over the last two decades (Mumby and Steneck, 2008). Such declines have been attributed to changing climate, disease (Hoegh-Guldberg, 1999; Hughes et al., 2003; Mumby and Steneck, 2008) and anthropogenic factors such as coastal development, which has affected 30% of the world’s reefs (Bryant et al., 1998).
The impact of using reefs to educate local high school, undergraduate students and communities living in close proximity to reefs extends towards being better stewards of our environment. This study can be used to present data to government authorities, as well as local stakeholders to illustrate the need to implement long-term reef monitoring initiatives in the area. Furthermore, knowledge of coral species distribution can be used to determine long-term changes, which may be on the horizon as climate change becomes more apparent.
ACKNOWLEDGMENTS
We thank Dr. James Reimer for valuable comments throughout the development of this paper. We also thank Dr. A. Ramsubhag and Grace Surju-Charran (UWI) for their support. We acknowledge Mr. Winston Montano for geological advice about the study sites and Dr. Dennis Mullen at Middle Tennessee State University (MTSU) for his advice on this project. This project was funded by a J. Gerald Parchment Biological Field Station scholarship (MTSU), Wayne Price Biological Research fund and Martin Methodist College.
REFERENCES
- Bastidas, C. and D. Bone, 1996. Competitive strategies between Palythoa caribaeorum and Zoanthus sociatus (Cnidaria: Anthozoa) at a reef flat environment in Venezuela. Bull. Mar. Sci., 59: 543-555.
Direct Link - Belford, S. and D. Phillip, 2011. Rapid assessment of a coral reef community in a marginal habitat in the Southern Caribbean: A simple way to know what's out there. Asian J. Biol. Sci., 4: 520-531.
CrossRefDirect Link - Bruno, J.F. and E.R. Selig, 2007. Regional decline of coral reef cover in the Indo-Pacific: timing, extent and subregional comparisons. PLoS One, Vol. 2.
CrossRef - Burnett, W.J., J.A.H. Benzie, J.A. Beardmore and J.S. Ryland, 1997. Zoanthids (Anthozoa, hexacorallia) from the Great Barrier Reef and Torres Strait, Australia: Systematics, evolution and a key to species. Coral Reefs, 16: 55-68.
CrossRef - Buss, L.W. and J.B. Jackson, 1979. Competitive networks: Nontransitive competitive relationships in cryptic coral reef environments. Am. Nat., 113: 223-234.
Direct Link - Connell, J.H., 1978. Diversity in tropical rain forests and coral reefs. Sci., 199: 1302-1310.
CrossRefDirect Link - Goreau, T.J., 1992. Bleaching and reef community change in Jamaica: 1951-1991. Am. Zool., 32: 683-695.
CrossRefDirect Link - Haywick, D. and E. Mueller, 1997. Sediment retention in encrusting Palythoa spp.: A biological twist to a geological process. Coral Reefs, 16: 39-46.
CrossRef - Hoegh-Guldberg, O., 1999. Climate change, coral bleaching and the future of the world's coral reefs. Mar. Freshwater Res., 50: 839-866.
CrossRefDirect Link - Hoegh-Guldberg, O., 2011. Coral reef ecosystems and anthropogenic climate change. Regional Environ. Change, 11: 215-227.
CrossRef - Karlson, R., 1983. Disturbance and monopolization of spatial resource by Zoanthus sociatus (Coelenterata, Anthozoa). Bull. Mar. Sci., 33: 118-131.
Direct Link - Mallela, J. and C. Harrod, 2008. δ13C and δ15N reveal significant differences in the coastal foodwebs of the seas surrounding Trinidad and Tobago. Mar. Ecol. Prog. Ser., 368: 41-51.
Direct Link - Mallela, J. and C. Perry, 2007. Calcium carbonate budgets for two coral reefs affected by different terrestrial runoff regimes, Rio Bueno, Jamaica. Coral Reefs, 26: 129-145.
Direct Link - Mendonca-Neto, J.P., C.E.L. Ferreira, L.C.T. Chaves and R.C. Pereira, 2008. Influence of Palythoa caribeorum (Anthozoa, Cnidaria) zonation on site-attached reef fishes. Ann. Bra. Acad. Sci., 80: 495-513.
CrossRef - Muscatine, L. and J.W. Porter, 1977. Reef corals: Mutualistic symbioses adaptive to nutrient-poor environments. Bioscience, 27: 454-460.
Direct Link - Muscatine, L., 1990. The role of symbiotic algae in carbon and energy flux in reef corals. Coral Reefs, 25: 1-29.
Direct Link - Muscatine, L., C. Goiran, L. Land, J. Jaubert, J.P. Cuif and D. Allemand, 2005. Stable isotopes (delta C-13 and delta N-15) of organic matrix from coral skeleton. Proc. Natl. Acad. Sci. USA, 102: 1525-1530.
CrossRef - Nansingh, P. and S. Jurawan, 1999. Environmental sensitivity of a tropical coastline (Trinidad, West Indies) to oil spills. Spill. Sci. Technol. B, 5: 161-172.
CrossRef - Perez, C., D.A. Vila-Nova and A.M. Santos, 2005. Associated community with the zoanthid Palythoa caribaeorum (Duchassaing and Michelotti, 1860) from littoral of Pernambuco, Brazil. Hydrobiologia, 548: 207-215.
CrossRef - Reimer, J.D., K. Takishita, S. Ono and T. Maruyama, 2007. Diversity and evolution in the zoanthid genus Palythoa (Cnidaria: Hexacorallia) based on nuclear ITS-r DNA. Coral Reefs, 26: 399-410.
CrossRef - Reimer, J.D., M. Nonaka, F. Sinniger and F. Iwase, 2008. Morphological and molecular characterization of a new genus and new species of parazoanthid (Anthozoa: Hexacorallia: Zoantharia) associated with Japanese Red Coral. Coral Reefs, 27: 935-949.
CrossRef - Souter, D. and O. Linden, 2000. The health and future of coral reef systems. Ocean Coastal Manage., 43: 657-688.
CrossRefDirect Link - Spalding, M., C. Ravilious and E.P. Green, 2001. World Atlas of Coral Reefs. University of California Press, California, ISBN: 9780520232556, Pages: 424.
Direct Link - Wilkinson, C.R., R.W. Buddemeier and UNEP-IOC-ASPEI-IUCN, 1994. Global Climate Change and Coral Reefs: Implications for People and Reefs. IUCN, Gland, Switzerland, ISBN: 9782831702049, Pages: 124.
Direct Link - Wilkinson, C., 2004. Status of Coral Reefs of the World: 2004. Vol. 4, Australian Institute of Marine Sciences, Townsville, Australia, Pages: 255.
Direct Link