
S.M.A. Ghazal
Department of Genetics and Cytology, Genetic Engineering and Biotechnology Division, National Research Center, Cairo, Egypt
W.S. Elsayed
Department of Microbiology, Faculty of Sciences, Ain Shams University, Cairo, Egypt
U.M. Badr
Department of Genetics and Cytology, Genetic Engineering and Biotechnology Division, National Research Center, Cairo, Egypt
H.M. Gebreel
Department of Microbiology, Faculty of Sciences, Ain Shams University, Cairo, Egypt
K.M.A. Khalil
Department of Genetics and Cytology, Genetic Engineering and Biotechnology Division, National Research Center, Cairo, Egypt
Current Research in Bacteriology
Year: 2011 | Volume: 4 | Issue: 2 | Page No.: 44-62







ABSTRACT
Xanthomonas campestris is very important industrial microorganism for its ability to produce xanthan gum which has multiple applications. X. campestris was genetically modified to increase its β-galactosidase content for subsequent growth on Whey as cheep cultivation medium and concomitant industrial infection of xanthan. Therefore, suicidal plasmid pSUP5011 was mobilized from E. coli to X. campestris by transconjugation to introduce the transposon Tn5. About 126 X. campestris transconjugants Kmr were selected and tested for its ability to produce xanthan gum in whey media. Strain TC 49, the highest xanthan producer transconjugant, was subjected to UV-irradiation for further genetic modification enhancing xanthan over production. About 45 mutants were selected and tested for their ability to produce xanthan in whey media. A new genetically modified mutant, Tn-UV-2 proved to be the best xanthan producer in lactose or whey media. Produced xanthan gum was characterized chemically using Fourier Transform Infrared (FTIR) spectra and was found to have the same FTIR spectral peaks of xanthan gum produced commercially.
PDF Abstract XML References Citation
Received: February 23, 2011;
Accepted: April 13, 2011;
Published: August 01, 2011
How to cite this article
S.M.A. Ghazal, W.S. Elsayed, U.M. Badr, H.M. Gebreel and K.M.A. Khalil, 2011. Genetically Modified Strains of Xanthomonas campestris Higher Xanthan Producer and Capable to Utilize Whey. Current Research in Bacteriology, 4: 44-62.
DOI: 10.3923/crb.2011.44.62
URL: https://scialert.net/abstract/?doi=crb.2011.44.62
DOI: 10.3923/crb.2011.44.62
URL: https://scialert.net/abstract/?doi=crb.2011.44.62
INTRODUCTION
Xanthan gum is widely used in a broad range of industries, such as in foods as a gelling and stabilizing agent in salad dressings, ice creams, also in toiletries, oil recovery, cosmetics, as to confer the required properties without affecting the taste of the final product (Jain-Raina and Babbar, 2011). In the agriculture industry, xanthan has been used to improve the flow-ability in fungicides, herbicides and insecticides formulations. It is also used in jet injection printing. In the petroleum industry, xanthan gum is used in oil drilling, fracturing and pipeline cleaning (Sarbatly and England, 2006; Nasr et al., 2007). Xanthomonas campestris, the phytopathogenic Gram-negative bacterium, responsible for a large number of economically important plant diseases is known for its ability to produce xanthan gum (Rottava et al., 2009). Xanthan gum is a water-soluble extracellular polysaccharide. It is the most principal microbial polysaccharide of industrial importance and is the only bacterial exopolysaccharides regarded by the Food and Drug Adminstration as safe food additive (Katzen et al., 1996). About 20,000 tons of xanthan are produced industrially from X. campestris each year. Many researchers have studied the fermentation conditions required for optimal gum production where 4% sucrose or glucose medium with a suitable nitrogen source gives the best gum yield (Giannouli and Morris, 2003). Gum production by X. campestris is genetically controlled by gene cluster composed of 12 genes designated gum B, -C, -D, -E, -F, -G, -H, -I, -J, -K, -L and -M. The transcriptional organization of this gene cluster was analyzed and indicated that such a gene cluster region was mainly expressed as an operon from a promoter located upstream of the first gene, gum B (Katzen et al., 1998; Vojnov et al., 1998). The most significant advances in microbial genetics are based on the study and use of mutations which can be induced either by chemical or physical mutagens such as Ultraviolet (UV) which is a non-ionizing form of radiation used to induce mutation in bacteria (Oppezzo and Pizarro, 2003; Belov et al., 2009). Or through transposon mutagenesis by transposable elements which are distinct DNA segments have the unique capacity to move (transpose) to new sites within the genome of their host organisms. Success in the isolation of transposon mutants with the desirable phenotypes depends on the availability of an efficient system for the delivery of specific transposon into the bacterium under study. Whey, a by-product of dairy industries, is a fluid containing very low quantities of milk solids and it possesses high biological value due to its consistency in lactose and proteins. The composition of whey is variable but generally includes 4 to 5% lactose, 0.8 to 1% protein and smaller amounts of organic acids, minerals and vitamins. However, Xanthomonas campestris pv. campestris possesses a low level of β-galactosidase and therefore, is not able to grow and produce significant amounts of xanthan gum in medium containing lactose as the sole carbon source, though does not produce large amounts of xanthan gum from whey (Silva et al., 2009; Fu and Tseng, 1990; Ekateriniadou et al., 1994). Therefore, many attempts has been made to solve this problem as done by El-Sawah and Microbiology (1999) also another approach to solve this problem is to manipulate X. campestris isolate for hydrolyzing lactose more efficiently and high xanthan synthesis. Some attempts have introduced for the production of xanthan in lactose or whey medium, e.g., by using X. campestris genetically engineered strain through using transposable elements and other microbial biotechnology techniques. The objective of this study was to obtain a new strain (s) of Xanthomonas campestris producing more quantities of xanthan gum and has/have the ability to grow on whey.
MATERIALS AND METHODS
Bacterial strains and plasmid: The present study was conducted between 2006-2009. The strain of X. campestris pv. campestris was obtained from Institute of Molecular Biology, National Chung Hsing Univ., China. The strain of E. coli k12 S17-1 harboring the plasmid pSUP5011::Tn5-mob was obtained from German culture collection DSMZ (DSM No. 5167, Germany).
Growth media and inoculum preparation: The preservation media used were LB media (Bertani, 1951) and Yeast Malt (YM) media (Roseiro et al., 1992). Maintenance media including Glucose Yeast extract Calcium carbonate Medium (GYCA) and Lactose Yeast extract Calcium carbonate Medium (LYCA) (Dye et al., 1980). Inocula were prepared according to Garcia-Ochoa et al. (2000). Commercial sweet powder whey was obtained from local market (Ukraine) to be supplemented in the culture media. Fermentation media and fermentation conditions were designed according to Garca-Ochoa et al. (1992).
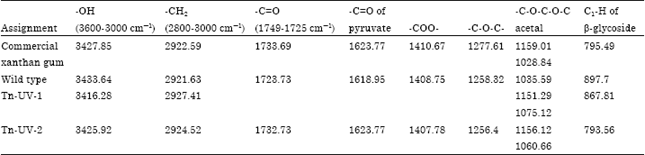
| Table 1: | FTIR spectral data (cm-1) of commercial xanthan gum, wild type strain (WT1), Tn-UV-1 and Tn-UV-2 |
 | |
Transconjugation of X. campestris: For conjugational transfer of plasmid, a procedure developed by Gerhardt et al. (1994) was used. Transconjugants were recognized on GYCA and LYCA medium supplemented with Kanamycin (Kmr). Negative controls were prepared by plating donor and recipient strains separately on selective media. Transconjugants were screened for plasmid DNA.
Ultraviolet treatment of X. campestris: UV-irradiation was used to induce mutations in X. campestris using Philips T-UV-30 W lamp type number 57413 p/40, according to (Al-Turki et al., 2008).
Xanthan production and purification: Precipitation and purification of xanthan gum was performed according to Cadmus et al. (1978). Relative viscosity of xanthan was determined viscometerically using conventional Ostwald viscometer. The viscometer was attached to water bath to maintain the temperature at 25°C. The relative viscosity to water of the sample solution was measured and calculated according to Page and Sadoff (1975).
Isolation of plasmid DNA: Plasmid DNA was extracted using Quick and easy kit for bacterial plasmids isolation (Khalil, 2007). Samples were electrophresed on 0.5% agarose in 1X TAE buffer. Gels were stained with ethidium bromide and examined by UV transilluminator.
Characterization of xanthan gum using FTIR spectroscopy: FTIR spectra were recorded on a Jasco FTIR 6100, Japan. The dry samples of the wild type strain, commercial xanthan gum, Tn-UV-1 and Tn-UV-2 were mixed with KBr and pressed into discs under reduced pressure. The FTIR spectra were obtained by scanning range between 4000 cm and 400 cm-1 and the data was listed in Table 1.
RESULTS
Culture conditions and fermentation media for xanthan production: Xanthan gum production by X. campestris was tested on the selected fermentation media supplemented with different carbon sources (lactose, sucrose and glucose) and the relative viscosity was estimated in relative to blank media, the results showed the lowest relative viscosity obtained on fermentation media contained lactose as it was 2.8 comparing with the highest one on sucrose 13.52 while the relative viscosity on glucose-containing media was 1.14 (Fig. 1a-c).
β-galactosidase gene improvement in X. campestris: Suicidal plasmid pSUP5011 was mobilized from E. coli k12 S17-1 as a donor to X. campestris as a recipient by transconjugation to introduce the transposon Tn 5.
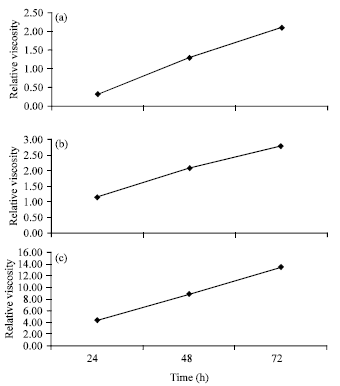 | |
| Fig. 1: | Relative viscosity of wild type strain on fermentation medium with different carbon sources; (a) glucose (b) Lactose and (c) Sucrose |
All available Ampr, Kanr and Nmr transconjugated colonies were isolated and characterized. After transconjugation, 126 transconjugants were selected as Kmr only. All of them tested for there ability to produce xanthan through determining Relative Viscosity using the four carbon sources, sucrose, glucose, lactose and whey compared with Wild Type (WT) strain. The results were shown in Table 2 as the relative viscosity of the wild type strain and 126 transconjugants.
Statistical analysis of bacterial transconjugation: The result showed xanthan production by 126 transconjugants compared with WT1 production using four different carbon sources (Table 2). They divided into three levels low, same and higher according to their production relative to WT1 production (Fig. 2). From the results no isolates showed the same production level of WT1 because all these transconjugants were isolated as a Kmr and contain a Tn5 transposable element. This variation in genetic material makes transconjugants genetically different from WT1 which reflects their production.
Gene environmental interaction between transconjugants and glucose as carbon source: From 124 transconjugants tested for their ability to utilize glucose for xanthan production, non of them showed the same level of WT1 production. While 109 transconjugants were lower producers than mother strain (equal 88% of all tested transconjugants), 15 transconjugants were higher producers than mother strain which represent about 12% of all tested transconjugants. Transconjugant No. 11 was the best xanthan producer on glucose containing media.
Gene environmental interaction between transconjugants and sucrose as carbon source: Sucrose is one of the recommended sources of carbon for xanthan production. About 31% of them (39 transconjugants), were less producers than WT1.
| Table 2: | Results of relative viscosity of the wild type strain and 126 transconjugants |
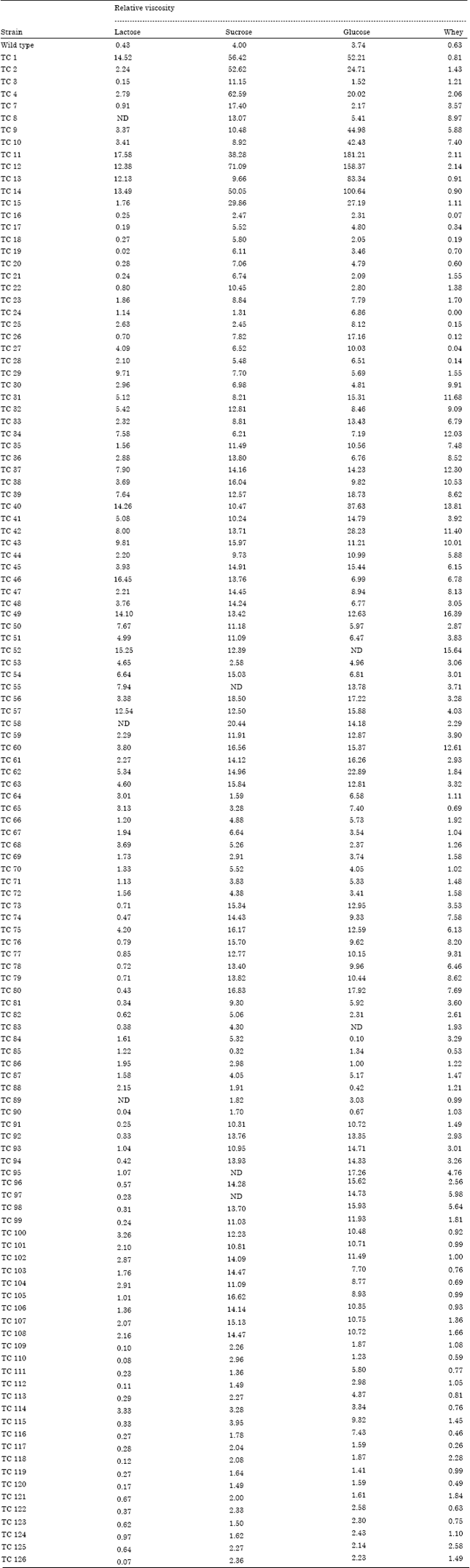 | |
| ND: Not determined | |
More than two third of the transconjugants were higher producers (84 transconjugants). Eight transconjugants were higher producer; they showed from four times to thirteen times production more than WT1.
Gene environmental interaction between transconjugants and Lactose as carbon source: In Lactose medium, 122 transconjugants were tested, only one transconjugants showed the same level of mother strain, about 36% (44 transconjugants) of 122 transconjugants tested using lactose media were less than mother strain and 77 transconjugants were higher producer than the mother strain which represented 63% of all tested transconjugants. However, 18 transconjugants showed more than six times more than WT1 which represent 15% of all tested transconjugants.
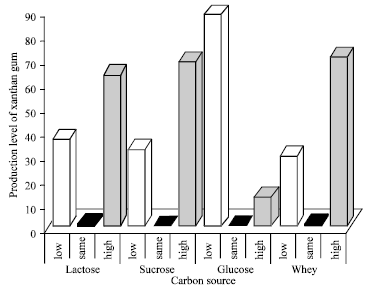 | |
| Fig. 2: | Xanthan production of 126 transconjugants using four different carbon sources, compared with wild type production, divided into three levels, low, same and higher than wild type production |
Gene environmental interaction between transconjugants and whey as carbon source: From 124 transconjugants tested, only one transconjugant showed the same level of xanthan production of WT1 in whey containing medium. While 36 transconjugants (about 29%) were lower producers, 87 transconjugants were higher producers than mother strain which represent 67% of total tested transconjugants. At least 9 transconjugants showed ten times more than WT1. However, only one transconjugant was the best producers, it shows 15 times more than WT1 which reflect its higher ability in utilizing whey for xanthan production.
Characterization of the best xanthan producer transconjugants: After transconjugation, five transconjugants (T 11, 34, 40, 46 and 49) showing the best xanthan production in lactose or whey-containing media compared with wild type strain were enrolled in a single fermentation cycle to unify the fermentation conditions (Fig. 3). In case of sucrose-containing media as a source of carbon, all selected transconjugants were higher xanthan producers than mother strain. Two of them, transconjugant No. 46 and 49 showed two and half folds more than mother strain. Transconjugant No. 11 was the best one it produce seven times more than mother strain. When glucose was used as a source of carbon, three transconjugants (34, 46 and 49) were fewer producers than mother strain. Transconjugant No. 40 produces two times more xanthan than mother strain. The other one, transconjugant No. 11 was the best, it produces over four times more than mother strain in glucose-containing media. In case of lactose and whey, all five transconjugants were higher producer than wild type when lactose and whey were used; transconjugant 11 was the higher xanthan producer when lactose was used as a source of carbon. It produces about eighteenth folds more than mother strain. But it is less producer one when whey was used as a source of carbon. While transconjugant No. 49 was the best xanthan producer in whey containing medium. It produced fifteen folds more than mother strain. The results showed that, transconjugant 11 was the best xanthan producer when lactose, glucose and sucrose were used as a source of carbon and transconjugant No. 49 proved to be the best xanthan producer, when whey was used.
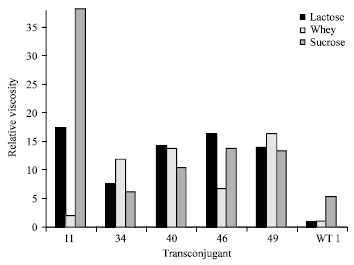 | |
| Fig. 3: | The best five selected transconjugants and their variant ability to utilize three different source of carbon |
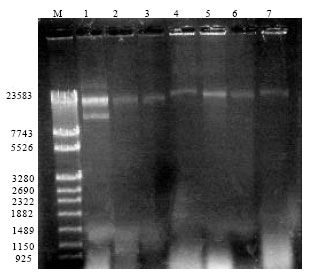 | |
| Fig. 4: | Plasmid pattern of the five selected higher xanthan producing transconjugants compared to E. coli and wild type strain, M, molecular weight marker; 1, E. coli; 2, wild type strain; 3-7, transconjugants 11, 34, 40, 46 and 49, respectively |
Both of them suitable and promising strain, as were shown in Fig. 3.
The five transconjugants were higher xanthan producer than wild type when lactose or whey was used. DNA electrophoresis pattern was performed for the five selected transconjugants showing higher xanthan production compared to wild type strain (Fig. 4).
UV-induced mutation: Transconjugant TC 49 proved to be the best xanthan producer when whey was used. Therefore, TC 49 was selected for second step of genetic improvement for xanthan overproduction through UV treatment. UV-induced mutants were selected as Ampr, Kmr and Nmr and the survival ratios were 3.73, 3.10, 2.11 and 0.08%, respectively after treatment. Mutants were selected as Ampr, Kmr, using LYCA, GYCA media. Followed by isolation of 45 remarkable xanthan producer mutants, each of them were tested for their ability to produce xanthan gum on fermentation media supplemented with different carbon sources: lactose, sucrose and whey.
In order to compare between 45 mutants and their mother strain TC 49 in xanthan production, they were divided into 5 groups according to their xanthan weight production in whey fermentation media. Each group contains nine mutants, a comparison were carried out between mutants productivity inside the same group. Three carbon source; whey, sucrose and lactose were used.
From the previous step, six promising mutants out of 45 were selected according to their xanthan production mainly in whey, they are 49/7/6, 49/9/1, 49/9/12, 49/11/2, 49/11/5 and 49/11/8, as shown in Fig. 5.
The highest xanthan-producers mutants on whey and lactose: From the previous results (Fig. 4) six promising mutants out of 45 were selected according to their xanthan production mainly in whey containing medium.
The selective mutants are, 49/7/6, 49/9/1, 49/9/12, 49/11/2, 49/11/5 and 49/11/8 in addition to mother strain (TC 49) and wild type were also used through this investigation. The results showed that (Fig. 5), mutant 49/9/1 was the second highest xanthan producer in whey and the third higher xanthan producer in lactose with production 10.8 and 9.2 g L-1, respectively. Mutant 49/11/2 proved to be the best xanthan producer when lactose and whey was used as a source of carbon. It produced about 201% compared with wild type and 169% compared with mother strain when lactose was used. Also it reached about 176% xanthan production and 122% compared with wild type and mother strain, respectively in whey containing media. This result indicated that, both mutants showed higher xanthan producer in lactose than whey. Although, four mutant out of six were higher xanthan producer than wild type, they were lower xanthan producer than mother strain (TC 49), when lactose and whey were used in fermentation media. Mutant 49/9/1 was the second highest xanthan producer in whey containing media and the third higher xanthan producer in lactose containing media. Mutant 49/11/2 proved to be the best xanthan producer when lactose was used as a source of carbon. It was also the best mutant in xanthan production in whey containing media. It produced about 201% compared with wild type and 169% compared with mother strain when lactose was used. Also it reached about 176% xanthan production and 122% compared with wild type and mother strain respectively, in whey containing media.
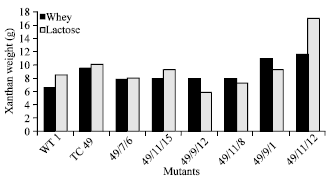 | |
| Fig. 5: | Xanthan production measured as xanthan weights of top six mutants on lactose and whey media |
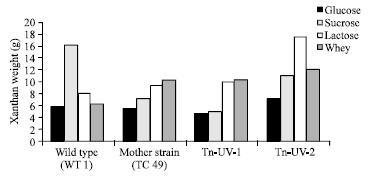 | |
| Fig. 6: | Final xanthan production of the best two mutants, Tn-UV-1 and Tn-UV-2 using four different carbon sources, measured as xanthan weights after seven days of fermentation |
Study of the two selected mutants and the gene environmental conditions interaction for xanthan production: Referring to the previous results showed that the best two mutants were 49/9/1 and 49/11/2 they renamed with Tn-UV-1 and Tn-UV-2, respectively referring to their genetic origin. Tn-UV-1 and Tn-UV-2 mutants subjected for more investigation compared with wild type strain and mother strain TC 49 in order to study the best environmental condition for production, using different carbon sources, glucose, sucrose, lactose and whey were used in this fermentation media (Fig. 6). Both mutants showed the reverse in their behaviors when whey or lactose was used. While mutant Tn-UV-1 showed its maximum xanthan production in whey media with production 10 g L-1, Lactose is the second favorite carbon source to Tn-UV-1 with production 10 g L-1. While whey is the second favorite carbon source to Tn-UV-2 showed 12 g L-1 compared with 17.5 g L-1 in lactose. Although Tn-UV-2 mutant was the second xanthan producer mutant in sucrose media after wild type, it was the best xanthan producer of all tested culture in glucose, lactose and whey containing media showing about 124, 217 and 193%, respectively comparing with the wild type for the same order of carbon source and about 133, 184 and 117%, respectively comparing with the mother strain TC 49 for the same order of carbon source. These two mutants able to utilize lactose or whey with high yield of xanthan gum, one of them Tn-UV-2 produced higher xanthan than wild type in three different carbon sources and higher than mother strain TC 49 in four different carbon sources including glucose and sucrose which considered to be the recommended carbon source proved. Gel electrophoresis and DNA profile was made for the best six mutants and mother strain TC 49, all of them showing a single band of chromosomal DNA. The previous results showed that the best two mutants were 49/9/1 and 49/11/2 they renamed with Tn-UV-1 and Tn-UV-2, respectively referring to their genetic origin.
Characterization of xanthan gum produced from the two selected mutants Tn-UV-1 and Tn-UV-2: Extracted and purified xanthan gum from the selected two mutants along with that from the wild type strain, was characterization by FTIR. The FTIR profile showed the presence of absorption peaks at 900-790 cm-1, characteristic for β-glycoside linkages. On the other hand, the presence of absorption peaks at 1625-1600 cm-1 indicating the presence of pyruvate function and absorption peaks at 1749-1725 cm-1. FTIR of commercial xanthan gum (Mallinckrodt, Paris, Kentucky 40361) compared with wild type strain, Tn-UV-1 and Tn-UV-2 proved that the mutants’ product has the same chemical composition of commercial xanthan gum (Fig. 7).
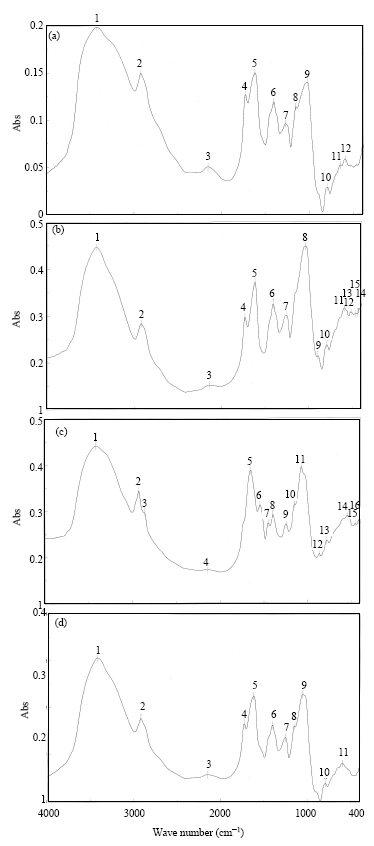 | |
| Fig. 7: | FTIR spectra of (a) commercial xanthan gum (b) wild type (c) Tn-UV-1 and (d) Tn-UV-2 |
DISCUSSION
During the second half of the 20th century, new and useful polysaccharides of medical and commercial interest have been discovered which can be obtained by microbial fermentations.
The usefulness of water-soluble carbohydrate polymers relies on their wide range of functional properties: The ability to modify the properties of aqueous environments that is their capacity to thicken, emulsify, stabilize, flocculate, swell and suspend or to form gels, films and membranes. Another very important aspect that polysaccharides obtained from natural, renewable sources is both biocompatible and biodegradable. Among these polysaccharides, Xanthan gum, a microbial biopolymer produced by Xanthomonas campestris (Hamcerencu et al., 2007; Kar et al., 2010). Present study aimed to induce genetic variation on wild type strain of Xanthomonas campestris to improve the quantification production of xanthan gum through induction of mutation using plasmids harboring transposable elements followed by UV Irradiation then isolation of remarkable producer mutant (s) of xanthan. In the present study a strain of Xanthomonas campestris tested for its xanthan production using three different carbon sources: lactose, sucrose and glucose. The variation in xanthan production of wild type strain has been observed; the results showed that the amount of xanthan produced in fermentation media containing lactose was the least amount compared with the amount of xanthan produced in sucrose-containing media which showed the highest amount. This result confirms the study of many authors such as Frank and Somkuti (1979), Castilho et al. (1984) and Stauffer and Leeder (1978) that the gene coding for β-galactosidase is present in the wild strains of Xanthomonas campestris and has degenerated as the result of the absence of selective pressure in soil and plant environment where lactose is not readily available.
The fermentation media used was recommended by Garcia-Ochoa et al. (2000), this media contained amonium, phosphate and magnesium showed to influence the growth where amonium, phosphate and sulphate showed to influence the production and small amount of organic acids (citric acid) found to enhance the production. Also, inoculums build-up on YM medium is an important step beside using a multiple stages of inoculums development showed a great assistance in xanthan production and these results were supported the work done by Moraine and Rogovin (1971), De Vuyst et al. (1987) and Peters et al. (1989) where they stated that multiple stages of inoculums on YM medium which contained peptone and yeast extracts as a complex organic nitrogen sources rich in proteins and glucose as a carbon source, increased the cell concentration and suppressed the production of xanthan because xanthan around the cell impedes mass transport of nutrients and extends the lag phase of growth as xanthan gum constitute the bacterial capsule.
In the present study emphasis has been imposed on genetic improvement of wild type strain through multiple steps of improvement in order to construct a new strain of Xanthomonas campestris higher xanthan producer and to be capable to utilize whey as a cheap by-product. Many studies have been done on genetic modification and construction of new mutant strains from X. campestris. This study tried to construct a mutant strains through two steps. The first step of genetic improvement was through transconjugation using E. coli k12 S17-1 harboring the plasmid pSUP5011::Tn5-mob. The results showed that all of 126 transconjugants tested for there ability to produce xanthan on fermentation media containing lactose and whey and the best two out of the best five transconjugants were higher producer than wild type when lactose and whey were used; were transconjugant 11 and transconjugant 49 (TC49) suitable and promising strain and (TC49) was used for more genetic improvement as it is produce 15 fold higher than mother strain in whey-containing media.
Although the production of the best transconjugants (TC 49) obtained from this step do not exceed almost 15 fold on whey-containing media and 14 fold in lactose-containing media comparing with the wild type strain, while the production of genetically-constructed lactose utilizing strain obtained from the study of Yang et al. (2002) produce much more (40-200 folds) and this genetically modified strains seem to be superior but the former strain is preferred than the later one. In case of (TC49) the mutation occurred through transposable element which exhibited a random mutagenesis via transconjugation using suicide plasmid; So first using transposable elements Tn5 in induction of random mutation which has low insertion specificity and therefore can insert into a large number of locations in bacterial genomes, as well as into multiple positions within single genes according to De Bruijn and Lupski (1984), gave us a wide scale of selection as many as the places in which the Tn 5 has been inserted. Second using a suicidal plasmid doesn’t replicate in X. campestris and only a delivery vector for Tn5-Mob as supported by De Bruijn and Lupski (1984), De Bruijn (1987), Nies et al. (1987), Bukhari et al. (1977) and Martiel and Blot (2002) limited the fear of spreading an antibiotic resistance genes.
One of the explanations for these differences levels of xanthan production for the same transconjugant between media containing lactose and whey is in lactose the best producer conjugant is that has high ability to utilize lactose as a sole carbon source which reflect a very high beta-galactosidase activity. While in whey, there were other components besides lactose including 4 to 5% lactose, 0.8 to 1% protein and smaller amounts of organic acids, minerals and vitamins these result was in supporting the result from Nahvi and Moeini (2004). Therefore, in whey containing medium, the best producer transconjugant should have remarkable beta-galactosidase activity beside the ability to utilize the other components or even grow in the present of it. This means, in lactose there is only one factor affecting the production. But in whey, besides lactose there are multiple factors affecting xanthan production and the production is the result of interaction between transconjugant and all interfering factors. The other explanation was that all conjugantes prove to have the ability to utilize lactose should have the ability to produce remarkable active beta-galactosidase due to mutation inducted by TN 5, transposable elements. These transconjugants may be differing in its mutation site in DNA but all of them have a high beta-galactosidase activity. When transconjugants cultured on whey, different factors were interacting beside lactose and this allow to other mutation in other sites in DNA to express, affecting positively or negatively in xanthan production. Therefore, the results reflect the interaction between all Xanthomonas genome including beta-galactosidase and whey media components.
Plasmid isolation and gel electrophoresis of the best selected transconjugants proved that they received the suicidal plasmid pSUP5011 and they lost it after Tn 5 insertion into transconjugants chromosomal DNA which expressed by their resistance to some antibiotics markers. This was clear by the absence of the plasmid band in transconjugants profile and these result confirmed the study done by Simon et al. (1983), Gonzalez et al. (1997) and Gil-serrano et al. (1998).
From the first steps, transconjugant TC 49 was the best xanthan producer when whey was used. The second step of genetic improvement was induction of mutation using Ultraviolet radiation; therefore, decreasing of survival ratio with increasing the exposure time agreed with the Hariharan and Gerutti (1977), Kunz and Glickman (1984) and Rodrigues et al. (2009). They used UV radiation as a tool for induction of mutation depending on that UV light does not cause ionization but it is highly mutagenic agent and its main effect is to cause the dimerization of adjacent pyrimidines. Although the results of the UV mutants still do not exceed the production fold of xanthan gum stated by Kamal et al. (2003) who used a chemical mutagenic agent ethyl methanesulfonate and obtained a mutant strain giving 30% over production of xanthan gum compared by the parent strain. But using UV irradiation as a physical mutagenic agent is still safer than other mutagenic chemicals used for induction of mutation where the chemical hazards has been eliminated specially if xanthan product will be used in many human and animal-related applications.
The results of UV treatment showed that some mutants managed to increase their production of xanthan gum with increasing the time of exposure, where some mutants that exposed to UV for 11 min. produce more xanthan than those which exposed to UV for 7 or 9 min. This could be explained according to Patel et al. (1996) and Rodriguez and Aguilar (1997) who stated that X. campestris and some polysaccharides-secreted microorganisms secreted more polysaccharides to protect its self against radiation. Although there is an overproduction of xanthan gum from the selected mutants and transconjugants on lactose-containing media and whey-containing media but wild type strain and some mutants as 49/11/2 and 49/11/5 still have a high xanthan production on both glucose-containing media and sucrose-containing media these results was agreed and supported by Funahashi et al. (1987) and Souw and Demain (1980) who stated glucose and sucrose were the best carbon sources for xanthan production.
The effect of different carbon sources on xanthan production by the best two mutants Tn-UV-1 and Tn-UV-2 through seven fermentation days. The results showed increasing in xanthan production measured as xanthan weights by time, these results agreed with Peters et al. (1992) who isolated xanthan gum through different fermentation periods ranging from two to eleven days. These two mutants able to utilize lactose or whey with high yield of xanthan gum, one of them Tn-UV-2 produced higher xanthan than wild type in three different carbon sources and higher than mother strain TC 49 in four different carbon sources including glucose and sucrose which considered to be the recommended carbon source proved.
After extraction and purification of xanthan gum from the two best mutants besides wild type strain, characterization of xanthan gum was performed using FTIR in order to ensure the purity of the obtained xanthan gum, The FTIR spectra of synthesized xanthan gum for identification functional groups were obtained (Fig. 6). The x-axis represents wavelength (cm-1) and y-axis shows the light transmittance through the sample. The FTIR spectrum of the sample shows the carboxyl, carbonyl, acetal and these groups are very important. Xanthan gum with high acetal content was easily soluble in water that was due to high hydrophilic properties of the produced xanthan. These results were in agreement with Gilani et al. (2011) who stated that these groups are as important as they were formed during the production of xanthan gum from different carbon sources. The spectra of functional groups are summarized in Table 1, these results indicate the accommodating between xanthan gum produced from the two mutant strains and commercial xanthan gum. The results showed that the presence of absorption peaks at 900-790 cm-1 which confirmed the presence of β-glycoside linkages (Stacy and Barker, 1960) who reported that all polyglucans peak containing β-glycoside. On the other hand, the presence of absorption peaks at 1625-1600 cm-1 indicating the presence of pyruvate function and absorption peaks at 1749-1725 cm-1 may support the presence of acetyl group as confirmed by Su et al. (2003). FTIR of commercial xanthan gum compared with wild type strain, Tn-UV-1 and Tn-UV-2 proved that the mutant’s product has the same chemical composition of commercial xanthan gum. All previous results including physical and chemical characterization of xanthan gum products which produced by the two mutants after extraction and purification showed that the obtained polysaccharide was xanthan gum.
CONCLUSION
Finally, it was clear from the all above result that Tn-UV-2 mutant was the best xanthan producer in all under investigation cultures when lactose or whey was used and after characterization of xanthan as well. This mutant is promising mutant for further studies focusing on increasing its ability to utilize media and xanthan production when its optimized condition of production applied.
REFERENCES
- Al-Turki, A.I., A.A. Khattab and A.M. Ihab, 2008. Improvement of glucoamylase production by Aspergillus awamori using microbial biotechnology techniques. Biotechnology, 7: 456-462.
CrossRefDirect Link - Bertani, G., 1951. Studies on lysogenesis I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol., 62: 293-300.
PubMedDirect Link - Belov, O.V., E.A. Krasavin and A.Y. Parkhomenko, 2009. Model of SOS-induced mutagenesis in bacteria Escherichia coli under ultraviolet irradiation. J. Theor. Biol., 261: 388-395.
CrossRef - Cadmus, M.C., C.A. Knutson, A.A. Lagoda, J.E. Pittsley and K.A. Burton, 1978. Synthetic media for production of quality xanthan gum in 20 liter fermentors. Biotechnol. Bioeng., 20: 1003-1014.
CrossRef - Castilho, B.A., P. Olfson and M.J. Casadaban, 1984. Plasmid insertion mutagenesis and lac gene fusion with Mini-Mu bacteriophage transposons. J. Bacteriol., 158: 488-495.
Direct Link - De Bruijn, F.J., 1987. Transposon Tn5 mutagenesis to map genes. Methods Enzymol., 154: 175-196.
PubMed - De Vuyst, L., A. Vermeire, J. van Loo and E.J. Vandamme, 1987. Nutritional, physiological and process technological improvements of xanthan fermentation process. Mec. Fac. Landbouwwet Rijkuniv Gent, 52: 1881-1900.
Direct Link - Dye, D.W., J.F. Bradbury, M. Goto, A.C. Hayward, R.A. Lelliott and M.N. Schroth, 1980. International standards for naming pathovars of phytopathogenic bacteria and a list of pathovar names and pathotype strains. Rev. Plant Pathol., 59: 153-168.
Direct Link - Ekateriniadou, L.V., S.V. Papoutsopoulou and D.A. Kyriakidis, 1994. High production of xanthan gum by a strain of Xanthomonas campestris conjugated with Lactococcus lactis. Biotechnol. Lett., 16: 517-522.
CrossRef - El-Sawah, M.M.A. and E.H. Ashour, 1999. Xanthan production from whey treated with immobilized lactase. Pak. J. Biol. Sci., 2: 1240-1244.
CrossRefDirect Link - Frank, J.F. and G.A. Somkuti, 1979. General properties of beta-galactosidase of Xanthomonas campestris. Applied Environ. Microbiol., 38: 554-556.
Direct Link - Fu, J.F. and Y.H. Tseng, 1990. Construction of lactose-utilizing Xanthomonas campestris and production of xanthan gum from whey. Applied Environ. Microbiol., 56: 919-923.
PubMedDirect Link - Funahashi, H., T. Yoshida and L.L. Taguchi, 1987. Effect of glucose concentration on xanthan gum production by Xanthomonas campestris. J. Ferment. Technol., 65: 603-606.
CrossRef - Garcia-Ochoa, F., V.E. Santos, J.A. Gasas and E. Goemz, 2000. Xanthan gum: Production, recovery and properties. Biotechnol. Adv., 18: 549-579.
CrossRefDirect Link - Garca-Ochoa, F., V.E. Santos and A.P. Fritsch, 1992. Nutritional study of Xanthomonas campestris in xanthan gum production by factorial design of experiments. Enzyme Microbiol. Technol., 14: 991-996.
CrossRefDirect Link - Giannouli, P. and E.R. Morris, 2003. Cryogelation of xanthan. Food Hydrocolloids, 17: 495-501.
CrossRef - Gilani, S.L., G.D. Najafpour, H.D. Heydarzadeh and H. Zare, 2011. Kinetic models for xanthan gum production using Xanthomonas campestris from molasses. Chem. Ind. Chem. Eng. Q.
Direct Link - Gil-Serrano, A.M., M.A. Rodriguez-Carvajal, P. Tejero-Mateo, J.L. Espartero, J. Thomas-Oates, J.E. Ruiz-Sainz and A.M. Buendia-Claveria, 1998. Structural determination of a 5-O-methyl-deaminated neuraminic acid (Kdn)-containing polysaccharide isolated from Sinorhizobium fredii. Biochem. J., 334: 585-594.
PubMedDirect Link - Gonzalez, C.F., E.A. Pettit, V.A. Valadez and E.M. Provin, 1997. Mobilization, cloning and sequence determination of a plasmid-encoded polygalacturonase from a phytopathogenic Burkholderia (Pseudomonas) cepacia. Mol. Plant Microbe. Interact., 10: 840-851.
PubMedDirect Link - Hamcerencu, M., J. Desbrieres, M. Popa, A. Khoukh and G. Riess, 2007. New unsaturated derivatives of Xanthan gum: Synthesis and characterization. Polymer, 48: 1921-1929.
CrossRefDirect Link - Hariharan, P.V. and P.A. Gerutti, 1977. Formation of products of the 5-6 dihydroxy dihydrotymine type by ultraviolet light in HeLa cells. Biochemistry, 16: 2791-2795.
CrossRef - Jain-Raina, R. and S.B. Babbar, 2011. Evaluation of blends of alternative gelling agents with agar and development of xanthagar, a gelling mix, suitable for plant tissue culture media. Asian J. Biotechnol., 3: 153-164.
CrossRefDirect Link - Kamal, F., H. Mehrgan, M.M. Assadi and S.A. Mortazavi, 2003. Mutagenesis of Xanthomonas campestris and selection of strains with enhanced xanthan production. Iran. Biomed. J., 7: 91-98.
Direct Link - Kar, R., S. Mohapatra, S. Bhanja, D. Das and B. Barik, 2010. Formulation and in vitro characterization of xanthan gum-based sustained release matrix tables of isosorbide-5- mononitrate. Iranian J. Pharm. Res., 9: 13-19.
Direct Link - Katzen, F., A. Becker, A. Zorreguieta, A. Puhler and L. Ielpi, 1996. Promoter analysis of the Xanthomonas campestris pv. campestris gum operon directing the biosynthesis of the xanthan polysaccharide. J. Bacteriol., 178: 4313-4318.
Direct Link - Katzen, F., D.U. Ferreiro, C.G. Oddo, M.V. Ielmini, A. Becker, A. Puhler and L. Ielpi, 1998. Xanthomonas campestris pv. campestris gum mutants: Effects on xanthan biosynthesis and plant virulence. J. Bacteriol., 180: 1607-1617.
PubMedDirect Link - Kunz, B.A. and B.W. Glickman, 1984. The role of pirimidine dymers as premutagenic lesions: A study of targeted vs untargeted mutagenesis in the lac1 gene of Escherichia coli. Genetics, 106: 347-364.
PubMedDirect Link - Moraine, R.A. and P. Rogovin, 1971. Xanthan biopolymer production at increase concentration by pH control. Biotechnol. Bioeng., 13: 381-391.
CrossRef - Nahvi, I. and H. Moeini, 2004. Isolation and identification of yeast strains with high beta-galactosidase activity from dairy products. Biotechnology, 3: 35-40.
CrossRefDirect Link - Nasr, S., M.R. Soudi and M. Haghighi, 2007. Xanthan production by a native strain of X. campestris and evaluation of application in EOR. Pak. J. Biol. Sci., 10: 3010-3013.
CrossRefPubMedDirect Link - Nies, D., M. Mergeay, B. Friedrich and H.G. Schlegel, 1987. Cloning of plasmid genes encoding resistance to cadmium, zinc and cobalt in Alcaligenes eutrophus CH34. J. Bacteriol., 169: 4865-4868.
PubMedDirect Link - Oppezzo, O.J. and R.A. Pizarro, 2003. Inhibition of sulfur incorporation to transfer RNA by ultraviolet-A radiation in Escherichia coli. J. Photochem. Photobiol., 71: 69-75.
CrossRef - Page, W.J. and H.L. Sadoff, 1975. Relationship between calcium and uroinic acids in the encystment of Azotobacter vinelandii. J. Bacteriol., 122: 145-151.
PubMedDirect Link - Patel, K.R., J.A. Wyman, K.A. Patel and B.J. Burden, 1996. A mutant of Bacillus thuringiensis producing a dark-brown pigment with increased UV resistance and insecticidal activity. J. Invert. Pathol., 67: 120-124.
Direct Link - Peters, H.U., H. Herbst, P.G.M. Hesselink, H. Lunsdorf, A. Schumpe and W.D. Deckwer, 1989. The influence of agitation rate on xanthan production by Xanthomonas campestris. Biotechnol. Bioeng., 34: 1393-1397.
CrossRef - Peters, H.U., I.S. Suh, A. Schumpe and W.D. Deckwer, 1992. Modeling of batchwise xanthan production. Can. J. Chem. Eng., 70: 742-750.
CrossRef - Rodriguez, H. and L. Aguilar, 1997. Detection of Xanthomonas campestris mutants with increased xanthan production. J. Ind. Microbiol. Biotechnol., 18: 232-234.
CrossRefDirect Link - Roseiro, J.C., M.E. Esgalhado, M.T.A. Collaco and A.N. Emery, 1992. Medium development for xanthan production. Process Biochem., 27: 167-175.
Direct Link - Rottava, L., G. Batesini, M.F. Silva, L. Lerina and D. De-Oliveirab et al., 2009. Xanthan gum production and rheological behavior using different strains of Xanthomonas sp. Carbohydrate Polymer, 77: 65-71.
CrossRef - Silva, M.F., R.C.G. Fornari, M.A. Mazutti, D. de Oliveira and F.F. Padilha et al., 2009. Production and characterization of xantham gum by Xanthomonas campestris using cheese whey as sole carbon source. J. Food Eng., 90: 119-123.
CrossRefDirect Link - Simon, R., U. Priefer and A. Puhler, 1983. A broad host range mobilization system for in vivo genetic engineering: Transposon mutagenesis in gram negative bacteria. Nat. Biotechnol., 1: 784-791.
CrossRef - Stauffer, K.R. and J.G. Leeder, 1978. Extracellular microbial polysaccharide production by fermentation on whey or hydrolyzed whey. J. Food Sci., 43: 756-758.
CrossRef - Su, L., W.K. Ji, W.Z. Lan and X.Q. Dong, 2003. Chemical modification of xanthan gum to increase dissolution rate. Carbohydrate Polymer, 53: 497-499.
CrossRef - Vojnov, A.A., A. Zorreguieta, J.M. Dow, M.J. Daniels and M.A. Dankert, 1998. Evidence for a role for the gumB and gumC gene products in the formation of xanthan from its pentasaccharide repeating unit by Xanthomonas campestris. Microbiology, 144: 1487-1493.
CrossRef - Rosalam, S. and R. England, 2003. Review of xanthan gum production from unmodified starches by Xanthomonas comprestris sp. Enzyme. Microbiol. Technol., 39: 197-207.
CrossRefDirect Link