
Ayoob Obaid Alfalahi
Department of Field Crops Sciences, College of Agriculture, University of Anbar, Anbar, Iraq
LiveDNA: 964.24069
Ahmed Talib Saleh
Department of Field Crops Sciences, College of Agriculture, University of Anbar, Anbar, Iraq
Asian Journal of Plant Sciences
Year: 2018 | Volume: 17 | Issue: 3 | Page No.: 160-172







ABSTRACT
Background and Objective: Weeds have exceptional capability to reproduce and evolved in response to weeding practices via modified genetic structure. Although, weeds stand for a consider concerns in the agriculture sector, it had received less attention on the molecular level. The current study designed to elucidate the evolution of geographically-isolated johnsongrass species using Random Amplified Polymorphic DNA (RAPD) and Internal Transcribed Spacer (ITS) molecular markers. Materials and Methods: Eight fresh samples of johnsongrass plants collected from different sites around Baghdad capital city. Total genomic DNA was extracted from the eight johnsongrass populations in addition to the most cultivated sorghum cultivars (Enqath and Cafair). The molecular assessment was conducted with aid of ITS and RAPD markers. Sequencing and RAPD data were statistically analyzed with MEGA6 (version 6.0) and MVSP (version 3.22) software, respectively. Results: The RAPD data revealed a polymorphism percentage of 55.8% indicating the biased genetic performance of some johnsongrass (Sorghum halepense L.) populations which demonstrating different evolution path in response to selective pressure. The ITS sequencing showed that the weed population still had a big opportunity to evolve in response to natural environment selection and/or weeding practices. Conclusion: The detected genetic relationship between domestic varieties of sorghum and wild johnsongrass populations showed that weeding practices and domestic-related species may play an important role in updating weeds gene pool.
PDF Abstract XML References Citation
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ayoob Obaid Alfalahi and Ahmed Talib Saleh, 2018. Molecular Detection of Johnsongrass (Sorghum halepense L.) Evolution in Iraq. Asian Journal of Plant Sciences, 17: 160-172.
DOI: 10.3923/ajps.2018.160.172
URL: https://scialert.net/abstract/?doi=ajps.2018.160.172
DOI: 10.3923/ajps.2018.160.172
URL: https://scialert.net/abstract/?doi=ajps.2018.160.172
INTRODUCTION
Genetic diversity is a mile stone in achieving biodiversity within and between different species, which undoubtedly will broaden the genetic base and improve individual and plant species ability to adapt in a wider range of environmental conditions. Weed species is a stumbling block encountered any successful agricultural project and results in significant deterioration in yield quantity and quality.
Despite its extreme importance, weeds have not been largely studied on the molecular level to trace the evolution of its populations and thus hinder their development in a timely manner to minimize their harm to the least limit. One of the most important issues in weed control is its high efficiency in evolving new techniques to survive and establishing in wide range of habits1.
Weed populations like johnsongrass (Sorghum halepense L.) showed rapid and continuous development in herbicides tolerance, especially when adopting the same techniques and herbicides. The problem of developed weed populations get complicated when many relative species belong to the same genus located near each other (Sorghum bicolor L.) within the same habits, which may provides a constant source of new genes that may update the gene pool of weed and in turn gives weed an opportunity to skip from herbicide effect in what well known term "Mimicry". Also, multiple types of herbicides used in controlling weed population create a kind of selective pressure on these populations combined with some genetic mutations that may confer herbicide resistance, therefore numerous weed species have evolved resistance to one or multiple herbicides2,3.
The importance of molecular markers no longer a secret to many, but its applications are renewed. So, it has been developed as a successful mean to estimate the size of genetic divergence between different genotypes and its high accuracy and environmentally independence may be its best features. Random Amplified Polymorphic DNA (RAPD) is one of the simplest molecular markers which provides a primary and fast evaluation to the genetic diversity without the need to previous knowledge about the targeted genome sequence, in addition to its low cost and less labor. Results based on such oligonucleotides would be priceless if it combined with more accurate molecular techniques especially those dealing with single nucleotide polymorphism like sequencing of Internal Transcribed Spacer (ITS) region of the 18S-5.8S-26S nuclear ribosomal cistron4.
The rapid evolution and the highly informative of ITS region compared with other non-coding regions make it one of the most popular sequences for phylogenetic inference at the molecular level in plants5 and it has been widely used for evaluating genetic relationship between different species and even between different populations within the same species6.
The ITS region located between the Small subunit (SSU) and the Large subunit (LSU) ribosomal RNA (rRNA) genes and consisting of two of non-coding spacer regions usually refers to as ITS-A or and ITS-B or 2 interspersed by the 5.8S rRNA gene. Two pairs of universal primers (ITS 1 = Forward and ITS 4 = Reverse) proposed by White et al.7 to amplify ITS region with molecular size ranged between 650-900 bp, including the 5.8S gene.
Far as it is known that there are no serious studies concerned with the evolution of weed populations in the local area and its impact on the emergence of new populations with new genes that may increase their chances in tolerating unfavorable conditions and withstanding herbicides effect. Therefore, this study aimed to use RAPD decamer primers for primary screening and ITS 1 and ITS 4 sequence to assess the genetic diversity within johnsongrass populations collected from different geographical sites around Baghdad capital city and compared their results with those of the most two cultivated varieties of sorghum (Sorghum bicolor L.) in the same area.
MATERIALS AND METHODS
Eight samples of fresh johnsongrass plants collected from different sites in the Baghdad capital city (Fig. 1), (latitude 33°18'0" N, longitude 44°24'0" E) in Iraq. Taken into consideration the different regions of collected samples between marginal lands, cultivated fields and canals in an attempt to bring weed samples going through different selective pressure. Also, two of the most cultivated varieties of Sorghum involved (Enqath and Cafair).
DNA extraction and RAPD amplification: Samples of johnsongrass plant collected and saved on ice in plastic bags until reaching the lab. Fresh leaves from each sample sterilized and washed with distilled water, then it was ground in liquid nitrogen and the Genomic DNA Mini Kit (Plant) (Geneaid Biotech Ltd.) was used to extract and purified the total DNA as mentioned in manufacturer's guide. DNA quality and quantity checked with the spectrophotometer using the following formulas8:
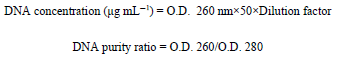 |
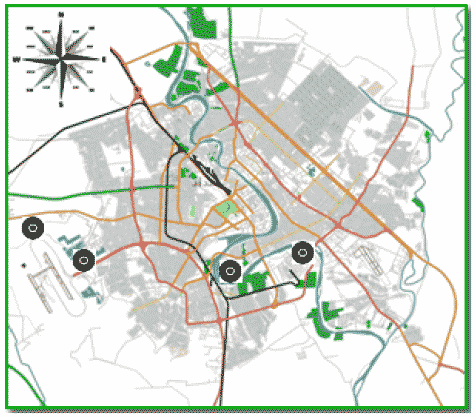 | |
| Fig. 1: | Map showed the collection sites |
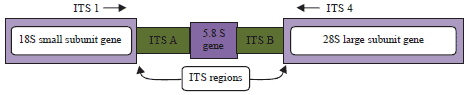 | |
| Fig. 2: | ITS region between the small subunit and the large subunit ribosomal RNA (rRNA) genes interspersed by the 5.8S rRNA gene |
Samples diluted to the final concentration of 50 ng μL–1 for amplification. Five of RAPD decamer primers (OPA-09 ('5-GGGTAACGCC-3'); OPB-03 ('5-GTGCGAGAAC-3'); OPH-13 ('5-GACGCCACAC-3'); OPH-18 ('5-GAATCGGCCA-3'); RI-16 ('5-GTCGCCGTCA-3')) supplied by Alpha DNA-Canada in lyophilized form diluted with deionized distilled water to 10 pmol/μL for DNA amplification with PCR.
PCR reactions were 25 μL in volume containing 12.5 μL of Green Master Mix (2X PCR buffer; pH = 8.5), dNTPs (400 μM) and Taq polymerase (5 U), 50 ng total genomic DNA, 1.5 μL of each 10 pmol μL–1 RAPD primer. Then reaction volume was completed to 25 μL with nuclease-free water. Thermal profile for PCR reaction was as follows: Initial denaturation was on 95ο for 4 min, denaturation step was on 94ο for 40 sec, annealing was on 44ο for 1.30 min, extension was done on 72ο for 2.30 min, final extension was on 72ο for 10 min.
Ten of each amplified product was analyzed on 2% agarose gel along with 1 kb DNA ladder. The gel is left to run for 90 min with a 100 volt/50 mAmp current. Then UV transilluminator was used to visualize the gel and the image was documented with gel documentation system.
Amplification and sequencing of ITS region: Pair of specific primers used to amplify the ITS region (Fig. 2), ITS 1 (F-5'-TCCGTAGGTGAACCTGCGG-'3) as forward and ITS 4 (ITS 4 R- 5'-TCCTCCGCTTATTGATATGC-'3) as reverse primer7.
The PCR reaction was carried out in a 25 μL reaction containing 12.5 μL of Green Master Mix, 1 μL of 10 pmol μL–1 of primer, 3 μL of DNA template and the volume was completed to 25 μL using nuclease-free water. Thermocycling conditions as follows:
| • | Initial denaturation was on 95ο for 4 min |
| • | Denaturation step was on 95ο for 30 sec |
| • | Annealing was on 55ο for 30 sec |
| • | Extension was done on 72ο for 1 min |
| • | Final extension was on 72ο for 7 min |
The PCR products resolved on 1% agarose gel. The gel was prepared by dissolving 1 g of agarose in 100 mL of 1×TAE buffer using a microwave oven. The mixture was left to cool to about 55-60 C before a 1 μL of 10 mg mL–1 of ethidium bromide was added. It was then poured into the electrophoresis tray, secure the combs in place and left to cool and solidify for about 30 min. After the gel was set, the combs removed carefully and the tank was placed in the electrophoresis system containing running buffer consisting of 1×TAE. The buffer was poured until it covered the gel for about 2 mm. About 5 μL of each PCR product along with the negative control and a 100 bp DNA ladder loaded into the wells, the system cover was then put into place and the system was turned on. The gel was left to run for 90 min with a 100 volt/50 mAmp current. Following electrophoresis, visualization was conducted with a UV transilluminator and the image was captured by digital camera (Canon, US). This camera has the appropriate filter and a suitable program for illumination of EtBr-stained gels.
Sequencing step for ITS region with the use of ITS 1 as forward and ITS 4 as reverse was carried out by Macrogen Inc. (Seoul, Korea) using their ABI 3730 Genetic Analyzer (Applied Biosystems, US).
Statistical analysis of molecular results: Every scorable band was considered as single allele/locus and was scored as 1 and 0 for the present and absent band, respectively. Bivariate data (1-0) adopted to calculate genetic similarity following Unweighted Pair Group of Arithmetic Means (UPGMA) proposed by Nei and Li 9 as follows:
Genetic similarity (GS) = 2Nij/(Ni+Nj) |
where, Gs is genetic similarity between two populations, Nij is total number of loci (bands) in populations i and j, Ni is total number of loci (bands) in population I and Nj is total number of loci (bands) in population j.
Polymorphism was estimated according to the following formula:
Polymorphism (%) = (Np/Nt)×100 |
where, Np is the number of polymorphic bands of primer and Nt is the total number of bands of same primer.
Evolutionary divergence between different populations was estimated according to the number of base substitutions per site from between different sequences. Analyses conducted by using neighbor-joining method10. Statistical analyses performed with aid of MEGA6 (Molecular Evolutionary Genetics Analysis version 6.0.) computer software11. Codon positions containing gaps and missing data were eliminated.
RESULTS AND DISCUSSION
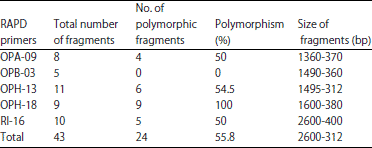
The five used RAPD primers amplified a total number of 43 fragment (Table 1), 24 of these were polymorphic scoring a polymorphism percentage of 55.8%. The amplified fragments ranged in size between 2600-312 bp. Similar findings documented by Abdel-Fatah et al.12.
Primer OPA-09 (Fig. 3) was succeeded in amplifying 8 bands, half of it were polymorphic scoring 50% polymorphism percentage. Fragments size ranged from 1360-370 bp. The used primer was able to distinguish between johnsongrass and Sorghum cv. by losing two annealing sites in Sorghum varieties with a molecular size of 1180 and 370 bp. The primer approved its efficiency by distinguishing between the two used Sorghum cultivars, Enqath and Cafair as amplifying unique fragment with a molecular size of 630 bp just in Cafair (population No. 8), on the other hand, the primer losing annealing site results in a fragment with 583 bp of size. These results indicated the magnitude of human breeding methods in modifying the plant genome even within the same species compared with nature effect on the same way.
Some primers like OPB-03 (Fig. 4) had so common sequence which resulting in trivial performance. Therefore, the primer failed in discriminate any of the used genotypes. Although, the primer generated 5 bands ranged in their molecular size between 1490-360 bp.
| Table 1: | Number and molecular size of scorable bands generated by five RAPD primers in eight of johnsongrass populations and two Sorghum cultivars |
 | |
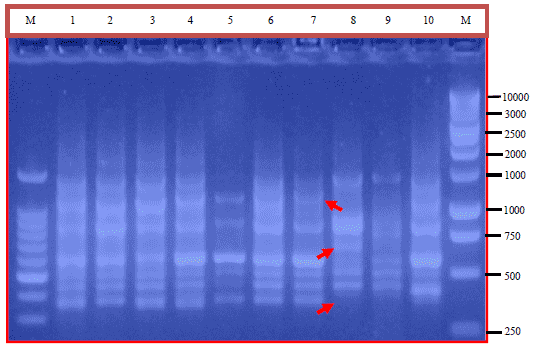 | |
| Fig. 3: | PCR amplified products of 8 johnsongrass populations (1, 2, 3, 4, 5, 6, 7 and 10) and two Sorghum cultivars Enqath (8) and Cafair (9) using RAPD primer OPA-09 running on agarose gel (2%). M = Molecular marker (100 bp and 1 kb ladder) |
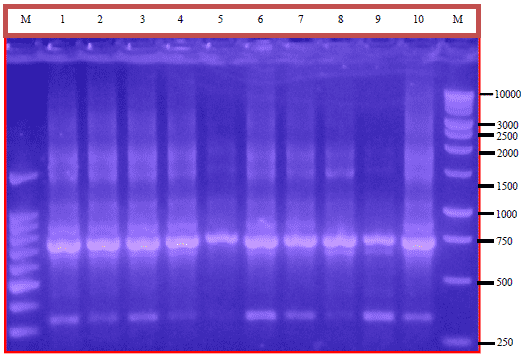 | |
Fig. 4: | PCR amplified products of 8 johnsongrass populations (1, 2, 3, 4, 5, 6, 7 and 10) and two Sorghum cultivars Enqath (8) and Cafair (9) using RAPD primer OPB-03 running on agarose gel (2%). M = Molecular marker (100 bp and 1 kb ladder) |
Eleven bands amplified with aid of OPH-13 primer (Fig. 5), six of these found to be polymorphic scoring 54.5% polymorphism percentage. The generated bands ranged in its molecular size between 1495-312 bp. The primer has identical annealing site in all collected johnsongrass populations, showing no differences among them. In the same time, it was efficient in characterizing genetic variation between johnsongrass and sorghum species on one hand and on the other hand between the two Sorghum cultivars.
 | |
| Fig. 5: | PCR amplified products of 8 johnsongrass populations (1, 2, 3, 4, 5, 6, 7 and 10) and two Sorghum cultivars Enqath (8) and Cafair (9) using RAPD primer OPH-13 running on agarose gel (2%). M = Molecular marker (100 bp and 1 kb ladder) |
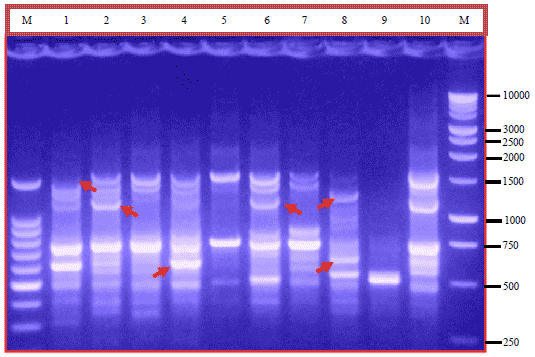 | |
| Fig. 6: | PCR amplified products of 8 johnsongrass populations (1, 2, 3, 4, 5, 6, 7 and 10) and two Sorghum cultivars Enqath (8) and Cafair (9) using RAPD primer OPH-18 running on agarose gel (2%). M = Molecular marker (100 bp and 1 kb ladder) |
Both Sorghum cv. lost two fragments compared with johnsongrass populations with a molecular size of 1250 and 570 bp. While Cafair cv. (population 9) gain two extra fragments its molecular size reached 410 and 250 bp.
OPH-18 primer (Fig. 6) was completely polymorphic as it produced a total of 9 bands, all were polymorphic which indicating its highly efficiency in detecting genetic variation between both species and within the same species. The amplified products differed in its molecular size between 1600 and 380 bp. Genetically, the first population acted differentially, when it lost the fragment of the highest molecular size (1600 bp) against the rest of johnsongrass populations. Population 3, 5 and 7 lost fragment size of 1200 bp.
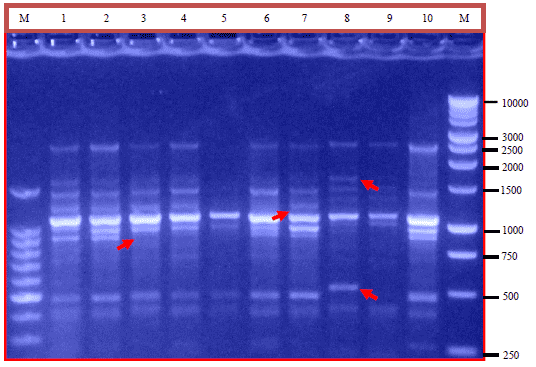 | |
| Fig. 7: | PCR amplified products of 8 johnsongrass populations (1, 2, 3, 4, 5, 6, 7 and 10) and two Sorghum cultivars Enqath (8) and Cafair (9) using RAPD primer RI-16 running on agarose gel (2%). M = Molecular marker (100 bp and 1 kb ladder) |
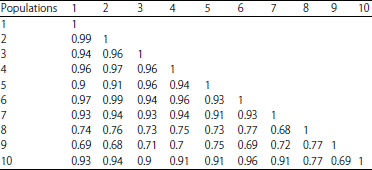
| Table 2: | Estimated values of Genetic Similarity (GS) based on generated fragments of five RAPD primers of johnsongrass populations (1, 2, 3, 4, 5, 6, 7 and 10) and Sorghum cultivars Enqath (8) and Cafair (9) |
 | |
The absence of 625 bp molecular size fragment in population 3 and 5 make it fingerprint for these populations. The Sorghum cultivars were differed against each other with the corresponding primer.
Polymorphic percentage resulting from the using of RI-16 primer (Fig. 7) was 50%, this was based on the five polymorphic fragments out of the 10 total bands generated by the previously mentioned primer. Population 3 and 4 lost annealing site to the used primer resulting in producing a fragment of 980 bp in other populations. Enqath cv. gained one unique fragment (560 bp) which seems it fingerprint against the rest of the studied populations.
Phylogenetic analysis based on RAPD data: Data matrix on the basis of the presence (1) or absence (0) of each DNA fragment for each RAPD primer was used to calculate the genetic similarity (GS) (Table 2) and construct the phylogenetic tree (Fig. 7) following unweighted pair group method with arithmetic average (UPGMA). Results from genetic similarity matrices created by using Nei and Li9 coefficients revealed that as expected, high genetic similarity among different populations within the same species13-15. It's so clear that the lowest GS values revealed by all johnsongrass populations against the two cultivars of Sorghum and it ranged between 0.68-0.77.
The genetic similarity between the two Sorghum cultivars was low enough (0.77) to draw attention especially as we compared these values with those among the natural population of johnsongrass, which scored higher values reached 0.99. This may a clear reference to the magnitude of breeding methods adopted by the human in shifting the gene pool of economic plant populations. The impressive thing that the same value of GS (0.77) was showed by Enqath sorghum cv. against one of johnsongrass populations (population 6) which may indicate the common ancestor or the rarely cross pollination between these populations13.
The GS values among different johnsongrass populations were no less than 0.90, while its highest value reached 0.99. Based on these values constructed the phylogenetic tree (Fig. 8) was consisting of two major clusters, each for one species matching what is expected, one consisting of all the natural populations of johnsongrass, whereas the second cluster contains the two Sorghum cultivars (Enqath and Cafair).
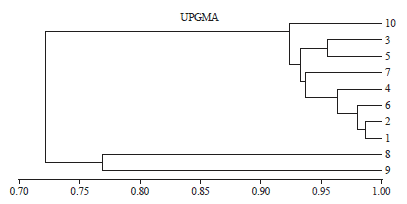 | |
| Fig. 8: | Constructed phylogenetic tree based on generated fragments of five RAPD primers of johnsongrass populations (1, 2, 3, 4, 5, 6, 7 and 10) and Sorghum cultivars Enqath (8) and Cafair (9) |
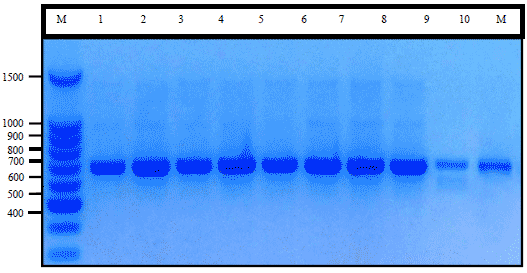 | |
| Fig. 9: | PCR amplified products of ITS region of eight johnsongrass populations (1 to 8) and two sorghum cv. 9 (Enqath) and 10 (Cafair) using ITS 1-F and ITS 4-R specific primers, running on agarose gel (2%). M = Molecular marker (100 bp) |
The first major cluster showed slight distinctness of the tenth population to be in sub-cluster alone. These minor differences encourage to go with ITS sequencing which may put us in better view of the genetic evolution of johnsongrass populations correlated with the most cultivated Sorghum cultivars.
ITS amplification and sequencing: The efficient amplifying to the ITS region with aid of pair of resulting in DNA fragments with molecular size of ~700 bp (Fig. 9).
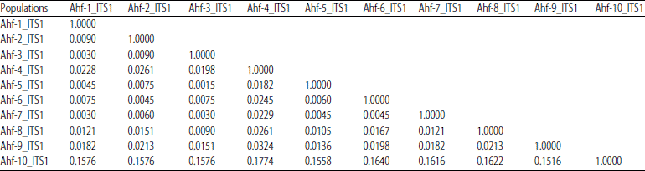
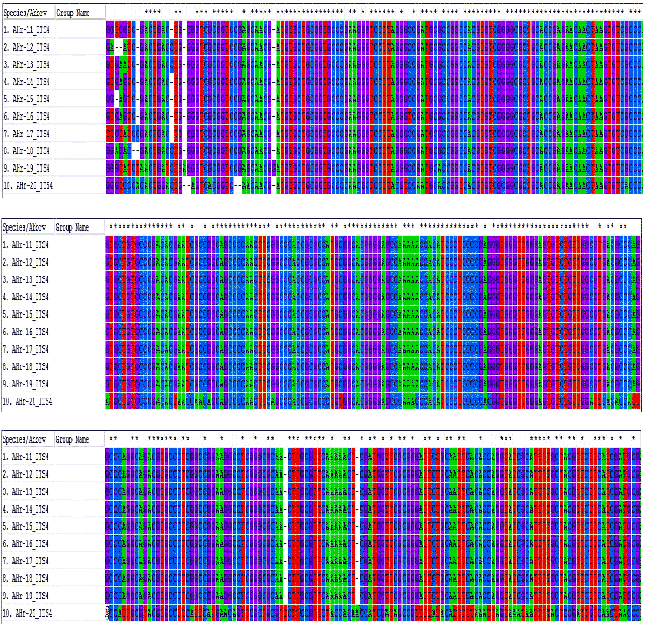
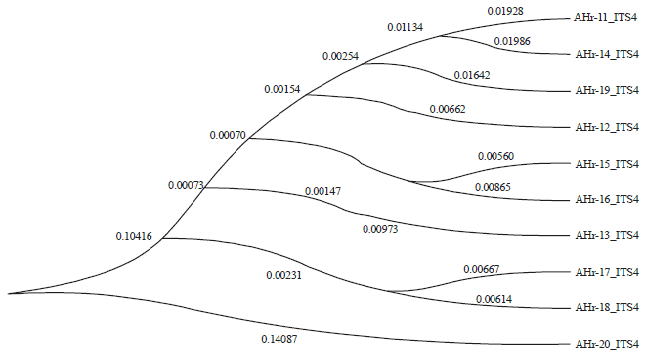
The established alignments of ITS-1 sequences (Fig. 10) showed that there is a clear contrast and expected to some extent between different Sorghum populations belongs to different species and this fairly satisfactory agree with the results of RADP analysis which separated both Sorghum cultivars in single sub-branch14. The distinctness of Sorghum Cafair cv. was easy to detect from its ITS sequence results combined with genetic distance based on ITS-1 single nucleotide polymorphism (Table 3) and it records the highest values (0.1774). These results strengthened by the evolutionary tree (Fig. 11) which separate the previously mentioned cv. in from all the other johnsongrass populations even its partner Enqath cv. Although the common ancestor that they have, they acted differently with the genetic distance of 0.1516 which considered high value compared with the lowest genetic distance (0.0015) showed by population No. 5 of johnsongrass against population No. 3, which allowed the latest to occupy the same sub cluster for their identical performance. The slight difference with ITS-1 sequence resulting in genetic distance among johnsongrass populations ranged between 0.0015-0.0229.
The divergent behavior showed by Sorghum cv. against each other, meanwhile one of them tend to be closely related to johnsongrass populations gave a signal to the magnitude of human breeding methods in amendment evolution path of different biota15.
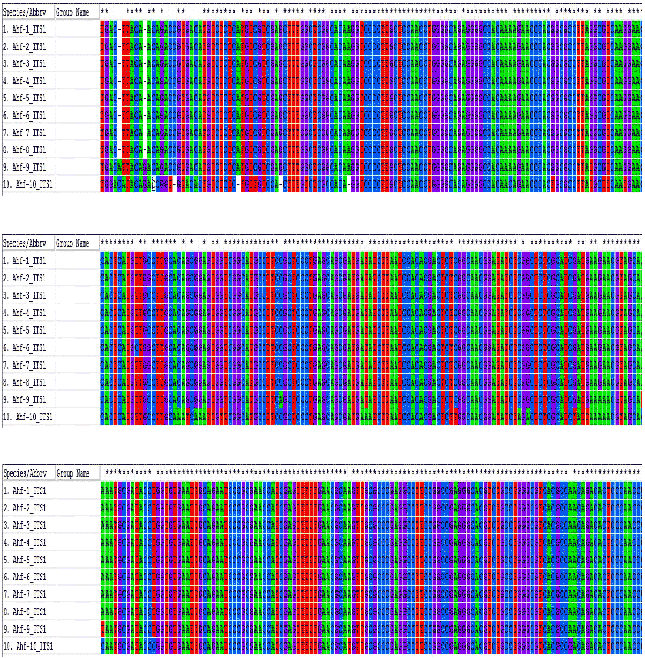 | |
| Fig. 10: | Internal Transcribed Spacer (ITS) sequencing using ITS 1 as forward amplifier for eight samples of Johnsongrass (Sorghum halepense L.) (1 to 8) and two of Sorghum cv. (Sorghum bicolor L.) 9 (Enqath) and 10 (Cafair) |
 | |
| Fig. 11: | Evolutionary divergence tree based on internal transcribed spacer (ITS) sequencing using ITS 1 as forward amplifier for eight populations of Johnsongrass (Sorghum halepense L.) (1 to 8) and two of Sorghum cv. (Sorghum bicolor L.) 9 (Enqath) and 10 |
| Table 3: | Estimates of evolutionary divergence based on Internal Transcribed Spacer (ITS) sequencing using ITS 1 as forward amplifier for eight populations of Johnsongrass (Sorghum halepense L.) (1 to 8) and two of Sorghum cv. (Sorghum bicolor L.) 9 (Enqath) and 10 (Cafair) |
 | |
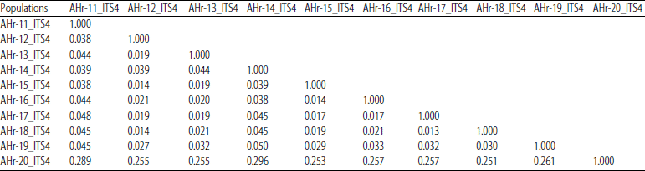
| Table 4: | Estimates of evolutionary divergence based on Internal Transcribed Spacer (ITS) sequencing using ITS 4 as reverse amplifier for eight populations of Johnsongrass (Sorghum halepense L.), (11, 12,13,14,15,16,17 and 18) two of Sorghum cv. (Sorghum bicolor L.) 19 (Enqath) and 20 (Cafair) |
 | |
On the other hand, the minor modification in DNA sequences detected between johnsongrass populations back in part of it to the nearly identical environment conditions involving the collecting sites, but in the same time, this tiny genetic variation within the restricted area indicated a big chance to significant modifications in johnsongrass population genomes in wider geographical areas16.
The ITS 4 results approved the unique genetic composition of Sorghum Cafair cv., whereas the other cv. (Enqath) acted differently as it tends to characterize his performance far away from his partner scoring one of the highest genetic distance mean (Table 4) reached 0.261 against each other. The evolution tree (Fig. 13) put cv. in a single cluster at the same time Enqath cv. showed its contradictory attitude being relatively close to the other species populations of sorghum, which may indicated the common ancestor17,18.
Generally, the genetic distances based on ITS 4 alignments (Fig. 12) among johnsongrass populations ranged from its lowest value 0.013 between population No. 7 and population No. 8 and its highest value 0.048 between population No. 7 and No. 1.
 | |
| Fig. 12: | Internal Transcribed Spacer (ITS) sequencing using ITS 4 as reverse amplifier for eight samples of Johnsongrass (Sorghum halepense L.) (11, 12, 13, 14, 15, 16, 17 and 18) and two of Sorghum cv. (Sorghum bicolor L.) 19 (Enqath) and 20 (Cafair) |
 | |
| Fig. 13: | Evolutionary divergence tree based on Internal transcribed spacer (ITS) sequencing using ITS 4 as reverse amplifier for eight populations of Johnsongrass (Sorghum halepense L.) (11, 12, 13, 14, 15, 16, 17 and 18) two of Sorghum cv. (Sorghum bicolor L.) 19 (Enqath) and 20 (Cafair) |
CONCLUSION
The evolution process in a specific weed populations is highly related with weeding practices. On the other hand, the cultivated species can play a critical role in emerging of new weeds populations with improved traits via horizontal gene transfer.
SIGNIFICANCE STATEMENTS
The current molecular study discovers the evolution of johnsongrass populations (Sorghum halepense L.) and the possible role of cultivated species (Sorghum bicolor) in this process, which may help the researcher to uncover the critical areas of genome contamination that many researchers were not able to explore. Outcomes approved the common genetic background of johnsongrass and sorghum populations, hence all the necessary precautions should be taken in case of genetically-related weeds prevalence next to the cultivated crops, especially the genetically modified to prevent genetic leakage.
REFERENCES
- Murray, B.R. and M.L. Phillips, 2010. Investment in seed dispersal structures is linked to invasiveness in exotic plant species of South-Eastern Australia. Biol. Invasions, 12: 2265-2275.
CrossRefDirect Link - Werle, R., A.J. Jhala, M.K. Yerka, J.A. Dille and J.L. Lindquist, 2016. Distribution of herbicide-resistant shattercane and johnsongrass populations in sorghum production areas of Nebraska and Northern Kansas. Agron. J., 108: 321-328.
CrossRefDirect Link - Alvarez, I. and J.F. Wendel, 2003. Ribosomal its sequences and plant phylogenetic inference. Mol. Phylogenet. Evol., 29: 417-434.
CrossRefPubMedDirect Link - Sun, Y.L., W.G. Park, H.K. Oh and S.K. Hong, 2013. Plant-specific primers for the amplification of the nrDNA ITS region in fungus-associated Pulsatilla species. J. Med. Plants Res., 7: 1969-1978.
CrossRefDirect Link - Wu, B., A. Yang, J. Yan, W. Xu and T. Yu et al., 2014. Comparison of the Internal Transcribed Spacer (ITS) regions between hybrid and their parents in Scallop. Int. J. Biol., 6: 103-109.
CrossRefDirect Link - White, T.J., T.D. Bruns, S.B. Lee and J.W. Taylor, 1990. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In: PCR Protocols: A Guide to Methods and Applications, Innis, M.A., D.H. Gelfand, J.J. Sninsky and T.J. White (Eds.), Academic Press, San Diego, CA, USA, ISBN-13: 9780123721808, pp: 315-322.
CrossRefDirect Link - Sambrook, J. and D.W. Russell, 2001. Molecular Cloning: A Laboratory Manual. 3rd Edn., Cold Spring Harbor Laboratory Press, New York, USA., ISBN-13: 9780879695774, Pages: 2344.
Direct Link - Nei, M. and W.H. Li, 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA., 76: 5269-5273.
CrossRefPubMedDirect Link - Tamura, K., M. Nei and S. Kumar, 2004. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. U.S.A., 101: 11030-11035.
CrossRefDirect Link - Tamura, K., G. Stecher, D. Peterson, A. Filipski and S. Kumar, 2013. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol., 30: 2725-2729.
CrossRefPubMedDirect Link - Abdel-Fatah, B.E., E.A. Ali, A.A. Tag El-Din and E.M. Hessein, 2013. Genetic Diversity among Egyptian Sorghum (Sorghum bicolor L. Moench) Landraces in Agro-Morphological Traits and Molecular Markers. Asian J. Crop Sci., 5: 106-124.
CrossRefDirect Link - Mader, G., L. Castro, S.L. Bonatto and L.B. de Freitas, 2016. Multiple introductions and gene flow in subtropical South American populations of the fireweed, Senecio madagascariensis (Asteraceae). Genet. Mol. Biol., 39: 135-144.
CrossRefPubMedDirect Link - Guo, Q.X., K.H. Huang, Y.U. Yun, Z. Huang and Z.Q. Wu, 2006. Phylogenetic relationships of Sorghum and related species inferred from sequence analysis of the nrDNA ITS region. Agric. Sci. China, 5: 250-256.
CrossRefDirect Link - Kryvokhyzha, D., K. Holm, J. Chen, A. Cornille and S. Glemin et al., 2016. The influence of population structure on gene expression and flowering time variation in the ubiquitous weed Capsella bursa-pastoris (Brassicaceae). Mol. Ecol., 25: 1106-1121.
CrossRefPubMedDirect Link - Vila-Aiub, M.M., M.C. Balbi, P.E. Gundel, C.M. Ghersa and S.B. Powles, 2007. Evolution of glyphosate-resistant johnsongrass (Sorghum halepense) in glyphosate-resistant soybean. Weed Sci., 55: 566-571.
CrossRefDirect Link - Sun, Y., D.Z. Skinner, G.H. Liang and S.H. Hulbert, 1994. Phylogenetic analysis of Sorghum and related taxa using internal transcribed spacers of nuclear ribosomal DNA. Theor. Applied Genet., 89: 26-32.
CrossRefDirect Link - Ng'uni, D., M. Geleta, M. Fatih and T. Bryngelsson, 2010. Phylogenetic analysis of the genus Sorghum based on combined sequence data from cpDNA regions and ITS generate well-supported trees with two major lineages. Ann. Bot., 105: 471-480.
CrossRefDirect Link