
I. Romani
Department of Biological Sciences, Universidade Paranaense, Rua Parigot de Souza 3636, Toledo, Parana, Brazil
A.R. Schuelter
Department of Pos-Graduation in Biotecnology Applied to Agriculture, Universidade Paranaense, Praca Mascarenhas de Moraes s/n, Umuarama, Parana, Brazil
F. Mora
Department of Agronomy, Universidade Estadual de Maringa, Avenida Colombo 5790, Maringa, Parana, Brazil
C.A. Scapim
Department of Agronomy, Universidade Estadual de Maringa, Avenida Colombo 5790, Maringa, Parana, Brazil
E.C.G. Vendruscolo
Department of Genetics, Universidade Federal do Parana, Rua Pioneiro 2153, Jardim Dallas, Palotina, Parana, Brazil
Asian Journal of Plant Sciences
Year: 2009 | Volume: 8 | Issue: 3 | Page No.: 199-205







ABSTRACT
Flower buds from peach tomato plants (Solanum sessiliflorum) were collected to the establishment of a protocol of calli induction from anthers culture. Aseptic cultures were achieved after the immersion of explants into 70% ethanol for 45 sec followed by 0.8% sodium hypochlorite for 5 min. Cultivated anthers developed 50 and 70% of calli with 0.1 mg L-1 indole-3-acetic acid (IAA)+5 mg L-1 kinetin (KIN) and 0.1 mg L-1 indole-3-acetic acid (IAA)+10 mg L-1 kinetin (KIN), respectively. The calli oxidation increased in treatments free of activated charcoal, daily pre-refrigeration (4°C for 48 h) and 20 mg L-1 of kinetin (KIN). The best plant growth regulator concentration for stimulating calli formation was 5 mg L-1 of kinetin (KIN).
PDF Abstract XML References Citation
How to cite this article
I. Romani, A.R. Schuelter, F. Mora, C.A. Scapim and E.C.G. Vendruscolo, 2009. Calli Induction through Anther Culture in Peach-Tomato Plants (Solanum sessiliflorum Dunal). Asian Journal of Plant Sciences, 8: 199-205.
DOI: 10.3923/ajps.2009.199.205
URL: https://scialert.net/abstract/?doi=ajps.2009.199.205
DOI: 10.3923/ajps.2009.199.205
URL: https://scialert.net/abstract/?doi=ajps.2009.199.205
INTRODUCTION
Peach-tomato (Solanum sessiliflorum) is also known as cocona, cubiu, orinoco apple or topiro and it has been cultivated under primitive agricultural methods by indians of eastern Amazonas state in Brazil. Actually, this plant is expanding outside its origin center as an excellent alternative for agroindustry due to some medical properties (Silva Filho et al., 1989, 1999, 2003; Pizzinato et al., 2008). The fruit of peach tomato accumulates high concentration of dietary iron, niacin (vitamin B5), citric acid and pectin. Native people from Amazon drink the fruit juice to control anemia, diabetes, uric acid and cholesterol. Macerated leaves were used in scabies, lice and other itching symptoms (Silva Filho et al., 2003).
The morphologic description of peach-tomato plants was conducted by Silva Filho et al. (2003). Doubts about the reproductive system still remains on this species: autogamy (Pahlen, 1977; Pizzinato et al., 2008) or alogamy (Storti, 1988; Pizzinato et al., 2008). Beyond the fact that peach-tomato seed germination is heterogeneous and decreases quickly by the time of storage (Silva Filho et al., 1999).
The reduction of generations of self-fertilization by just one androgenic step is the main advantage of anther culture in obtaining homozygous lines (Pasqual et al., 2002; Kaltchuk-Santos 2003; Palu et al., 2004; Polci et al., 2004). Anther culture is based on the gametophyte capability to generate haploid plants restored in double-haploids (Kaltchuk-Santos and Bodanese-Zanettini, 2002).
Some additional understanding of relative factors has also been useful for a successful anther culture (Shtereva et al., 1998). Androgenic capability of anther cells is also affected by donor plants physiological conditions, growing medium, plant genotype and flower bud stage (Shtereva et al., 1998; Zargorska et al., 1998; Brasileiro et al., 1999; Chani et al., 2000; Ozkum-Ciner and Tipirdamaz, 2002; Pacheco-Sanchez et al., 2003; Gambino et al., 2007; Fu et al., 2008; Perera et al., 2008).
Temperature as a pre-treatment (pre-refrigeration or refrigeration) is another important factor that has been related with the improvement of androgeny in Solanceae plants (Roca et al., 1991; Peters et al., 1999). Also, one efficient protocol must have calli oxidation control. Some researchers have been pointed out that the explants desinfested in ethanol and chloride solutions; followed by ascorbic acid treatments (Utino et al., 2001) or by amending activated charcoal into growing media (Ribeiro et al., 2000) or both treatments were sufficient to avoid calli oxidation.
The objective of this study was the establishment of a protocol of plant regeneration through calli formation in peach-tomato plants. This study represents the first research on in vitro response in this species, since no other report of anther culture has been described in the literature.
MATERIALS AND METHODS
This research was conducted at the Laboratory of Biotechnology of Unipar (Universidade Paranaense), Toledo ( Pr) from February/2006 to December/2008. Seeds were sowed in plastic trays for seedlings and plantlets were changed to 1L plastic bags (40 days old) and after to 10L plastic pots (90 days old). The growing media were a mixture 3:1 of soil and sand. Diseases, insect control and mineral fertilization followed the usual recommendation of Silva Filho (1999), while water was normally supplied to the plants.
Culture media preparation, in vitro culture conditions and induction to androgenesis: Flower buds with 0.6-0.7 cm (Romani et al., 2006) were collected, rinsed three times in distilled water, rinsed once in distilled water plus Tween 20 and finally rinsed again in distilled water. They were desinfected under laminar flow cabinet (Table 1). Following sterilization, flower buds were rinsed again in distilled water supplied with 600 mg L-1 of ascorbic acid filter-sterilized.
Anthers obtained from excision of aseptic buds were inoculated into test tubes containing 10 mL of MS growing medium (Murashige and Skoog, 1962). Following, the test tubes were sealed with paraffin for further development at 25±1°C and at a photoperiod cycle of 16 h light and 8 h dark. The aseptic treatments were replicated 10 times and each experimental unit had four anthers.Twenty-one days later, the efficiency of the treatments was evaluated by visual observation.
| Table 1: | Evaluation of different aseptic treatments used on anther culture |
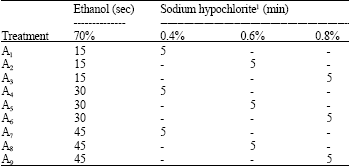 | |
| 1Two drops of Tween 20 | |
The anthers were displayed on MS growing media (pH 5.8) containing growth regulators, forming the following treatments combinations: IAA = 0, 0.01, 0.02, 0.04, 0.08 and 0.1 mg L-1 and Kinetin = 0, 5, 10 and 20 mg L-1 (Table 2). The test tubes were previously maintained under dark room conditions for 7 days for further development at 25±1°C and 16/8 h (light/dark photoperiod conditions, respectively).
The experimental design was a randomized complete block with 24 treatments under the factorial arrangement of six doses of IAA versus four doses of KIN in which the experimental unit was the test tube inoculated with two anthers per treatment and each treatment had five replications. Androgenesis was evaluated thirty-five days later.
Pre-treatments and oxidation control: Flower buds collected as described before were pre-treated under refrigeration. A total of 160 anthers were inoculated into test tubes containing 3 mL of the MS growing medium, plant growth regulator and activated charcoal (Table 3).
| Table 2: | Different indolacetic-3-acid (IAA) and kinetin (KIN) concentrations on peach-tomato anther culture |
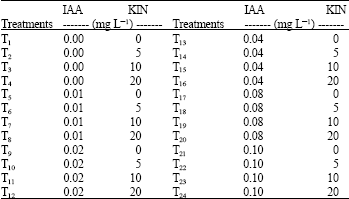 | |
| IAA: Indole-3-acetic acid; KIN: Kinetin | |
| Table 3: | Evaluation of different treatments in order to avoid oxidation on peach-tomato in vitro culture |
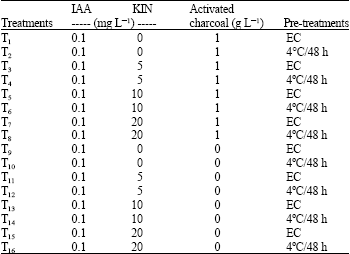 | |
| IAA: Indole-3-acetic acid; KIN: Kinetin; EC: Environmental conditions (25°C and 16/8 h-light/dark photoperiod) | |
The temperature treatments consisted of submitting anthers to 25°C (Environmental condition) and 4°C (Pre-refrigeration condition) for 48 h.
To check oxidation control, the treatments were evaluated according to the presence and absence of activated charcoal (0 and 1 mg L-1) (Table 3). The tubes were stored for 7 days under dark conditions before growth at 25±1°C and 16/8 h (light/dark conditions, respectively).
The experimental design was a randomized complete block with 16 treatments and five replications. Each experimental unit consisted of one test tube inoculated with two anthers. The level of oxidation was evaluated visually, thirty-five days later using the following numerical scale: 0 = non-oxidation, 1 = low oxidation level, 2 = middle oxidation level and 3 = complete oxidation. The calli were evaluated after 50 days. Calli were classified by size according to the following scale: 0 = no calli formation, 1 = small callus, 2 = callus similar to the anthers, 3 = callus larger than anthers.
Effects of plant growth regulators, activated charcoal and refrigeration on explants oxidation and calli induction were investigated by deviance analysis using GLM procedures (Nelder and Wendderburn, 1972). The multinomial distribution with the cumulative logit from the exponential family was assumed as the link function (Hern and Dorn, 2001). This statistic strategy was used due the fact that the parameters did not present a normal data distribution.
The data obtained below were analysed using the PROC GENMOD statistical program and the subroutine TYPE1 was run to apply the likelihood ratio test. The option CONTRAST was used for detecting differences among levels within the sources of variation by orthogonal contrasts (Myers et al., 2002).
RESULTS AND DISCUSSION
Aseptic treatments: The aseptic treatments A8 and A9 (Table 1), using sodium hypochlorite (0.6 e 0.8%) and ethanol for 45 sec, respectively, were the most efficient since 90% of explants were free of contamination. In contrast, the treatments using sodium hypochlorite (0.4 and 0.6%) and ethanol for 15 sec, respectively, showed the higher percentage of contamination (50%). Treatment A5 had 40% of contamination while treatments A3, A4, A6 and A7 had the average of 30% of contaminated tubes. Treatments where the explants were immersed into 70% ethanol for 45 sec seemed to cause no injuries to explants (Table 1). Pasqual et al. (2002) related that 2% sodium hypochlorite for 15 min was the best treatment in coffee explants although their contamination level was 50%. Gambino et al. (2007) also described an efficient aseptic protocol for flower, anther and ovaries in grapevine (Vitis spp.) using 1.5% sodium hypochlorite and 2 drops of Tween 20 for 10 min. Grattapaglia and Machado (1999) commented that an efficient protocol should have the premise in not causing explant damage during the control of microorganisms.
Plant growth regulators effects on androgenesis: Morphogenetic responses were found in 10 out of 24 treatments containing plant growth regulators in which the percentage of induced calli varied from 10 to 70%. The treatments, T22 (0.1 mg L-1 IAA+5 mg L-1 KIN) and T23 (0.1 mg L-1 IAA+10 mg L-1 KIN) promoted 50 and 70% of calli, respectively (Table 4). However, no callogenic responses were found in the other treatments. Pacheco-Sanchez et al. (2003) studied the factors affecting androgenesis in Solanum iopetalum and commented that 27.5% of calli were obtained due to a treatment of anthers with 1 mg L-1 BA+1 mg L-1 IAA.
Furthermore, Shtereva et al. (1998) studied the androgenic capability of Lycopersicon esculentum verified that factors as genotype and physiological state from donor plant could act upon plant growth regulators leading to regeneration of plantlets through androgenesis. However, no adventitious buds or somatic embryos were obtained in the present trial.
The anthers oxidation was visually detected in a large scale, although it was not an expected fact. Pacheco-Sanchez et al. (2003) reported that growth regulators can affect the anthers oxidation and the use of ascorbic acid would prevent such effects and would improve the embryogenic responses.
Pre-treatments and Oxidation control: The percentage of calli formation was 25.4% in which 1.2% had any oxidation, 68.1% had partial oxidation and 30.7% had strong level of oxidation. Probably, the oxidation stress could be the main cause in calli formation (Csiszar et al., 2004).
| Table 4: | Responses of calli induction in different levels of plant growth regulators |
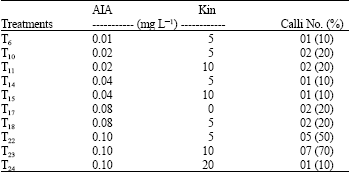 | |
| IAA: Indole-3-acetic acid; KIN: Kinetin | |
The deviance analysis indicated significant differences (p<0.05) for temperature, activated charcoal, plant growth regulators and the interaction between them (data not shown).
Refrigeration (4°C for 48 h continuously) seemed to increase the oxidation level in calli compared to environmental temperature (2.0 and 2.2 oxidized calli, respectively). The data obtained disagree to Fu et al. (2008) that studied anther culture in Chinese pink (Dianthus chinensis L.) and observed that cold pre-treatment (3 days long and 4°C) on flowers buds before removing anthers, increased 4 times the frequency of embryogenic calli in this species. Otherwise, Pacheco-Sanchez et al. (2003) found that low temperatures had significant oxidation effects on preservation and anther survival over the culture period.
However, Perera et al. (2008) found that androgenesis in Cocos nucifera L. was induced after a high temperature pre-treatment (38°C for 6 days), increasing calli formation in 22% however, low temperature pre-treatment (4°C for 1, 3, 6 and 14 days) did not induce androgenesis.
Treatments free of activated charcoal (Table 3) had higher levels of oxidation compared to that ones with presence of activated charcoal: (2.3 and 2.0, mean value, respectively). Ribeiro et al. (2000) studied the influence of activated charcoal in Citrus limonia Osbeck and Poncirus trifoliata (L.) and suggested that inhibitory components acting on growing explants could be adsorbed by the activated charcoal causing a positive effect verified in vitro culture.
Perera et al. (2008) commented the beneficial effects of activated charcoal which is attributed to its adsorption capacity of phenols and other growth inhibitory substances. In anther culture, activated charcoal is reported to remove toxic substances allowing more embryogenic cells to develop.
The analysis of plant growth regulators effects by orthogonal contrasts (Table 5) indicated that 20 mg L-1 of KIN increased the level of oxidation of explants compared to other levels of plant growth regulators. The interactive effects of activated charcoal and plant growth regulators demanded a comparison by orthogonal contrasts within the levels of activated charcoal (Table 6). Growing medium with 20 mg L-1 of KIN and without activated charcoal had the higher values for explant oxidation compared to that ones added with activated charcoal.
Boufleuher et al. (2008) have reported different combinations among auxins and citokinins in order to obtain in vitro propagation in peach-tomato, including calli formation using single nodal segments from shoot-buds, but in this study, calli induction was obtained by anther culture using combinations with KIN and IAA what represents the first study reported in the literature.
Factors affecting calli induction: The induction and the growth of calli from anther culture were visible after 50 days. The calli were soft, white and friable as seen on Fig. 1 evaluated by different treatments. The deviance analysis for callus induction identified significant differences for plant growth regulators and activated charcoal and the interaction between them (p<0.05); pre-refrigeration and activated charcoal had no influence on calli formation. Other researchers reported significant interaction of pre-refrigeration, hormones and activated charcoal in calli formation of Capsicum annuum (Ozkum-Ciner and Tipirdamaz, 2002) but Moraes et al. (2004) reported non-significant thermal effects on soybeans andr ogenesis.
| Table 5: | Orthogonal contrasts for plant growth regulators effects on calli oxidation |
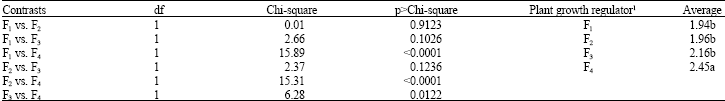 | |
| 1Plant growth regulators − F1: 0 mg L-1, F2: 5 mg L-1, F3: 10 mg L-1 e F4: 20 mg L-1 Kinetin. Mean values with the same letter(s) are not statistically significant | |
| Table 6: | Partition of plant growth regulators effects within the levels of calli oxidation |
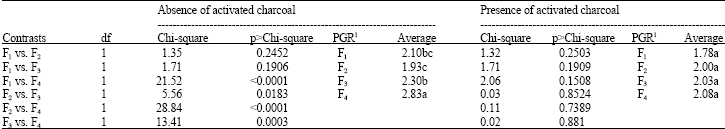 | |
| 1Plant growth regulators − F1: 0 mg L-1, F2: 5 mg L-1, F3: 10 mg L-1 e F4: 20 mg L-1 Kinetin. Mean values with the same letter(s) are not statistically significant | |
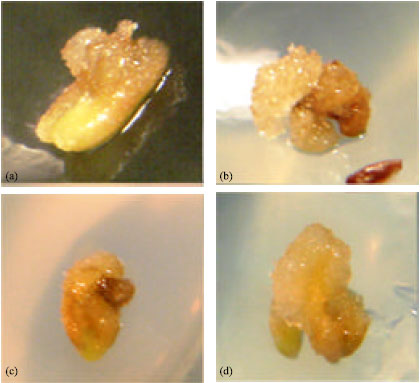 | |
| Fig. 1: | Peach-tomato calli induction from anther culture: (a) Treatment 3 = 0.1 mg L-1 AIA+5 mg L-1 Kin+1 g L-1 Activated charcoal; (b) Treatment 9 = 0.1 mg L-1 +0 mg L-1 Kin; (c) Treatment 10 = 0.1 mg L-1+0 mg L-1 Kin and (d) Treatment 11 = 0.1 mg L-1 AIA+5 mg L-1 Kin |
| Table 7: | Orthogonal contrasts for plant growth regulators effects on calli induction |
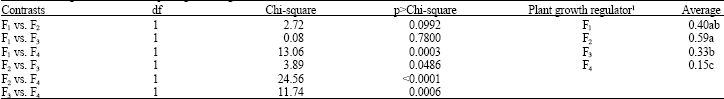 | |
| 1Plant growth regulators − F1: 0 mg L-1, F2: 5 mg L-1, F3: 10 mg L-1 e F4: 20 mg L-1 Kinetin. Mean values with the same letter(s) are not statistically significant | |
| Table 8: | Partition of plant growth regulators effects within the levels of calli induction |
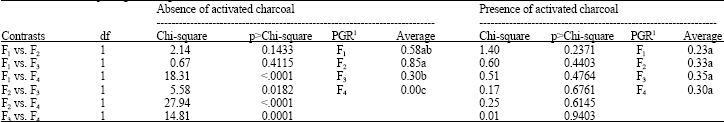 | |
| 1Plant growth regulators − F1: 0 mg L-1, F2: 5 mg L-1, F3: 10 mg L-1 e F4: 20 mg L-1 Kinetin. Mean values with the same letter(s) are not statistically significant | |
According to Pacheco-Sanchez et al. (2003) there was no calli induction in Solanum iopetalum in those treatments under environmental conditions even in the presence or absence of plant growth regulators. Just a few number of embryos obtained were also reported by Ozkum-Ciner and Tipirdamaz (2002) under 4°C for 48 or 96 h with significant differences between the presence or ausence of pre-refrigeration but similar under both periods.
Orthogonal contrasts among the levels of plant growth regulator-Kinetin (Table 7) indicated that calli induction was not different statistically to levels of 0, 5 and 10 mg L-1 of KIN. Only 20 mg L-1 of KIN was statistically different having the worst performance probably due to the higher levels of oxidation.
The partition of plant growth regulators effects within the levels of activated charcoal (Table 8) indicates no significant differences among the concentration of plant growth regulators. In fact, the use of 0, 5 and 10 mg L-1 of KIN promoted calli induction (0.30 to 0.85 mean value) in those treatments free of activated charcoal. These data represents the first research on in vitro response in this species to calli induction from anther culture. The generation of calli from anther culture certainly would contribute to the achievement of homozygous plant regeneration because it represents the first step to obtain homozygous lines. Relevant studies are underway.
CONCLUSION
In conclusion, aseptic cultures were established by flower buds treatment with 70% ethanol for 45 sec followed by sodium hypochlorite 0.8% for 5 min. The best plant hormone treatments for improving calluses formation are 0.1 mg L-1 of IAA+5 mg L-1 or 10 mg L-1 of Kinetin. Furthermore, there was high level of anther oxidation due to temperature, activated charcoal, plant hormones and by the interactive effect plant hormones-by-activated charcoal. Non-activated charcoal and pre-refrigeration at 4°C for 48 h increased the anther oxidation under the high concentration of Kinetin (20 mg L-1).
ACKNOWLEDGMENT
The authors thank the Universidade Paranaense (IPEAC/UNIPAR) for financial support (grant No. 11169).
REFERENCES
- Boufleuher, L.M., A.R. Schuelter, C.L. Luz, C.L. Da-Luz and V.A. Antes et al., 2008. In vitro propagation of Solanum sessiliflorum as affected by auxin and cytokinin combinations and concentrations. Asian J. Plant. Sci., 7: 639-646.
CrossRefDirect Link - Chani, E., R.E. Veilleux and T. Boluarte-Medina, 2000. Improved androgenesis of interspecific potato and efficiency of SSR markers to identify homozygous regenerants. Plant Cell Tissue Organ. Cult., 60: 101-112.
CrossRefDirect Link - Csiszar, J., M. Szabo, L. Erdei, L. Marton, F. Horvath and I. Tari, 2004. Auxin autotrophic tobacco callus tissues resist oxidative stress: The importance of glutathione S-transferase and glutathione peroxidase activities in auxin heterotrophic and autotrophic calli. J. Plant Physiol., 161: 691-699.
CrossRefDirect Link - Fu, X., S. Yang and M. Bao, 2008. Factors affecting somatic embryogenesis in anther cultures of Chinese pink (Dianthus chinensis L.). In Vitro Cell. Dev. Biol. Plant, 44: 194-202.
CrossRef - Gambino, G., P. Ruffa, R. Vallania and I. Gribaudo, 2007. Somatic embryogenesis from whole flowers, anthers and ovaries of grapevine (Vitis spp.). Plant Cell Tissue Organ. Cult., 90: 79-83.
CrossRef - Hern, A. and S. Dorn, 2001. Statistical modelling of insect behavioural response in relation to the chemical composition of test extracts. Physiol. Entomol., 26: 381-390.
CrossRef - Kaltchuk-Santos, E. and M.H. Bodanese-Zanettini, 2002. Androgenesis: An alternative route in the pollen development. Cienc. Rural., 32: 165-173.
CrossRefDirect Link - Moraes, A.P., M.H. Bodanese-Zanettini, S.M. Callegari-Jacques and E. Kaltchuk-Santos, 2004. Effect of temperature shock on soybean microspore embryogenesis. Braz. Arch. Biol. Technol., 47: 537-544.
CrossRef - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Ozkum-Ciner, D. and R. Tipirdamaz, 2002. The effects of cold treatment and charcoal on the in vitro androgenesis of pepper (Capsicum annuum L.). Turk. J. Bot., 26: 131-139.
Direct Link - Pacheco-Sanchez, M., H. Lozoya-Saldana and M. Colinas-Leon, 2003. Reguladores de crecimiento y pretr at amiento con frio en la androgenesis in vitro de Solanum iopetalum L. Agrociencia, 37: 257-265.
Direct Link - Palu, E.G., A.B. Silva and M. Pasqual, 2004. In vitro callogenesis in anthers of Coffea arabica L. Cienc. Agrotec., 28: 736-742.
Direct Link - Pasqual, M., A.L.R. Maciel, K.P. Campos, E.C. Santos and R.J.C. Campos, 2002. Callus induction from anther cultures of coffee (Coffea arabica L.). Cienc. Agrotec., 26: 71-76.
Direct Link - Perera, P.I.P., V. Hocher, J.L. Verdeil, H.D.D. Bandupriya, D.M.D. Yakandawala and L.K. Weerakoon, 2008. Androgenic potential in coconut (Cocos nucifera L.). Plant Cell Tissue Organ Cult., 92: 293-302.
CrossRefDirect Link - Pizzinato, J.R., A.R. Schuelter, A.T. Amaral Jr., A.C.S. Rocha and J.M. Silva et al., 2008. Crossing and diagnostic methods of cubiu hybrid plants base don genetic markers. Crop Breed. Applied Biotechnol., 8: 283-290.
Direct Link - Shtereva, L.A., N.A. Zagorska, B.D. Dimitrov, M.M. Kruleva and H.K. Oanh, 1998. Induced androgenesis in tomato (Lycopersicon esculentum Mill). II. Factors affecting induction of androgenesis. Plant Cell Rep., 18: 312-317.
CrossRefDirect Link - Storti, E.F., 1988. Floral biology of Solanum sessiliflorum Dun. var. sessiliflorum, in the Manaus region, Amazonia. Acta Amazon, 18: 55-65.
Direct Link - Utino, S., I.F. Carneiro and L.J. Chaves, 2001. Explants growth and oxidation of banana cv. prata (Musa AAB) in vitro. I. Iron, Copper and zinc salt concentrations. Rev. Bras. Frutic., 23: 225-229.
Direct Link - Zagorska, N.A., A. Shtereva, B.D. Dimitrov and M.M. Kruleva, 1998. Induced androgenesis in tomato (Lycopersicon esculentum Mill.) I. Influence of genotype on androgenetic ability. Plant Cell Rep., 17: 968-973.
CrossRefDirect Link