
Oliul Hassan
Division of Ecology and Environmental System, College of Ecology and Environmental Sciences, Kyungpook National University,
37224 Sangju-si, Gyeongsangbuk-do, Republic of Korea
Taehyun Chang
Division of Ecology and Environmental System, College of Ecology and Environmental Sciences, Kyungpook National University,
37224 Sangju-si, Gyeongsangbuk-do, Republic of Korea
Asian Journal of Plant Pathology
Year: 2017 | Volume: 11 | Issue: 2 | Page No.: 53-70







ABSTRACT
Due to the high environmental risk of chemical pesticides, biological control of plant diseases with bio-pesticides are highly encouraged and recommend. Chitosan and oligochitosan are the well-known biological control agent for its nontoxic, biodegradable and biocompatible properties. Chitosan is considered as the most abundant natural polymer with a dual effect: It controls pathogenic microorganisms by preventing growth, sporulation, spore viability, germination and disrupting cell and inducement of different defense responses in host plant inducing and/or inhibiting different biochemical activities during the plant-pathogen interaction. Chitosan has been assayed for control numerous pre and post-harvest diseases of many crops. Chitosan also has the positive effect of enriching biodiversity in the rhizosphere. Meteorological effect on chitosan is little evaluated. For achieving the goal of sustainable agriculture, chitosan will become a popular plant protectant.
PDF Abstract XML References Citation
Received: December 19, 2016;
Accepted: February 24, 2017;
Published: March 15, 2017
How to cite this article
Oliul Hassan and Taehyun Chang, 2017. Chitosan for Eco-friendly Control of Plant Disease. Asian Journal of Plant Pathology, 11: 53-70.
URL: https://scialert.net/abstract/?doi=ajppaj.2017.53.70
URL: https://scialert.net/abstract/?doi=ajppaj.2017.53.70
INTRODUCTION
Chemical pesticide is of increasing concern as their negative impact on human health and environment1,2. The broad spectrum pesticides may cause problems by targeting beneficial organisms and repeated use of these synthetic pesticides leads to loss of biodiversity3,4. Some pesticides are not biodegradable and persistence in environment from few to many years for these reasons, they are responsible for soil, water and a wide range of environmental pollution5-7. Some pesticides bio-accumulated in the food chains which is in consequence affecting the human health8,9. There is lots of evidence of the emerging resistance to fungicides due to the indiscriminate use of synthetic fungicides. In these circumstances control of pest with the bio-pesticides is the better alternative to the synthetic ones as the bio-pesticides have good efficacy, low residues and little or no toxicity to non-target organisms and human, causes low environmental pollution and several other advantages10.
Chitin and its derivative, chitosan have been known worldwide for its potential as a biocontrol agent. After cellulose, the second most abundant and important natural biopolymer is chitin in the world11. Chitin has a variety of applications such as in medical science, chemical science and agricultural industry. In medical science, chitin has been used as a wound dressing material, drug carrier and anti-cancer drug etc., while in chemical science it has been used to prepare an affinity chromatography column and to immobilized enzyme etc.11. It has been used as a food additive and bio-pesticides in the agricultural industries12. The most important properties of chitosan such as biodegradability, nontoxicity, biocompatibility and fungicidal effect11,13.
A lot of biological control agents, including natural product (derived from microorganism, higher plant and animal) and microorganisms (fungi, bacteria, virus nematodes and protozoa)10,14 have been used to control the plant diseases and to inhibit the growth of plant pathogens. Chitosan has some advantages over other biocontrol agent not only its, potential to control plant diseases but also its ability to induce resistance in the host plants15 and enhance biodiversity in the rhizosphere16.
The objectives this study were to summarized the efficacy of chitosan and oligo-chitosan on controlling plant diseases, mechanism of induction resistance in plant and influence on microbial community in rhizosphere.
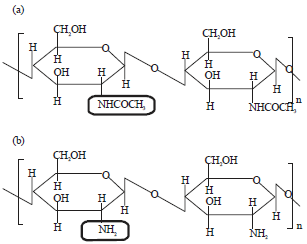 | |
| Fig. 1(a-b): | Structures of (a) Chitin and (b) Chitosan12 |
CHEMICAL STRUCTURE AND FUNCTION OF CHITOSAN
Chitosan is the derivatives of chitin. It is prepared by deacetylation of chitin to varying degree, although the deacetylation is almost never complete. When the deacetylation of chitin is reaching below 50% (depending on the origin of chitin), the product is called chitosan, it became soluble in acidic aqueous media11,17. To convert chitin to chitosan, chitin is N-deacetylated by enzymatic preparation or chemical process. In the chemical process, acids or alkalis (NaOH) are used to deacetylate chitin17,18. In case of enzymatic preparation, chitin deacetylases (EC 3.5.1.41) is used to catalyze the hydrolysis of N-acetamido bonds in chitin to convert it to chitosan17. The structures of chitin and chitosan12 are shown in Fig. 1.
Due to biological (biodegradable, nontoxic and biocompatibility) and chemical (reactive amino group, hydroxyl groups, positive charge, etc.)19 properties, the chitosan has a broad range of industrial applications.
Table 1 summarizes the main application of chitosan in different industries. In this text, we will give emphasis on the use of chitosan in agriculture as elicitors, bio fungicides and modifier of plant-microbial interaction etc.
In vitro ANTIMICROBIAL ACTIVITY
In vitro antifungal activity: There are several techniques that have been used to understand the antimicrobial activities of chitosan. In vitro antifungal activities of chitosan against several genera of fungi have been investigated and documented by several studies. Most of the study revealed that the chitosan affects mycelium growth, sporulation, morphology and molecular organization of fungus.
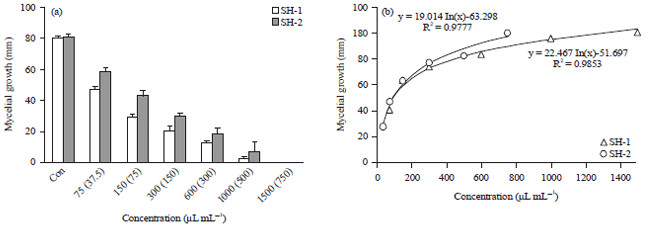 | |
| Fig. 2(a-b): | Effect chitosan preparation concentrations on the rate of mycelial growth and inhibition in V-8 juice agar medium amended with SH-1 (75 μg a.i. mL–1 total glucosamine to 1,500 μg a.i. mL–1 in V-8 juice agar) and SH-2 (37.5 μg a.i. mL–1 total glucosamine to 750 μg a.i. mL–1 in V-8 juice agar) after 10 days of inoculation, (a) Effect of mycelial radial growth and (b) Control value of mycelial radical growth43 |
| Table 1: | Pottential application of chitosan in different field |
 | |
Chitosan effectively inhibits radial mycelium growth, sporulation, spore germination and elongation of Fusarium spp.,32-34, Rhizopus spp.,35,36, Phytophthora spp.,37,38, Botrytis spp.,39,40 and Alternaria spp.,41 etc. Meng et al.42 demonstrated the inhibition rate of mycelium growth of fungi depend on the concentration of chitosan and its derivatives, oligochitosan. Growth inhibition of mycelium increases with the increasing concentration of chitosan preparation SH-1 (75 μg a.i. mL–1 total glucosamine to 1,500 μg a.i. mL–1 in V-8 juice agar) and SH-2 (37.5 μg a.i. mL–1 total glucosamine to 750 μg a.i. mL–1 in V-8 juice agar) after 10 days of inoculation (Fig. 2)43.
In almost all cases, spore-producing fungi produce lower spore when treated with chitosan than untreated fungi. Moreover, in some reports complete sporulation inhibition was observed after treating with chitosan. In contrary to sporulation inhibition, chitosan stimulates sporulation of Penicillium digitatum and Alternaria alternate44,45. The researchers explained this phenomenon by the responses of these fungi against the chitosan induced stress.
The inhibitory effect of chitosan depends on the type of solvent. Chitosan dissolved in lactic acid show the best inhibitory effect as compared to dissolved in formic acid and acetic acid33. Some studies have reported the antifungal activities of chitosan depend on the incubation period. The growth inhibition of F. oxysporum f. sp. radicis-lycopersici decreases with the increased incubation period when treated with chitosan at low concentration (1.0 and 2.0 mg mL–1)46 while for Aspergillus niger, incubation time is very important for chitosan efficacy47. Efficacy of chitosan also depended on the type of chitosan. Low Molecular Weight (LMW) chitosan has the highest inhibitory effect on Rhizopus stolonifera, Botrytis cinerea and Penicillium expansum39,48 while High Molecular Weight (HMW) chitosan show better efficacy on F. oxysporum f. sp. vasinfectum, Alternaria solani and Valsa mali49. All fungus species are not similarly sensitive to chitosan35. Chitosan (0.5 and 1.5%) appears to be more effective in inhibiting mycelium growth of F. oxysporum than P. digitatum and R. stolonifera44. The chitosan could be formulated and applied as a natural antifungal agent into nanoparticles form, to enhance its antifungal activity. Ing et al.50 reported that chitosan nanoparticles prepared from low molecular weight and high molecular weight chitosan have better inhibitory activity against Candida albicans, F. solani. Chitosan silver nanoparticles inhibit conidia germination of Colletotrichum gloeosporioides more effectively than chitosan alone51. Special characters of the chitosan nanoparticles such as small and compact particle as well as high surface charge are responsible for higher antifungal activities than sole chitosan polymer50.
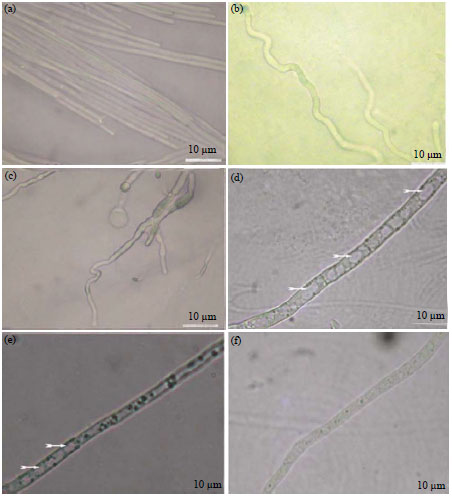 | |
| Fig. 3(a-f): | Effect of chitosan on fungal morphology (a) Hyphal agglomeration, (b-c) Abnormal shapes, (d-e) Large vesicles in the mycelia (arrows) and (f) Normal hyphal growth in control34 |
Effect of chitin, chitosan and its oligomers on fungal morphology is well documented. Scanning electron microscope analysis reveal that inhibition of fungal growth in response to chitosan and its oligomers was accompanied with excessive mycelial branching, abnormal shapes, swelling and hyphae size reduction in Fusarium spp.32,34,46,52, B. cinerea, R. stolonifera, A. alternate and Penicillium expansum35,53,54. Chitosan also responsible for cytological alteration, protoplasm dissolution and large vesicle of fungus34 shown in Fig. 3.
In vitro antibacterial activity: As like as fungi, bacteria also highly sensitive to the chitosan and its derivatives. Most of the chitosan antibacterial reports are on human disease causing bacteria such as Escherichia coli, Staphylococcus aureus and certain Bacillus species. Since chitosan shows the bactericidal activity against a variety of human disease-causing bacteria, it could be expected that chitosan may protect the plant from bacterial diseases. Recently, some researchers revealed that chitosan has strong in vitro antibacterial activities against plant pathogenic bacteria such as Streptomyces scabies55, Xanthomonas56, Pseudomonas syringae57, Pseudomonas fluorescens58, Agrobacterium tumefaciens41, Erwinia41, Burkholderia seminalis59, Acidovorax citrulli60 and Acidovorax avenae61. Inhibitory activity of chitosan against bacteria depends on, concentration41,61 molecular weight of chitosan62 type of bacteria (Gram-positive and Gram-negative)63, type of solvent41, incubation period and abiotic factors64. From these studies, it is clear that chitosan could be used as potential bacteria controlling agent.
In vitro antiviral activity: Despite chitosan is not being a component of the virus, chitosan has been shown to control plant viral diseases. Suppression of viral infection was reported with the tobacco mosaic and necrosis viruses, potato virus X, peanut stunt virus, cucumber mosaic virus and alfalfa mosaic virus65-67. It is hypothesized that chitosan can inhibit viral infection by inducing hypersensitive response, decreasing viability of phage bacterial cells, neutralizing the infectivity of phage particle and blocking the virulent phage replication68.
In vitro nematicidal and anti-protozoal activity: Nematicidal as well as the anti-protozoal activity of chitosan and its derivatives has not been sufficiently studied. From the 1980s onward, a number of studies suggested that chitosan is effective to control plant pathogenic nematode population69-71. Chitinous amendments result in significant reduction in egg hatching, the viability of larvae and adult of the plant pathogenic nematode species Meloidogyne arenaria68, Meloidogyne incognita72, Meloidogyne javanica73 and Heterodera glycines70. The nematicidal activity of chitosan depends on the concentration, molecular weight of chitosan. Khalil and Badawy72 suggested that the chitosan at low molecular weight may serve as a natural nematicide. A few studies have demonstrated that application of chitosan biomolecule is promising option for treatment trichomoniasis74,75. Deacetylated chitosan oligosaccharide significantly decreases the viability of Trichomonas vaginalis74 and Trichomonas gallinae75. Still, a long way to go to make conclusive statements on chitosan as the antiprotozoal agent.
USE OF CHITOSAN FOR CONTROLLING PLANT DISEASE (In vivo STUDY)
Control of Fusarium spp.: Fusarium spp., is a very important plant pathogen causes disease in a wide range of plant, leading significant economic loss and mycotoxin in food and animal feed. At least 81 of 101 economically important crops are affected by Fusarium spp., infection. Chitosan provides a satisfactory level of pant-protection against Fusarium spp., as compared to chemical pesticide33,76,77. To control pathogens, chitosan has been utilized in seed, soil amendment and as foliar treatment78. Fusarium oxysporum the causal agent of crown and root rot disease of tomato. Foliar spraying of chitosan at concentrations ranging from 0.5-2 mg mL–1 markedly reduces the number of root lesions caused by the F. oxysporum79. Bhaskara Reddy et al.78 reported that chitosan treatment (2-8 mg mL–1) of wheat seeds reduces seed born F. graminearum >50%. They also suggested that chitosan treatment increase resistance in seedling and grain yield. Soil amendment with optimal concentrations of chitosan able reduced plant wilting caused by Fusarium spp., especially F. acuminatum80. Hadwiger and Beckman81 found that the tissue is protected from F. solani f. sp., pisi, infection when chitosan applied to pea pod tissue. Fusarium moniliforme is an opportunistic microorganism associated with maize, which is able to colonize in the roots and uptake nutrient. Use of chitosan as maize seed coating agent inhibit the F. moniliforme growth82. Dry rot is an important disease of stored potato caused by F. sulphureum. Li et al.33 reported that chitosan application at the rate of 0.5 or 1% effectively controlled the dry rot of potato tuber.
Control of Phytopthera spp.: According to the results of several studies, Phytopthera spp., is effectively controlled by chitosan. Spraying and soil drench with chitosan reduced Phytopthera infestans43,83,84, Phytophthora cactorum85 and Phytophthora capsici85 diseases. Plant sprayed with N, O-carboxymethyl chitosan was shown to delay late blight development (P. infestans)83. Cho et al.84reported that oligochitosan suppressed the development of late blight on potato with the control value of 72%. Chang and Kim43 after evaluating chitosan preparations (SH-1 and SH-2) on late blight suggested that SH-1 and SH-2 can be used as the natural fungicide in organic farming to increases tuber yield and to control the late blight disease. Phytophthora cactorum is the causal organism for crown rot in strawberry. Effect of chitosan on crown rot disease depends on the concentration of chitosan and time between application and inoculation. Disease severity decrease as the time between treatment and inoculation increased. It may take certain time interval for the plant to install its defenses. Low concentrations of chitosan were more effective against crown rot than high concentrations. Phytophthora root and crown rot of bell pepper caused by Phytophthora capsicum were better controlled when the soil of nursery bed is amended with the chitosan (0.2% w/v)86. Phytophthora parasitica is a root pathogen with a wide range of host. For example, black shank (P. parasitica ) a devastating disease of all types of cultivated tobacco. The result of Falcon et al.87 showed that partially acetylated chitosan was very effective in protecting tobacco against P. parasitica.
Control of Rhizoctonia spp. and Sclerotinia spp.: The potential of chitosan to suppress the diseases caused by Rhizoctonia spp. and Sclerotina spp., has been confirmed various investigations. Rhizoctonia solani is a soil born plant pathogen which is responsible for damping-off, sheath rot and root rot of many plant species. Acid soluble chitosan protects rice seedling from sheath blight (R. solani) and causes a 66-91% inhibition lesion length88. In other studies carried out with lettuce treated with biochikol 020 PC, (2% of chitosan) showing symptoms of damping-off (R. solani) was significantly lower89.
 | |
| Fig. 4: | Protective activity of chitosan preparations on tomato leaf mold103 |
Sclerotinia sclerotiorum is another soil born plant pathogen causing root crown and stem rot in various plant species. Application of chitosan hydrolysate at the rate of 0.2% (w/v) to carrot 3 days before inoculation reduced the size and frequency of Sclerotinia rot as well as induced resistance90. Yin et al.91 found the similar result when oligochitosan when applied to oilseed rape 3 days before inoculation.
Control of Pythium spp., Penicillium spp. and Puccinia spp.: Pythium species cause diseases in a wide range of crops. As for the example root rot of cucumber and rhizome rot disease of turmeric are the very destructive disease caused by Pythium aphanidermatum. El Ghaouth et al.92 investigated the biological activity of chitosan on Pythium rot of cucumber. It is found that chitosan (100-400 mg mL–1) not only control the root rot by adversely affecting the growth of P. aphanidermatum but also stimulated physical barrier and enzymatic defense mechanism without affecting normal growth of cucumber (Fig. 4). Symptom of phytotoxicity also was not found even in higher concentration of chitosan. Anusuya and Sathiyabama93 reported that turmeric plant treated with chitosan (0.1% w/v spraying on leaves) showed increased resistance towards rhizome rot disease caused by P. aphanidermatum.
Blue mould and green mould diseases (caused by several species of Penicillium) cause considerable economic losses on fruit and vegetable during the storing period. Coating fruits and vegetable with chitosan protect them from pathogen and increase their self-life39,94,95. Tomato fruits treated with 0.5 and 1% chitosan could significantly decrease blue mould caused by P. expansum when stored at 2°C and the effects of chitosan on this disease were increase with the increasing concentration of chitosan39. Wang et al.95 found that chitosan significantly inhibits blue mold caused by P. expansum and natural decay on jujube fruit in concentration dependent manner. Coating citrus (Murcott tangor) fruit with chitosan significantly reduced postharvest decay caused by Penicillium. digitatum and Penicillium italicum in respect of molecular weight (high or low)95.
Peanut (Arachis hypogaea) is one of the popular and high demanding crops all over the world for it nutritional value. The study conducted on peanut infection with Puccinia arachidis indicated that chitosan (1000 ppm) has a good controlling effect on leaf rust disease. Chitosan reduced the number of lesions and lesion diameter when applied before inoculation96.
Control of Botrytis spp., Colletotrichum spp. and other fungal species: Botrytis cinerea Pers., a necrotroph plant pathogen, causes gray mould disease in over 200 plant species. Many studies showed that chitosan and its derivatives have the ability to inhibit the development of gray mould in different plant species such as cucumber97, grapes98 and tomato99 etc. The chitosan has the ability decrease in the gray mould disease index from 3.5 in the control plants to 0.45 when spraying on cucumber plants before 24 h of inoculation97. Xu et al.98 evaluated the antifungal activity of grapes seed extract and chitosan, alone or combined, against fungal rot (B. cinerea) of the grapefruits. It is found that GSE and chitosan have synergistic effect and treatment with 1% chitosan plus 0.1% GSE also significantly reduced gray mold incidence flowed by treatment with 1% chitosan coating. As a biocontrol agent chitosan also found effective against tomato gray mold. About 0.2% of chitosan solution significantly inhibited tomato gray mold99.
Colletotrichum anthracnose (Colletotrichum spp.) is one of the most economically important plant disease causing serious economic loss. A wide range of crops is affected by this diseases. The studies that have been conducted to determine the effects of chitosan on Colletotrichum anthracnose revealed that it (chitosan) has satisfiable controlling effect. More than 60% control of anthracnose on papaya fruit is achievable when applied before C. gloeosporioides inoculation100. Significant reduction of anthracnose disease after coating tomato fruits and grape berry with 2.5% chitosan reported by Munoz et al.101. Chitosan also induces resistance in common bean plant hence effectively protected it (common bean) from anthracnose caused by Colletotrichum lindemuthianum102.
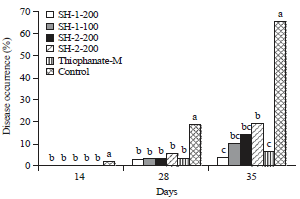
We evaluated the disease control efficacy of two chitosan preparations (SH-1 and SH-2) against tomato leaf mold caused by Fulvia fulva was under plastic greenhouse conditions103. We made SH-1 preparation by mixing chitosan 2 g, chitooligosaccharide 20 g, wood vinegar 20 mL, rice vinegar 40 g and SH-2 by mixing chitosan 2 g, chitooligosaccharide 20 g, wood vinegar 40 mL and rice vinegar 20 g.
| Table 2: | Effect of chitosan on viral infection in different ’Plant-virus’ systems114 |
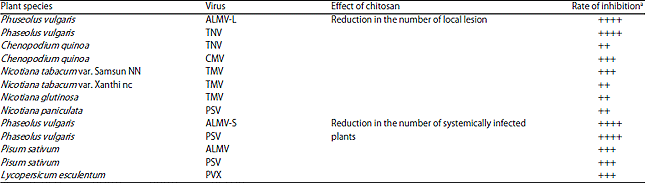 | |
| aRate of inhibition: ++: 20-50%, +++: 50-75%, ++++: 75-100% | |
In this study, we applied chitosan preparation prior to the occurrence of leaf mold on tomato plants and disease severity was rated at different days after three applications at 7 days interval. It is found that both SH-1 and SH-2 formulations displayed potential disease control activity in every case and which was comparable to synthetic thiophanate-M (Fig. 4).
Similarly chitosan potential of controlling fungal diseases such as kernel rot (Aspergillus flavus) in preharvest maize104, root rot (Bipolaris sorokiniana) of wheat65, disease of pears cause by A. kikuchiana and Physalospora piricola42, post-harvest pathogenic fungi (Rhizopus stolonife, Aspergillus niger) in grapes105, brown rot (Monilinia fructicola) in peach fruit106, downy mildew (Sclerospora graminicola) of pearl millet107 and downy mildew (Elsinoe ampelina) and anthracnose (Plasmopara viticola) in grapevines108 was also reported.
Control of Bacterial, viral and nematode disease: Experimental (in vivo) evidence has shown the bactericidal (kill bacteria directly) and bacteriostatic (stop bacteria from growing) ability of chitosan. Soil drench with chitosan significantly reduced Ralstonia wilt (Ralstonia solanacearum) incidence by 72% when the tomato is grown in the greenhouse. Regardless of application of chitosan, it improves the normal growth of tomato109.
The different species of Xanthomonas are responsible for the many destructive diseases in different plant species such as leaf spot of tomato, leaf blight and leaf sreak in rice, bacterial blight of cotton and citrus canker. For evaluating the antibacterial effect of chitosans with different molecular weights on X. gardneri (leaf spot) development and tomato plants protection. Coqueiro and Di Piero110 sprayed chitosan (1 mg mL–1) onto plants 72, 48 or 24 h before inoculation with X. gardneri. It is found that chitosans (low molecular weight, medium molecular weight and commercial) controlled tomato bacterial spot irrespective of molecular weight and reduced disease severity by 70% when applied 72 h before inoculation. Foliar spray of a commercial formulation of chitosan (Armour-Zen®) has the ability to reduce the incidences of Xanthomonas vesicatoria in greenhouse and field grown tomato plants111. The strong antibacterial activity against leaf blight (Xanthomonas oryzae pv. oryzae) and leaf streak (Xanthomonas oryzae pv. oryzicola) of rice also reported by Li et al.112.
Broccoli (Brassica oleracea L. var. italica) is one of the popular vegetables in the world. Growing of broccoli has been seriously hampered by the bacterial head rot (Pseudomonas fluorescens) disease. Regardless of application time (pre or post), six different combinations of chitosan solutions significantly reduced the disease incidence and the lesion diameter of broccoli inoculated with P. fluorescens56. Elexa, the commercial formulation of chitosan effectively protect cucumber from the damage caused by bacterial angular leaf spot (P. lachrymans)113.
To date, a few studies have reported in vivo viral disease control effect of chitosan. The host-virus combination, chitosan concentration and mode of its application are the chitosan efficacy determining factors. Pospieszny et al.114 demonstrated that maximum 99.5-100% alfalfa mosaic virus (ALMV) inhibition is possible when 0.01-0.1% chitosan sprayed into the surface of the leaves before (1 day) inoculation of plants. It is also demonstrated that the varying effect of chitosan on viral infection in different ‘Plant-virus’ systems. The highest level of inhibition of viral infection was observed in bean leaves while TMV, TNV and CMV’s local infection inhibition in tobacco and C. quinoa leaves was not so pronounced in these host (Table 2). Induction of antiviral defensive reactions may varies with the host plant species.
Tobacco Necrosis Virus (TNV) is one of the greatest threats for the cultivation of the tobacco. Chitosan reduces the virus-induced necrotic lesions by 32- 83% when applied on tobacco at the rate of 0.1%115. Chitosan has the ability to delay the viral diseases. In comparison to control plant, chitosan retarded tomato yellow leaf curl disease symptoms about 7-14 days.
A soil amendment with chitosan was shown to reduce nematode parameters, i.e., number of galls, egg masses, females/root system and number of juveniles/250 g soil. El-Sayed et al.73 reported that the standard concentration (1:1 dilution) of LMW and HMW chitosan reduces the number of gall by 90 and 93% respectively. Nunes da Silva et al.116 also reported that application of all molecular weight chitosan decreases the severity of the Pine Wilt Disease (PWD) caused by Bursaphelenchus xylophilus (pine wood nematode). They also reported that HMW chitosan is more effective than LMW chitosan.
MECHANISMS OF CHITOSAN AND ITS DERIVATIVES IN REDUCING PLANT DISEASES
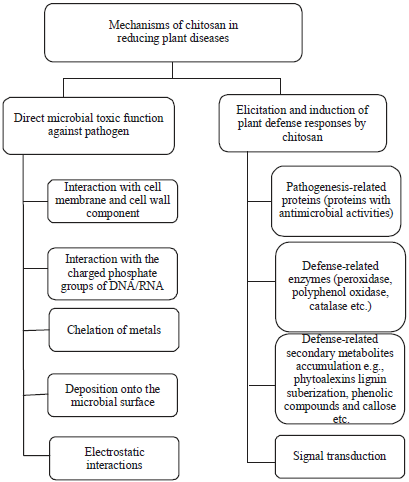
There are a few concrete proofs regarding the anti-microbial mechanism of chitosan and its ability to elicit plant defense responses. The growing evidence is showing that the chitosan and its derivatives have a double mechanism of action: They inhibits the growth of the pathogen and they modify the plant defense responses99,117-119. The possible mechanisms of chitosan as a bio-pesticides is summarized in Fig. 5.
It is hypothesized that the presence of positive charge and reactive amino groups (protonated amino group on the structure of chitosan) are the base of the direct antimicrobial activities of chitosan. The electrostatic interaction between positively charged chitosan and its derivatives with negatively charged microbial cell membranes resulting in agglutination, disruption and alteration of the intracellular ultrastructure of the cell membrane and thereby causing the death organism (Fig. 6)33,120-122.
Chung and Chen123 reported that chitosan react on both the cell membrane and the cell wall and inactivated the pathogen via a two-step sequential mechanism: An initial separation of the cell wall from its cell membrane, followed by destruction of the cell membrane. Chitosan may interfere the function of the genetic material of the pathogen. The nucleic acid (DNA/RNA) possesses the phosphate groups in the main chain which are negatively charged.
 | |
| Fig. 5: | Modes of action of chitosan and its derivatives as antimicrobial compounds and the ability to elicit natural plant defense responses117 |
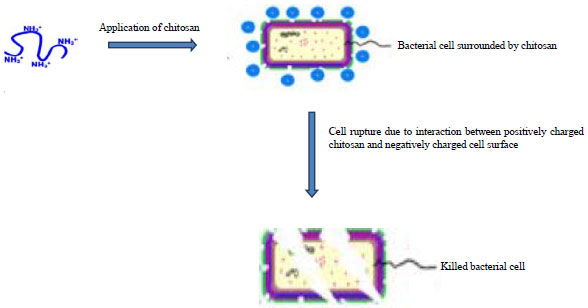 | |
| Fig. 6: | Mechanism of killing of bacterial cell by chitosan122 |
The chitosan could penetrate cell wall17 and bind to the negatively charged DNA and alter its confirmation in consequence block the synthesis of mRNA and essential protein of pathogen89,121,124. The mode of action of chitosan may also be related to its ability to chelate some essential nutrients, metal ions and trace elements necessary for the growth of bacteria and fungi125-127.
Another possible mean of antimicrobial potential of chitosan is to deposit on the pathogen surface and the formation of a dense polymer film. This dense polymer film blocks the nutrient flow and metabolism process of microbes which are essential for their survival in nature117.
Plant protect themselves from pathogens by developing a stunning array of structural, chemical and protein-based defenses designed to recognize invading pathogens and arrest them before they are able to cause extensive damages128.
Some reports indicate that chitosan and its derivatives are effective elicitor and inducer of plant systemic acquired resistance to pathogens. Chitosan and oligochitosan induce hosts to produce defense-related protein117,124, enzymes117,129 and secondary metabolites117. Chitosan and its derivatives increase level and activity of glucanase in rice, tobacco, wheat, etc.15,130,131. It is also reported that chitosan and its derivatives increase, level and activity of chitinase, phenylalanine ammonia-lyase, peroxidase, polyphenol oxidase, catalase and superoxide dismutase in orange, tomato, cucumber, table grapes, strawberries, sweet cherries, pears and Zanthoxylum bungeanum (Huajiao in Chinese)43,117-119. All the mentioned protein and enzymes directly or indirectly degrade the pathogen.
The genes that functionally develop disease resistance are generally called pathogenesis-related genes. Some reports showed that chitosan employs multiple modes to increase pathogenesis-related gene function. As, for example, Hoat et al.132 reported that chitosan and chitin strongly activated the expression of the pathogenesis-related gene (PR-10) in oat leave.
It is well accepted that plant secondary metabolites such as phytoalexins, lignin, suberization and phenolic compounds play a predominant role in plant defense system. The effect of chitosan on the accumulation of defense-related secondary metabolites in the plant tissue was studied from the 1980s when Hadwiger and Beckman81 found chitosan at a concentration as low as 0.9 μg mL–1 can induce a phytoalexin accumulation in soybean pod in 24 h. These biopolymers (chitosan and its derivatives) also have the ability to enhance production and accumulation of phenolic compound (polyphenol), lignin, suberin and callose in different host plant83,78,131,133,134. Accumulation of the polyphenol is the plant part dependent phenomenon. As for example it is found that the content of total polyphenol in the stem was significantly increased by chitosan preparation application in the field (Fig. 7)43.
The resistance plants have the ability of timely recognition of the plant pathogen and the rapid and fruitful activation of the defense system. The activation of defense system manifested by the Hypersensitive Response (HR) at the site of infection and activating defense in uninfected plant part135. The development of HR and the Systematic Acquired Resistance (SAR) govern by the combined genotypes of host and pathogen (gene for gen theory) and depend on a complex exchange of signal occurring under given environmental condition.
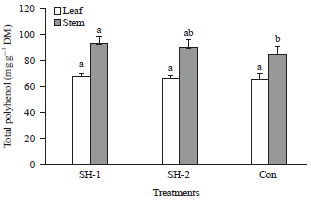 | |
| Fig. 7: | Accumulation of the polyphenol is the plant part after application of chitosan preparation. SH-1 and SH-2 are the chitosan preparation43 |
The signal transduction pathways composed of activation of the specific receptor on the cell membrane or located intracellular followed by transmission of the signal into the target cell mediated by one or more second messengers and create a series of physiological responses. To our knowledge, only the kinase 1 (CERK1) and the lectin are the two cell surface or membrane receptors that are likely to bind chitosan and its oligomers136,137. Chitosan also influences the plant’s DNA conformation. Due to the high affinity for negatively charged phosphates of the DNA backbone, chitosan may alter chromatin via competition with basic nuclear proteins for DNA attachment sites that block gene transcription in turn124,132.
On the basis of the result of several studies, Reactive Oxygen Species (ROS), Ca2+, Nitric Oxide (NO), ethylene (ET), Jasmonic Acid (JA), Salicylic Acid (SA) and abscisic acid (ABA) all involved in chitosan-mediated signal pathway117,138.
It may conclude that chitosan and its derivatives can directly destroy the pathogen and activated the plant immune system through different signaling pathways or involved in signal transduction as a regulatory molecules. Despite extensive research, the mechanism and mode of action of chitosan in reducing plant diseases have not been explicated clearly. However, the mechanisms governing the mode of action of these biopolymers yet to be studied.
ECOLOGICAL CONSIDERATION
Effect of chitosan on environment: Chitosan and its derivatives have emerged as the best ecofriendly bio-pesticides for their unique plant pathogen controlling properties, safety to human and mammals and non-toxicity to other beneficial microbes. The US Environmental Protection Agency (EPA) concluded in 2015139 that there are no risk to the environment because chitosan has not shown toxicity in mammals and it is natural biopolymers. The National Pesticides Information Center (NPIC) database also indicates that there has been no report on environmental degradation due to the use of chitosan.
Effect of chitosan on microbial community in soils: Soil harbours most of the microorganism. Plant interact with soil microorganism in a different way that directly or indirectly influence the growth and development of the plant. Symbiotic microorganism such as mycorrhiza and rhizobia (N-fixing bacteria) promote the growth of the plant by enhancing the plant nutritional status while soil born pathogen limit the plant growth. There are other groups of microorganism which are antagonistic to soil born pathogen hence indirectly influence the plant growth. It is believed that addition of chitosan and its oligomers improves the soil structure and alter the rhizospheric environment to shift the microbial balance in favour of beneficial organisms and to the detriment of plant pathogens24,140.
There is no substantial body of evidence that chitosan supplementation increases both population and efficacy of antagonists in the rhizosphere. Mulawarman et al.141 reported that bacterial population densities of bacterial genera: Acinetobacter and Pseuodomonas increased by 8.9 and 81%, respectively when soil amended with chitosan.
Pastucha142 isolated the greatest number of bacteria and fungi with antagonistic effect towards plant pathogens from rhizosphere soil of soybean after using chitosan three times. Lysobacter enzymogenes is an antagonist biocontrol agent for managing root and crown rot in cucumber caused by Pythium aphanidermatum (Edson) Fitzp. The addition of chitosan (depending on concentration) in soil increases cell number (cells per gram root) on the root as well as the efficacy of L. enzymogenes143. Bacillus subtilis is another, the most widely used biopesticide in agriculture which action against powdery mildew and number of population increases with the addition of chitosan in strawberry144,145. Mishra et al.146 observed the substantially higher number of bacterial cells (Pseudomonas sp.) in tomato roots by scanning electron microscopy after the application of Pseudomonas sp., in combination with chitosan. It is also observed that chitosan enhance the efficacy of Pseudomonas sp., against tomato leaf curl virus (ToLCV) in tomato.
Mycorrhizal fungi, the most ubiquitous of root-associated plant symbioses, enhance the nutrient and water uptake efficiency of the plant. Our previous study16 showed that vesicular-arbuscular mycorrhizal fungi (VAM) significantly increased in soils amended with chitosan powder.
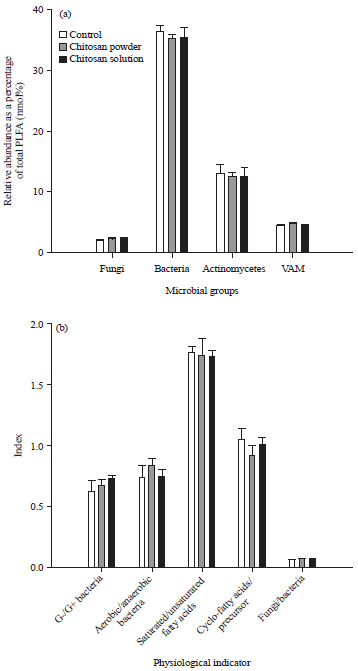 | |
| Fig. 8(a-b): | Effect of chitosan on soil microbial community. Relative abundance of soil microbial groups and physiological parameter were determined by analyzing phospholipid fatty acids (PLFA) in the soils amended with chitosan powder and solution in a cucumber greenhouse at 160 days after soil amendment, (a) Mean microbial group and (b) Physiological indicator of microbes16 |
It is also subsisted that chitosan powder changed the soil microbial community and the effects maintained up to 160 days after soil application (Fig. 8).
In legume nitrogen fixation process, a group of free living bacteria (rhizobia) colonized in the specialized root nodules. The addition of short-chain acetylated chitin derivatives can induce nodulation in Medicago sativa hence, influence the N-fixing bacterial population size and activity147.
Effects of chitosan in stimulating beneficial soil microbial activity can be explained by following major points: (1) Chitosan may serve as nutrient (C and N) source for the microbes142,145, (2) Chitosan may induce the chitinases producing and antagonistic gene expression in biological control agents24,143, (3) Chitosan may induce horizontal gene transfer (natural competence) where DNA was absorbed and recombined into the chitinolytic bacterium and able to being adapted quickly to changing conditions24, (4) Chitosan may help the antagonistic biological control agents to attack plant pathogen by disrupting the formation of biofilms produced by pathogenic microbes24, (5) Chitinous nod (lipochito-oligosaccharides) and Myc (signal molecules) factors in growth-promoting microbes (mycorrhiza and Rhizobium) sensitizes plant roots to nodule formation and to interact with mycorrhizal fungi. Chitin-derivatives which are biochemically similar to nod and myc factors may induce nodulation and the formation of root connections with nitrogen-fixing bacteria and mycorrhizal fungi24,148.
The soil microbial activities are influenced not only by chitosan but also root exudates, organic matter content soil and applied fertilizer149,150. Abundant occurrence of microorganisms in the soil might have been caused by the synergistic effect of chitosan and root exudates, organic matter and fertilizer. So actual effect of chitosan and its derivatives on beneficial soil microbial activities need to be discovered.
Regulation of tolerance to abiotic stresses: Abiotic stresses such as drought, soil pollution with heavy metal, heat and high and low temperature etc., adversely affect the potential production of crops. Chitosan and its derivatives based treatments can improve plant survival and performance in stressful growing environment. Chitosan increases the chlorophyll concentration and performance of photosynthesis under drought condition151,134. Plant need well developed roots system to survive under scare water condition. Zeng and Luo151 reported that chitosan stimulates the root system development of wheat seedling and in consequence strengthen the capability of water absorption. Excessive transpiration is not desirable in dry condition. The concentration of ABA and JA have been found to increase in response to chitosan treatment and these hormones are well known for their ability to control stomatal aperture23,152. Herde et al.152 observed that both ABA and JA reduce the transcription rate in the tomato plant. It is also reported that chitosan induces drought resistance in rice, drought tolerance apple, coffee and paper23,153,154. Bittelli et al.153 found that foliar application of chitosan on pepper leaves induced partial or full closure of plant stomata, resulting in reduced water use by 26-43%.
Under the adverse environmental condition, Reactive Oxygen Species (ROS) such as peroxides, superoxide, hydroxyl radical and singlet oxygen level can increase dramatically. This may cause significant damage to the lipid cell membrane, protein and nucleic acid instead of acting as a messenger in normal cell signal transduction, cell cycling and the plant immune system. The chitosan Reactive Oxygen Species (ROS) scavenging activities were investigated by Je and Kim155. It is found that chitosan showed scavenging effects against hydroxyl and superoxide anion radical, the effects were 91.67 and 65.34%, respectively. These results suggest that the scavenging effect of chitosan depends on their degree of deacetylation and substituted group. Rajalakshmi et al.156 also reported that chitosan and oligo chitosan possess potent antioxidant activity and hydrogen peroxide and superoxide scavenging activity.
The chelating ability of chitosan and oligo-chitosan led to the interest in their potential application to the remediation of contaminated soil and water with heavy metal, dye and accidently application of excess fertilizer and pesticides25,26,157-159. The presence of amino groups (-NH) in chitosan give the vital binding site for complexing with non-nutrient elemental ions, including a number of heavy metal cations157,159. In this way, chitosan protects the plant from the effects of heavy metals, excessive fertilizers and pesticides.
CONCLUSION
In the past few decades, chitosan and its derivatives based pesticides are gaining more attention because of their non-toxic, biodegradable, biocompatible and eco-friendly nature. Chitosan and its derivatives have been repeatedly shown very impressive results as bio-pesticide against many pests of agricultural importance which is comparable to those achieved with current synthetic pesticides and fertilizers. Chitosan is considered as the most abundant natural polymer with a dual effect: It controls pathogenic microorganisms by preventing growth, sporulation, spore viability, germination and disrupting cell and inducement of different defense responses in host plant inducing and/or inhibiting different biochemical activities during the plant-pathogen interaction. The plant can also acquire enhanced tolerance to the stressful growing environment after application of chitosan and oligochitosan. Along with these, the stimulating effect of chitosan on beneficial microorganism lead this natural product to incorporate in to the Integrated Pest Management (IPM) strategies.
There are several issues associated with the commercial use of chitosan based pesticides that are still to be investigated. Like photosensitivity of chitosan derivatives (amorphous chitosan derivatives may absorb UV spectrum) has to be managed for its long lasting effectiveness. The effective concentration and application method of chitosan products is not defined yet. The biochemical mode of action of chitosan and oligochitosan for controlling plant diseases are still limited and unclear. So, much of study is still to be done in this field.
SIGNIFICANCE STATEMENTS
The second most abundant and important natural biopolymer is chitin in the world. Chitin has been used different industries such as in medicine, chemical, cosmetics and agriculture. In the agricultural industry, these biopolymers have been using different ways including biopesticides for controlling plant diseases, bio-transformant for the production of quality food products preservative to protect foods from microbial deterioration, the chelator of waste material from food processing discards, the purifier of water and fruit juices etc. Use of chitosan as biopesticides is very effective and eco-friendly because, it is biodegradable, non-toxic and biocompatible. This study presents a comprehensive review of the antifungal action of and plant protection by chitosan.
ACKNOWLEDGMENTS
This study was supported by a grant from "Cooperative research program for FTA correspondence competitiveness improvement technology development (Project No. PJ011697032016)" in rural development administration, Korea.
REFERENCES
- Jeyaratnam, J., 1985. Health problems of pesticide usage in the third world. Br. J. Ind. Med., 42: 505-506.
Direct Link - Igbedioh, S.O., 1991. Effects of agricultural pesticides on humans, animals and higher plants in developing countries. Arch. Environ. Health, 46: 218-224.
CrossRefPubMedDirect Link - Liebich, J., A. Schaffer and P. Burauel, 2003. Structural and functional approach to studying pesticide side-effects on specific soil functions. Environ. Toxicol. Chem., 22: 784-790.
CrossRefDirect Link - Yasmin, S. and D. D'Souza, 2010. Effects of pesticides on the growth and reproduction of earthworm: A review. Applied Environ. Soil Sci.
CrossRefDirect Link - Bocquene, G. and A. Franco, 2005. Pesticide contamination of the coastline of Martinique. Mar. Pollut. Bull., 51: 612-619.
CrossRefDirect Link - Toan, V.D., V.D. Thao, J. Walder, H.R. Schmutz and T.C. Ha, 2007. Contamination by Selected Organochlorine Pesticides (OCPs) in surface soils in Hanoi, Vietnam. Bull. Environ. Contam. Toxicol., 78: 195-200.
CrossRefDirect Link - Firbank, L.G., S. Petit, S. Smart, A. Blain and R.J. Fuller, 2008. Assessing the impacts of agricultural intensification on biodiversity: A British perspective. Philos. Trans. R. Soc. London B: Biol. Sci., 363: 777-787.
CrossRefDirect Link - Ferencz, L. and A. Balog, 2010. A pesticide survey in soil, water and foodstuffs from Central Romania. Carpath. J. Earth Environ., 5: 111-118.
Direct Link - Cui, L., J. Ge, Y. Zhu, Y. Yang and J. Wang, 2015. Concentrations, bioaccumulation and human health risk assessment of organochlorine pesticides and heavy metals in edible fish from Wuhan, China. Environ. Sci. Pollut. Res., 22: 15866-15879.
CrossRefDirect Link - Copping, L.G. and J.J. Menn, 2000. Biopesticides: A review of their action, applications and efficacy. Pest Manage. Sci., 56: 651-676.
CrossRefDirect Link - Rinaudo, M., 2006. Chitin and chitosan: Properties and applications. Prog. Polym. Sci., 31: 603-632.
CrossRefDirect Link - Zhang, H., R. Li and W. Liu, 2011. Effects of chitin and its derivative chitosan on postharvest decay of fruits: A review. Int. J. Mol. Sci., 12: 917-934.
CrossRefDirect Link - Kumar, M.N.V.R., 2000. A review of chitin and chitosan applications. React. Funct. Polym., 46: 1-27.
CrossRefDirect Link - Yoon, M.Y., B. Cha and J.C. Kim, 2013. Recent trends in studies on botanical fungicides in agriculture. Plant Pathol. J., 29: 1-9.
CrossRefDirect Link - Yin, H., X. Zhao and Y. Du, 2010. Oligochitosan: A plant diseases vaccine-a review. Carbohydr. Polym., 82: 1-8.
CrossRefDirect Link - Park, K.C. and T.H. Chang, 2012. Effect of chitosan on microbial community in soils planted with cucumber under protected cultivation. Korean J. Hortic. Sci. Technol., 30: 261-269.
Direct Link - Younes, I. and M. Rinaudo, 2015. Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs, 13: 1133-1174.
CrossRefDirect Link - No, H.K. and S.P. Meyers, 1995. Preparation and characterization of chitin and chitosan-a review. J. Aquat. Food Prod. Technol., 4: 27-52.
CrossRefDirect Link - Dutta, P.J., J. Dutta and V.S. Tripathi, 2004. Chitin and chitosan: Chemistry, properties and applications. J. Scient. Ind. Res., 63: 20-31.
Direct Link - Miwa, A., A. Ishibe, M. Nakano, T. Yamahira, S. Itai, S. Jinno and H. Kawahara, 1998. Development of novel chitosan derivatives as micellar carriers of taxol. Pharmaceut. Res., 15: 1844-1850.
CrossRefDirect Link - Liu, W., S.J. Sun, X. Zhang and K.D, Yao, 2003. Self-aggregation behavior of alkylated chitosan and its effect on the release of a hydrophobic drug. J. Biomater. Sci. Polym. Edn., 14: 851-859.
CrossRefDirect Link - Dai, T., M. Tanaka, Y.Y. Huang and M.R. Hamblin, 2011. Chitosan preparations for wounds and burns: Antimicrobial and wound-healing effects. Expert Rev. Anti-Infect. Ther., 9: 857-879.
CrossRefDirect Link - Hoagland, P.D. and N. Parris, 1996. Chitosan/pectin laminated films. J. Agric. Food Chem., 44: 1915-1919.
CrossRefDirect Link - Sharp, R.G., 2013. A review of the applications of chitin and its derivatives in agriculture to modify plant-microbial interactions and improve crop yields. Agronomy, 3: 757-793.
CrossRefDirect Link - Sobahi, T.R.A., M.Y. Abdelaal and M.S.I. Makki, 2014. Chemical modification of Chitosan for metal ion removal. Arab. J. Chem., 7: 741-746.
CrossRefDirect Link - Zhang, L., Y. Zeng and Z. Cheng, 2016. Removal of heavy metal ions using Chitosan and modified Chitosan: A review. J. Mol. Liquids, 214: 175-191.
CrossRefDirect Link - Hirano, S., 1996. Chitin biotechnology applications. Biotechnol. Annu. Rev., 2: 237-258.
CrossRefDirect Link - Sapers, G.M., 1992. Chitosan enhances control of enzymatic browning in apple and pear juice by filtration. J. Food Sci., 57: 1192-1193.
CrossRefDirect Link - Roller, S. and N. Covill, 1999. The antifungal properties of chitosan in laboratory media and apple juice. Int. J. Food Microbiol., 47: 67-77.
CrossRefDirect Link - Shahidi, F., J.K.V. Arachchi and Y.J. Jeon, 1999. Food applications of chitin and chitosans-A review. Trends Food Sci. Technol., 10: 37-51.
CrossRefDirect Link - Jimtaisong, A. and N. Saewan, 2014. Utilization of carboxymethyl chitosan in cosmetics. Int. J. Cosmet. Sci., 36: 12-21.
CrossRefDirect Link - El Hassni, M., A. El Hadrami, F. Daayf, E.A. Barka and I. El Hadrami, 2004. Chitosan, antifungal product against Fusarium oxysporum f. sp. albedinis and elicitor of defence reactions in date palm roots. Phytopathologia Mediterranea, 43: 195-204.
Direct Link - Li, Y.C., X.J. Sun, Y. Bi, Y.H. Ge and Y. Wang, 2009. Antifungal activity of Chitosan on Fusarium sulphureum in relation to dry rot of potato tuber. Agric. Sci. China, 8: 597-604.
CrossRefDirect Link - Al-Hetar, M.Y., M.A. Zainal Abidin, M. Sariah and M.Y. Wong, 2011. Antifungal activity of chitosan against Fusarium oxysporum f. sp. Cubense. J. Applied Polym. Sci., 120: 2434-2439.
CrossRefDirect Link - El Ghaouth, A., J. Arul, J. Grenier and A. Asselin, 1992. Antifungal activity of chitosan on two postharvest pathogens of strawberry fruits. Phytopathology, 82: 398-402.
CrossRefDirect Link - Hernandez-Lauzardo, A.N., M.G. Velazquez-del Valle, L. Veranza-Castelan, G.E. Melo-Giorgana and M.G. Guerra-Sanchez, 2010. Effect of chitosan on three isolates of Rhizopus stolonifer obtained from peach, papaya and tomato. Fruits, 65: 245-253.
CrossRefDirect Link - Xu, J., X. Zhao, X. Han and Y. Du, 2007. Antifungal activity of oligochitosan against Phytophthora capsici and other plant pathogenic fungi in vitro. Pestic. Biochem. Physiol., 87: 220-228.
CrossRefDirect Link - Sunpapao, A. and C. Pornsuriya, 2014. Effects of chitosan treatments on para rubber leaf fall disease caused by Phytophthora palmivora Butler-a laboratory study. Songklanakarin J. Sci. Technol., 36: 507-512.
Direct Link - Liu, J., S.P. Tian, X. Meng and Y. Xu, 2007. Effects of chitosan on control of postharvest diseases and physiological responses of tomato fruit. Postharvest Biol. Technol., 44: 300-306.
CrossRefDirect Link - Silva, Jr. S., N.P. Stamford, M.A.B. Lima, T.M.S. Arnaud, M.M. Pintado and B.F. Sarmento, 2014. Characterization and inhibitory activity of chitosan on hyphae growth and morphology of Botrytis cinerea plant pathogen. Int. J. Applied Res. Nat. Prod., 7: 31-38.
Direct Link - Rabea, E.I. and W. Steurbaut, 2010. Chemically modified chitosans as antimicrobial agents against some plant pathogenic bacteria and fungi. Plant Protect. Sci., 46: 149-158.
Direct Link - Meng, X., L. Yang, J.F. Kennedy and S. Tian, 2010. Effects of chitosan and oligochitosan on growth of two fungal pathogens and physiological properties in pear fruit. Carbohydr. Polym., 81: 70-75.
CrossRefDirect Link - Chang, T. and B.S. Kim, 2012. Application of chitosan preparations for eco-friendly control of potato late blight. Res. Plant Dis., 18: 338-348.
CrossRefDirect Link - Bautista-Banos, S., M. Hernandez-Lopez and E. Bosquez-Molina, 2004. Growth inhibition of selected fungi by chitosan and plant extracts. Revista Mexicana Fitopatologia, 22: 178-186.
Direct Link - Bhaskara Reddy, M.V., J. Arul, E. Ait-Barka, P. Angers, C. Richard and F. Castaigne, 1998. Effect of chitosan on growth and toxin production by Alternaria alternata f. sp. lycopersici . Biocontrol Sci. Technol., 8: 33-43.
CrossRefDirect Link - Benhamou, N., 1992. Ultrastructural and cytochemical aspects of chitosan on Fusarium oxysporum f. sp. radicis-lycopersici, agent of tomato crown and root rot. Phytopathology, 82: 1185-1193.
Direct Link - Plascencia-Jatomea, M., G. Viniegra, R. Olayo, M.M. Castillo-Ortega and K. Shirai, 2003. Effect of chitosan and temperature on spore germination of Aspergillus niger. Macromol. Biosci., 3: 582-586.
CrossRefDirect Link - Hernandez-Lauzardo, A.N., S. Bautista-Banos, M.G. Velazquez-del Valle, M.G. Mendez-Montealvo, M.M. Sanchez-Rivera and L.A. Bello-Perez, 2008. Antifungal effects of chitosan with different molecular weights on in vitro development of Rhizopus stolonifer (Ehrenb.:Fr.) Vuill. Carbohydr. Polym., 73: 541-547.
CrossRefDirect Link - Guo, Z., R. Chen, R. Xing, S. Liu and H. Yu et al., 2006. Novel derivatives of chitosan and their antifungal activities in vitro. Carbohydr. Res., 341: 351-354.
CrossRefPubMedDirect Link - Ing, L.Y., N.M. Zin, A. Sarwar and H. Katas, 2012. Antifungal activity of chitosan nanoparticles and correlation with their physical properties. Int. J. Biomater.
CrossRef - Chowdappa, P., S. Gowda, C.S. Chethana and S. Madhura, 2014. Antifungal activity of chitosan-silver nanoparticle composite against Colletotrichum gloeosporioides associated with mango anthracnose. Afr. J. Microbiol. Res., 8: 1803-1812.
CrossRefDirect Link - Prapagdee, B., K. Kotchadat, A. Kumsopa and N. Visarathanonth, 2007. The role of chitosan in protection of soybean from sudden death syndrome caused by Fusarium solani f. sp. glycines. Bioresour. Technol., 98: 1353-1358.
CrossRefDirect Link - De Oliveira, Jr. E.N., I.S. de Melo and T.T. Franco, 2012. Changes in hyphal morphology due to chitosan treatment in some fungal species. Braz. Arch. Biol. Technol., 55: 637-646.
CrossRefDirect Link - Alfaro-Gutierrez, I.C., M.G. Guerra-Sanchez, A.N. Hernandez-Lauzardo and M.G. Velazquez-del Valle, 2014. Morphological and physiological changes on Rhizopus stolonifer by effect of chitosan, oligochitosan or essential oils. J. Phytopathol., 162: 723-730.
CrossRefDirect Link - Beausejour, J., N. Clermont and C. Beaulieu, 2003. Effect of Streptomyces melanosporofaciens strain EF-76 and of chitosan on common scab of potato. Plant Soil, 256: 463-468.
CrossRefDirect Link - Li, B., X. Wang, R. Chen, W. Huangfu and G.L. Xie, 2008. Antibacterial activity of chitosan solution against Xanthomonas pathogenic bacteria isolated from Euphorbia pulcherrima. Carbohydr. Polym., 72: 287-292.
CrossRefDirect Link - Ferrante, P. and M. Scortichini, 2010. Molecular and phenotypic features of Pseudomonas syringae pv. actinidiae isolated during recent epidemics of bacterial canker on yellow kiwifruit (Actinidia chinensis) in central Italy. Plant Pathol., 59: 954-962.
CrossRefDirect Link - Li, B., B.P. Liu, T. Su, Y. Wang and G. Xie et al., 2010. Effect of chitosan solution on the inhibition of Pseudomonas fluorescens causing bacterial head rot of broccoli. Plant Pathol. J., 26: 189-193.
CrossRefDirect Link - Lou, M.M., B. Zhu, I. Muhammad, B. Li and G.L. Xie et al., 2011. Antibacterial activity and mechanism of action of chitosan solutions against apricot fruit rot pathogen Burkholderia seminalis. Carbohydr. Res., 346: 1294-1301.
CrossRefDirect Link - Li, B., S. Shi, C. Shan, Q. Zhou and M. Ibrahim et al., 2013. Effect of chitosan solution on the inhibition of Acidovorax citrulli causing bacterial fruit blotch of watermelon. J. Sci. Food Agric., 93: 1010-1015.
CrossRefPubMedDirect Link - Yang, C., B. Li, M. Ge, K. Zhou and Y. Wang et al., 2014. Inhibitory effect and mode of action of chitosan solution against rice bacterial brown stripe pathogen Acidovorax avenae subsp. avenae RS-1. Carbohydr. Res., 391: 48-54.
CrossRefDirect Link - Liu, N., X.G. Chen, H.J. Park, C.G. Liu, C.S. Liu, X.H. Meng and L.J. Yu, 2006. Effect of MW and concentration of chitosan on antibacterial activity of Escherichia coli. Carbohydr. Polym., 64: 60-65.
CrossRefDirect Link - Shanmugam, A., K. Kathiresan and L. Nayak, 2016. Preparation, characterization and antibacterial activity of chitosan and phosphorylated chitosan from cuttlebone of Sepia kobiensis (Hoyle, 1885). Biotechnol. Rep., 9: 25-30.
CrossRefDirect Link - Chung, Y.C., H.L. Wang, Y.M. Chen and S.L. Li, 2003. Effect of abiotic factors on the antibacterial activity of chitosan against waterborne pathogens. Bioresour. Technol., 88: 179-184.
CrossRefPubMedDirect Link - Badawy, M.E.I. and E.I. Rabea, 2011. A biopolymer chitosan and its derivatives as promising antimicrobial agents against plant pathogens and their applications in crop protection. Int. J. Carbohydr. Chem., Vol. 2011.
CrossRefDirect Link - Nagorskaya, V., A. Reunov, L. Lapshina, V. Davydova and I. Yermak, 2014. Effect of chitosan on Tobacco Mosaic Virus (TMV) accumulation, hydrolase activity and morphological abnormalities of the viral particles in leaves of N. tabacum L. cv. Samsun. Virologica Sinica, 29: 250-256.
CrossRefDirect Link - Jia, X., Q. Meng, H. Zeng, W. Wang and H. Yin, 2016. Chitosan oligosaccharide induces resistance to Tobacco mosaic virus in Arabidopsis via the salicylic acid-mediated signalling pathway. Scient. Rep., Vol. 6.
CrossRefDirect Link - Chirkov, S.N., 2002. The antiviral activity of chitosan (review). Applied Biochem. Microbiol., 38: 1-8.
CrossRefDirect Link - Mian, I.H., O. Godoy, R.A. Shelby, R. Rodriguez-Kabana and G. Morgan-Jones, 1982. Chitin amendments for control of Meloidogyne arenaria in infested soil. Nematropica, 12: 71-84.
Direct Link - Rodriguez-Kabana, R., G. Morgan-Jones and B.O. Gintis, 1984. Effects of chitin amendments to soil on Heterodera glycines, microbial populations and colonization of cysts by fungi. Nematropica, 14: 10-25.
Direct Link - Kalaiarasan, P., P. Lakshmanan, G. Rajendran and R. Samiyappan, 2006. Chitin and chitinolytic biocontrol agents for the management of root knot nematode, Meloidogyne arenaria in groundnut (Arachis hypogaea L.) cv. Co3. Indian J. Nematol., 36: 181-186.
Direct Link - Khalil, M.S. and M.E.I. Badawy, 2012. Nematicidal activity of a biopolymer chitosan at different molecular weights against root-knot nematode, Meloidogyne incognita. Plant Protect. Sci., 48: 170-178.
Direct Link - El-Sayed, S.M. and M.E. Mahdy, 2015. Effect of chitosan on root-knot nematode, Meloidogyne javanica on tomato plants. Int. J. ChemTech Res., 7: 1985-1992.
Direct Link - Shin, W.S., J.C. Kil and G.M. Park, 2006. Antiprotozoal activity of deacetylated chitosan oligosaccharide (dp 2-8) on Trichomonas vaginalis. J. Microbiol. Biotechnol., 16: 1984-1989.
Direct Link - Tavassoli, M., A. Imani, H. Tajik, M. Moradi and S.H. Pourseyed, 2012. Novel in vitro efficiency of chitosan biomolecule against Trichomonas gallinae. Iran. J. Parasitol., 7: 92-96.
Direct Link - Benhamou, N., P.J. Lafontaine and M. Nicole, 1994. Induction of systemic resistance to Fusarium crown and root rot in tomato plants by seed treatment with chitosan. Phytopathology, 84: 1432-1444.
CrossRefDirect Link - Benhamou, N., J.W. Kloepper and S. Tuzun, 1998. Induction of resistance against Fusarium wilt of tomato by combination of chitosan with an endophytic bacterial strain: Ultrastructure and cytochemistry of the host response. Planta, 204: 153-168.
CrossRefDirect Link - Bhaskara Reddy, M.V., J. Arul, P. Angers and L. Couture, 1999. Chitosan treatment of wheat seeds induces resistance to Fusarium graminearum and improves seed quality. J. Agric. Food Chem., 47: 1208-1216.
CrossRefPubMedDirect Link - Benhamou, N. and G. Theriault, 1992. Treatment with chitosan enhances resistance of tomato plants to the crown and root rot pathogen Fusarium oxysporum f. sp. radicis-lycopersici. Physiol. Mol. Plant Pathol., 41: 33-52.
CrossRefDirect Link - Laflamme, P., N. Benhamou, N. Bussieres and M. Dessureault, 2000. Differential effect of chitosan on root rot fungal pathogens in forest nurseries. Can. J. Bot., 77: 1460-1468.
CrossRefDirect Link - Hadwiger, L.A. and J.M. Beckman, 1980. Chitosan as a component of pea-Fusarium solani interactions. Plant Physiol., 66: 205-211.
CrossRefDirect Link - Lizarraga-Paulin, E.G., I. Torres-Pacheco, E. Moreno-Martinez and S.P. Miranda-Castro, 2011. Chitosan application in maize (Zea mays) to counteract the effects of abiotic stress at seedling level. Afr. J. Biotechnol., 10: 6439-6446.
Direct Link - O'Herlihy, E.A., E.M. Duffy and A, C. Cassells, 2003. The effects of arbuscular mycorrhizal fungi and chitosan sprays on yield and late blight resistance in potato crops from microplants. Folia Geobotanica, 38: 201-207.
CrossRefDirect Link - Cho, Y.H., G.J. Choi, B.S. Kim, K.S. Jang, M.Y. Yoon, M.S. Park and J.C Kim, 2011. Control of late blight of tomato and potato by oilgochitosan. Res. Plant Dis., 17: 129-135.
CrossRefDirect Link - Eikemo, H., A. Stensvand and A.M. Tronsmo, 2003. Induced resistance as a possible means to control diseases of strawberry caused by Phytophthora spp. Plant Dis., 87: 345-350.
CrossRefDirect Link - Kim, K.D., S. Nemec and G. Musson, 1997. Control of Phytophthora root and crown rot of bell pepper with composts and soil amendments in the greenhouse. Applied Soil Ecol., 5: 169-179.
CrossRefDirect Link - Falcon, A.B., J.C. Cabrera, D. Costales, M.A. Ramirez, G. Cabrera, V. Toledo and M.A. Martinez-Tellez, 2008. The effect of size and acetylation degree of chitosan derivatives on tobacco plant protection against Phytophthora parasitica nicotianae. World J. Microbiol. Biotechnol., 24: 103-112.
CrossRefDirect Link - Liu, H., W. Tian, B. Li, G. Wu and M. Ibrahim et al., 2012. Antifungal effect and mechanism of chitosan against the rice sheath blight pathogen, Rhizoctonia solani. Biotechnol. Lett., 34: 2291-2298.
CrossRefDirect Link - Molloy, C., L.H. Cheah and J.P. Koolaard, 2004. Induced resistance against Sclerotinia sclerotiorum in carrots treated with enzymatically hydrolysed chitosan. Postharvest Biol. Technol., 33: 61-65.
CrossRefDirect Link - Yin, H., Y. Li, H.Y. Zhang, W.X. Wang and H. Lu et al., 2013. Chitosan oligosaccharides-triggered innate immunity contributes to oilseed rape resistance against Sclerotinia sclerotiorum. Int. J. Plant. Sci., 174: 722-732.
CrossRefDirect Link - El Ghaouth, L.A., J. Arul, N. Benhamou, A. Assselin and R.R. Belanger, 1994. Effect of chitosan on cucumber plants: Suppression of Pythium aphanidermatum and induction of defense reactions. Phytopathology, 84: 313-320.
Direct Link - Anusuya, S. and M. Sathiyabama, 2014. Effect of chitosan on rhizome rot disease of turmeric caused by Pythium aphanidermatum. ISRN Biotechnol.
CrossRefDirect Link - Chien, P.J., F. Sheu and H.R. Lin, 2007. Coating citrus (Murcott tangor) fruit with low molecular weight chitosan increases postharvest quality and shelf life. Food Chem., 100: 1160-1164.
CrossRefDirect Link - Wang, L., H. Wu, G. Qin and X. Meng, 2014. Chitosan disrupts Penicillium expansum and controls postharvest blue mold of jujube fruit. Food Control, 41: 56-62.
CrossRefDirect Link - Sathiyabama, M. and R. Balasubramanian, 1998. Chitosan induces resistance components in Arachis hypogaea against leaf rust caused by Puccinia arachidis Speg. Crop Protec., 17: 307-313.
CrossRefDirect Link - Ben-Shalom, N., R. Ardi, R., Pinto, C. Aki and E. Fallik, 2003. Controlling gray mould caused by Botrytis cinerea in cucumber plants by means of chitosan. Crop Protect., 22: 285-290.
CrossRefDirect Link - Xu, W.T., K.L. Huang, F. Guo, W. Qu, J.J. Yang, Z.H. Liang and Y.B. Luo, 2007. Postharvest grapefruit seed extract and chitosan treatments of table grapes to control Botrytis cinerea. Postharvest Biol. Technol., 46: 86-94.
CrossRefDirect Link - Zhang, D., H. Wang, Y. Hu and Y. Liu, 2015. Chitosan controls postharvest decay on cherry tomato fruit possibly via the mitogen-activated protein kinase signaling pathway. J. Agric. Food Chem., 63: 7399-7404.
CrossRefDirect Link - Bautista-Banos, S., M. Hernandez-Lopez, E. Bosquez-Molina and C.L. Wilson, 2003. Effects of chitosan and plant extracts on growth of Colletotrichum gloeosporioides, anthracnose levels and quality of papaya fruit. Crop Protect., 22: 1087-1092.
CrossRefDirect Link - Munoz, Z., A. Moret and S. Garces, 2009. Assessment of chitosan for inhibition of Colletotrichum sp. on tomatoes and grapes. Crop Protect., 28: 36-40.
CrossRefDirect Link - Di Piero, R.M. and M.V. Garda, 2008. Chitosan reduces the anthracnose severity and increases the glucanase activity in common bean plants. Pesquisa Agropecuaria Brasileira, 43: 1121-1128.
CrossRefDirect Link - Chang, T.H., 2009. Disease control efficacy of chitosan preparations against tomato leaf mold. Res. Plant Dis., 15: 248-253.
CrossRefDirect Link - Cuero, R.G., E. Duffus, G. Osuji and R. Pettit, 1991. Aflatoxin control in preharvest maize: Effects of chitosan and two microbial agents. J. Agric. Sci., 117: 165-169.
CrossRefDirect Link - Dos Santos, N.S.T., A.J.A.A. Aguiar, C.E.V. de Oliveira, C.V. de Sales and S. de Melo e Silva et al., 2012. Efficacy of the application of a coating composed of chitosan and Origanum vulgare L. essential oil to control Rhizopus stolonifer and Aspergillus niger in grapes (Vitis labrusca L.). Food Microbiol., 32: 345-353.
CrossRefDirect Link - Yang, L.Y., J.L. Zhang, C.L. Bassett and X.H. Meng, 2012. Difference between chitosan and oligochitosan in growth of Monilinia fructicola and control of brown rot in peach fruit. LWT-Food Sci. Technol., 46: 254-259.
CrossRefDirect Link - Manjunatha, G., K.S. Roopa, G.N. Prashanth and H.S. Shetty, 2008. Chitosan enhances disease resistance in pearl millet against downy mildew caused by Sclerospora graminicola and defence-related enzyme activation. Pestic. Manage. Sci., 64: 1250-1257.
CrossRefDirect Link - Maia, A.J., R.V. Botelho, C.M.D.R. Faria and C.D. Leite, 2010. Chitosan action on Plasmopara viticola and Elsinoe ampelina development in vitro and in grapevines cv. Isabel. Summa Phytopathologica, 36: 203-209.
CrossRefDirect Link - Algam, S.A.E., G.L. Xie, B. Li, S.H. Yu, T. Su and J. Larsen, 2010. Effects of Paenibacillus strains and chitosan on plant growth promotion and control of Ralstonia wilt in tomato. J. Plant Pathol., 92: 593-600.
Direct Link - Coqueiro, D.S.O. and M.R. di Piero, 2011. Antibiotic activity against Xanthomonas gardneri and protection of tomato plants by chitosan. J. Plant Pathol., 93: 337-344.
Direct Link - Ramkissoon, A., J. Francis, V. Bowrin, R. Ramjegathesh, A. Ramsubhag and J. Jayaraman, 2016. Bio‐efficacy of a chitosan based elicitor on Alternaria solani and Xanthomonas vesicatoria infections in tomato under tropical conditions. Ann. Applied Biol., 169: 274-283.
CrossRefDirect Link - Li, B., B. Liu, C. Shan, M. Ibrahim and Y. Lou et al., 2013. Antibacterial activity of two chitosan solutions and their effect on rice bacterial leaf blight and leaf streak. Pest Manage. Sci., 69: 312-320.
CrossRefDirect Link - Pospieszny, H. S. Chirkov and J. Atabekov, 1991. Induction of antiviral resistance in plants by chitosan. Plant Sci., 79: 63-68.
CrossRefDirect Link - Iriti, M., M. Sironi, S. Gomarasca, A.P. Casazza, C. Soave and F. Faoro, 2006. Cell death-mediated antiviral effect of chitosan in tobacco. Plant Physiol. Biochem., 44: 893-900.
CrossRefDirect Link - Nunes da Silva, M., A.R. Cardoso, D. Ferreira, M. Brito, M.E. Pintado and M.W. Vasconcelos, 2014. Chitosan as a biocontrol agent against the pinewood nematode (Bursaphelenchus xylophilus). For. Pathol., 44: 420-423.
CrossRefDirect Link - Xing, K., X. Zhu, X. Peng and S. Qin, 2015. Chitosan antimicrobial and eliciting properties for pest control in agriculture: A review. Agron. Sustain. Dev., 35: 569-588.
CrossRefDirect Link - Romanazzi, G., 2010. Chitosan treatment for the control of postharvest decay of table grapes, strawberries and sweet cherries. Fresh Produce, 4: 111-115.
Direct Link - Li, P., Z. Cao, Z. Wu, X. Wang and X. Li, 2016. The effect and action mechanisms of oligochitosan on control of stem dry rot of Zanthoxylum bungeanum. Int. J. Mol. Sci., Vol. 17.
CrossRefDirect Link - Sudarshan, N.R., D.G. Hoover and D. Knorr, 1992. Antibacterial action of chitosan. Food Biotechnol., 6: 257-272.
CrossRefDirect Link - Helander, I.M., E.L. Nurmiaho-Lassila, R. Ahvenainen, J. Rhoades and S. Roller, 2001. Chitosan disrupts the barrier properties of the outer membrane of gram-negative bacteria. Int. J. Food Microbiol., 71: 235-244.
CrossRefPubMedDirect Link - Chopra, H. and G. Ruhi, 2016. Eco friendly chitosan: An efficient material for water purification. Pharm. Innov. J., 5: 92-95.
Direct Link - Chung, Y.C. and C.Y. Chen, 2008. Antibacterial characteristics and activity of acid-soluble chitosan. Bioresour. Technol., 99: 2806-2814.
CrossRefDirect Link - Hadwiger, L.A., 2013. Multiple effects of chitosan on plant systems: Solid science or hype. Plant Sci., 208: 42-49.
CrossRefDirect Link - Qin, Y., 1993. The chelating properties of chitosan fibers. J. Applied Polym. Sci., 49: 727-731.
CrossRefDirect Link - Rabea, E.I., M.E.T. Badawy, C.V. Stevens, G. Smagghe and W. Steurbaut, 2003. Chitosan as antimicrobial agent: Applications and mode of action. Biomacromolecules, 4: 1457-1465.
CrossRefDirect Link - Bhuvaneshwari, S. and V. Sivasubramanian, 2013. Comparative studies for Chitosan yield and chelating ability of Aspergillus niger and Rhizopus oryzae. Indian J. Biotechnol., 12: 429-431.
Direct Link - Freeman, B.C. and G.A. Beattie, 2008. An overview of plant defenses against pathogens and herbivores. Plant Health Instructor.
CrossRefDirect Link - Lin, W., X. Hu, W. Zhang, W.J. Rogers and W. Cai, 2005. Hydrogen peroxide mediates defence responses induced by Chitosans of different molecular weights in rice. J. Plant Physiol., 162: 937-944.
CrossRefDirect Link - Eilenberg, H., S. Pnini-Cohen, Y. Rahamim, E. Sionov, E. Segal, S. Carmeli and A. Zilberstein, 2009. Induced production of antifungal naphthoquinones in the pitchers of the carnivorous plant Nepenthes khasiana. J. Exp. Bot., 61: 911-922.
CrossRefDirect Link - Burkhanova, G.F., L.G. Yarullina and I.V. Maksimov, 2007. The control of wheat defense responses during infection with Bipolaris sorokiniana by chitooligosaccharides. Russian J. Plant Physiol., 54: 104-110.
CrossRefDirect Link - Hoat, T.X., H. Nakayashiki, Q. Yang, Y. Tosa and S. Mayama, 2013. Molecular cloning of the apoptosis-related calcium-binding protein AsALG-2 in Avena sativa. Mol. Plant Pathol., 14: 222-229.
CrossRefDirect Link - Fan, G., X. Li, X. Wang, Q. Zhai and Y. Zhan, 2010. Chitosan activates defense responses and triterpenoid production in cell suspension cultures of Betula platyphylla suk. Afr. J. Biotechnol., 9: 2816-2820.
Direct Link - Katiyar, D., A. Hemantaranjan and B. Singh, 2015. Chitosan as a promising natural compound to enhance potential physiological responses in plant: A review. Indian J. Plant Physiol.
CrossRefDirect Link - Petutschnig, E.K., A.M. Jones, L. Serazetdinova, U. Lipka and V. Lipka, 2010. The Lysin Motif Receptor-Like Kinase (LysM-RLK) CERK1 is a major chitin-binding protein in Arabidopsis thaliana and subject to chitin-induced phosphorylation. J. Biol. Chem., 285: 28902-28911.
CrossRefDirect Link - Lienart, Y., C. Gautier and A. Domard, 1991. Isolation from Rubus cell-suspension cultures of a lectin specific for glucosamine oligomers. Planta, 184: 8-13.
CrossRefDirect Link - Malerba, M. and R. Cerana, 2016. Chitosan effects on plant systems-A review. Int. J. Mol. Sci., Vol. 17, No. 7.
CrossRefDirect Link - Cretoiu, M.S., G.W. Korthals, J.H. Visser and J.D. van Elsas, 2013. Chitin amendment increases soil suppressiveness toward plant pathogens and modulates the actinobacterial and oxalobacteraceal communities in an experimental agricultural field. Applied Environ. Microbiol., 79: 5291-5301.
CrossRefDirect Link - Mulawarman, J. Hallmann, D. Bell, B. Kopp-Holtwiesche and R.A. Sikora, 2001. Effects of natural products on soil organisms and plant health enhancement. Meded. Rijksuniv. Gent. Fak. Landbouwkd. Toegep. Biol. Wet., 66: 609-617.
PubMedDirect Link - Pastucha, A., 2005. The effect of chitosan on the formation of microorganism communities in the rhizosphere soil of soybean. Acta Scientiarum Polonorum-Hortorum Cultus, 4: 69-77.
Direct Link - Postma, J., L.H. Stevens, G.L. Wiegers, E. Davelaar and E.H. Nijhuis, 2009. Biological control of Pythium aphanidermatum in cucumber with a combined application of Lysobacter enzymogenes strain 3.1T8 and chitosan. Biol. Control, 48: 301-309.
CrossRefDirect Link - Manjula, K. and A.R. Podile, 2001. Chitin-supplemented formulations improve biocontrol and plant growth promoting efficiency of Bacillus subtilis AF 1. Can. J. Microbiol. 47: 618-625.
CrossRefPubMedDirect Link - Lowe, A., S.M. Rafferty-McArdle and A.C. Cassells, 2012. Effects of AMF- and PGPR-root inoculation and a foliar chitosan spray in single and combined treatments on powdery mildew disease in strawberry. Agric. Food Sci., 21: 28-38.
Direct Link - Mishra, S., K.S. Jagadeesh, P.U. Krishnaraj and S. Prem, 2014. Biocontrol of tomato leaf curl virus (ToLCV) in tomato with chitosan supplemented formulations of Pseudomonas sp. under field conditions. Aust. J. Crop Sci., 8: 347-355.
Direct Link - Staehelin, C., M. Schultze, K. Tokuyasu, V. Poinsot, J.C. Prome, E. Kondorosi and A. Kondorosi, 2000. N-deacetylation of Sinorhizobium meliloti Nod factors increases their stability in the Medicago sativa rhizosphere and decreases their biological activity. Mol. Plant-Microbe Interact., 13: 72-79.
CrossRefDirect Link - Genre, A., M. Chabaud, C. Balzergue, V. Puech-Pages and M. Novero et al., 2013. Short-chain chitin oligomers from arbuscular mycorrhizal fungi trigger nuclear Ca2+ spiking in Medicago truncatula roots and their production is enhanced by strigolactone. New Phytol., 198: 190-202.
CrossRefDirect Link - Walker, T.S., H.P. Bais, E. Grotewold and J.M. Vivanco, 2003. Root exudation and rhizosphere biology. Plant Physiol., 132: 44-51.
CrossRefPubMedDirect Link - Bais, H.P., T.L. Weir, L.G. Perry, S. Gilroy and J.M. Vivanco, 2006. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol., 57: 233-266.
CrossRefPubMedDirect Link - Zeng, D. and D. Luo, 2012. Physiological effects of chitosan coating on wheat growth and activities of protective enzyme with drought tolerance. Open J. Soil Sci., 2: 282-288.
CrossRefDirect Link - Herde, O., H. Pena-Cortes, L. Willmitzer and J. Fisahn, 1997. Stomatal responses to jasmonic acid, linolenic acid and abscisic acid in wild-type and ABA-deficient tomato plants. Plant. Cell. Environ., 20: 136-141.
CrossRefDirect Link - Bittelli, M., M. Flury, G.S. Campbell and E.J. Nichols, 2001. Reduction of transpiration through foliar application of chitosan. Agric. Forest Meteorol., 107: 167-175.
CrossRefDirect Link - Pongprayoon, W., S. Roytrakul, R. Pichayangkura and S. Chadchawan, 2013. The role of hydrogen peroxide in chitosan-induced resistance to osmotic stress in rice (Oryza sativa L.). Plant Growth Regul., 70: 159-173.
CrossRefDirect Link - Je, J.Y. and S.K. Kim, 2006. Reactive oxygen species scavenging activity of aminoderivatized chitosan with different degree of deacetylation. Bioorg. Med. Chem., 14: 5989-5994.
CrossRefDirect Link - Rajalakshmi, A., N. Krithiga and A. Jayachitra, 2013. Antioxidant activity of the chitosan extracted from shrimp exoskeleton. Middle-East J. Scient. Res., 16: 1446-1451.
Direct Link - Kamari, A., I.D. Pulford and J.S.J. Hargreaves, 2011. Binding of heavy metal contaminants onto chitosans-An evaluation for remediation of metal contaminated soil and water. J. Environ. Manage., 92: 2675-2682.
CrossRefDirect Link - Sanchez-Duarte, R.G., D.I. Sanchez-Machado, J. Lopez-Cervantes and M.A. Correa-Murrieta, 2012. Adsorption of allura red dye by cross-linked chitosan from shrimp waste. Water Sci. Technol., 65: 618-623.
CrossRefDirect Link - Yong, S.K., M. Shrivastava, P. Srivastava, A. Kunhikrishnan and N. Bolan, 2015. Environmental applications of chitosan and its derivatives. Rev. Environ. Contam. Toxicol., 233: 1-43.
CrossRefDirect Link