
V. Sathya
Department of Biochemistry and Bioinformatics, Karpagam University, Coimbatore, Tamil Nadu, India
V.K. Gopalakrishnan
Department of Biochemistry and Bioinformatics, Karpagam University, Coimbatore, Tamil Nadu, India
American Journal of Drug Discovery and Development
Year: 2014 | Volume: 4 | Issue: 4 | Page No.: 232-240







ABSTRACT
Hepatocellular carcinoma ranks as the eighth most common cancer, sixth among men and eleventh among women and accounts for half a million deaths per year. Natural products, especially plants have been used for the treatment of various diseases for thousands of years which contain no side effects. The main aim and objective of the study is in vitro analysis of compounds such as epicatechin, myricetin, naringenin against Hep3B cell line and VERO cell line. In vitro assay of Hep3B cell line and VERO cell line was done by MTT assay. The assay is based on the reduction of soluble yellow tetrazolium salt to insoluble purple formation crystals by metabolically active cells. In vitro assay revealed that compounds such as epicatechin, myricetin and naringenin have efficacy in Hep3B cancer cell line and VERO normal cell line. By comparing the obtained results of MTT assay with these literature survey reports, it could be concluded that the compound has the potential to protect the cells from damage and the possible mechanism might be the inhibition of oxidative and inflammatory processes.
PDF Abstract XML References Citation
Received: September 13, 2014;
Accepted: September 22, 2014;
Published: December 27, 2014
How to cite this article
V. Sathya and V.K. Gopalakrishnan, 2014. In vitro Analysis of Phytochemicals Epicatechin, Naringenin, Myricetin
Against Hep3B Cell Lines and VERO Cell Lines. American Journal of Drug Discovery and Development, 4: 232-240.
DOI: 10.3923/ajdd.2014.232.240
URL: https://scialert.net/abstract/?doi=ajdd.2014.232.240
DOI: 10.3923/ajdd.2014.232.240
URL: https://scialert.net/abstract/?doi=ajdd.2014.232.240
INTRODUCTION
Cell viability is basically the determination of living cells within a cord blood sample. If the cell walls are generally intact and there is no damage to the cell structure, then the cell will be viable. Viability assay is an assay to determine the ability of cells or tissues to maintain or recover its viability. Viability assays are used to assess the success of cryopreservation techniques, the toxicity of substances or the effectiveness of substances in mitigating effects of toxic substances (Menendez et al., 2005).
The VERO cell line was derived from the kidney of a normal, adult, African green monkey (Cercopithecus) on March 27, 1962, by Y. Yasumura and Y. Kawakita at the Chiba University in Japan. The cell line was brought to the Laboratory of Tropical Vkology, NIAID and NIH at passage I level 93 from Chiba University by Dr. B. Simizu on June 15, 1964 (Murthy and Mathew, 2004). The cell line was submitted to the ATCC at passage level 113 and was propagated to passage level 121 at the ATCC to establish a bank of cells for availability. Initial characterization of VERO cells as a substrate for vaccine production was performed by the Aventis Pasteur (Mimeault et al., 2005). VERO cells have been extensively tested and shown to be free from adventitious agents for which there are currently available tests and to the limit of sensitivity of those tests. However, this cell line is an aneuploid cell line which does not form tumors at the passage levels used to produce vaccines but develops tumorogenic potential at higher passage levels (Szliszka et al., 2008).
Hep3B cells is a well-differentiated human hepatoma-derived cell line (Shukla and Gupta, 2010). The Hep3B cell lines provide an excellent in vitro system in which the physiological regulation of Epo expression is studied. The HBxAg stimulates the growth of Hep3B cells in serum free medium which is associated with elevated levels of alpha-catenin. A Hep3B cell was stably transfected with different truncation mutants of HBxAg (Tan et al., 2009). The Hep3B cells was also tested for susceptibility to the nucleoside 2'-C-methyladenosine known to be active against the HCV replicon in Huh7 cells and as a triphosphate against the HCV NS5B polymerase in vitro (Feng et al., 2001).
Epicatechin, a major polyphenolic component of green tea, is reported to have insulin mimetic action with protective effects on erythrocytes in a manner similar to insulin. (-) epicatechin also offers antioxidant protection against lipid peroxidation and inhibits platelet aggregation. (-) epicatechin effects would be beneficial in supporting the management of secondary complications of diabetes. Epicatechin is found in cocoa and may stimulate capillarity and mitochondrial proliferation in skeletal muscle (Young and Woodside, 2001). On the other hand, epigallocatechin and gallocatechin contain an additional phenolic hydroxyl group when compared to the former (Eisenbrand et al., 2002). Epicatechin reduces lipid peroxidation and inhibits platelet aggregation. Studies show that epicatechin and other flavonoids exert a protective role in osmotic fragility of cells, similar to that of insulin.
Myricetin is a naturally occurring flavonol, a flavonoid found in many grapes, berries, fruits, vegetables, herbs, as well as other plants. It is one of the phenolic compounds present in red wine. In vitro study suggests that myricetin in high concentrations can modify LDL cholesterol such that uptake by white blood cells is increased (Wakisaka et al., 2005). Myricetin have systemic effects on sensory neurons (Stefani et al., 1999).
Naringenin is a flavanone, a type of flavonoid, that is considered to have a bioactive effect on human health as antioxidant, free radical scavenger, anti-inflammatory, carbohydrate metabolism promoter and immune system modulator. The antiviral effects of naringenin are currently under clinical investigation. Naringenin lowers the plasma and hepatic cholesterol concentrations by suppressing HMG-CoA reductase and ACAT in rats fed a high-cholesterol diet. Scientists recently reported that naringenin could reduce tumor size, ROS levels and enhance antioxidant status. However, the mechanism of action of naringenin to suppress cell growth is still ambiguous since this compound appears to have multiple cellular targets including cytochrome P450 enzymes. The gastric cancer induced by MNNG which reportedly induce cellular oxidative damage through DNA strand breaks and explored the possible protective role of naringenin by measuring the levels of protein damage and polyamine synthesis in MNNG induced gastric carcinogenesis in experimental rats. Naringenin lowers the plasma and hepatic cholesterol concentrations by suppressing HMG-CoA reductase and ACAT in rats fed a high-cholesterol diet (Zhong et al., 2010).
The main objective of the present study is in vitro analysis of epicatechin, naringenin and myricetin against Hep3B cell line to study the anticancer activity of the above compounds and also to study the effect against the normal VERO cell line.
MATERIALS AND METHODS
Cell viability test-MTT assay (Mosmann, 1983): The viability of cells was assessed by MTT assay. The assay is based on the reduction of soluble yellow tetrazolium salt to insoluble purple formation crystals by metabolically active cells. The cells were plated separately in 96 well plates at a concentration of 1x105 cells well. After 24 h, cells were washed twice with 100 μL of serum free medium and starved for 1 h at 37°C. After starvation, cells were treated with different concentration of plant extract (500-15.25 μg mL-1) for 12, 24, 48, 72 h. At the end of treatment period, the medium was apirated and serum free medium containing MTT (0.5 mg mL-1) was added and incubated for 4 h at 37°C in a CO2 incubator.
The MTT containing medium was then discarded and the cells were washed with PBS (200 μL). The dark blue crystals were then dissolved by adding 10 μL of solubilising solution and mixed thoroughly. After a few minutes at room temperature to ensure that all crystals were dissolved, the plates were read on microplate reader at 570 nm, a reference wavelength of 630 nm and a calibration setting of 1.99 (or 1.00) if the samples were strongly coloured. Experiments of each test samples were carried out in triplicate including untreated cell control and a blank cell free control. The percentage cellular viability was calculated with the appropriate controls taken into account. The concentration which inhibited 50% of cellular growth (CTC50 value) was determined. The inhibitory rate of cell proliferation was calculated by the following equation:
The cytotoxic effect of sample on cancer cells were expressed as CTC50 values. All the experiments results were centered using three parallel measurements of the Mean±standard deviation.
RESULTS AND DISCUSSION
Compounds such as epicatechin, myricetin and naringenin were brought from Sigma Chemicals, Bangalore. The cytotoxicity study was carried out for compounds epicatechin, myricetin and naringenin. These compounds were screened for their cytotoxicity against Hep3B cell lines and VERO cell lines at different concentrations to determine the growth inhibition by and MTT assay. It was found that the growth inhibition (%) increasing with increasing concentration steadily up to 500 mg mL-1 on Hep3B cell line. Different compounds showed different cytotoxicity at different concentrations. Reports have shown that compounds are more active. The MTT assay is probably the most commonly used colorimetric indicator of cell viability and it has been used to evaluate cytotoxicity in a quantitative way in contrast with cell morphology evaluation by inverted light microscopy which is qualitative and more subjective. The reduction of MTT in the cell assesses the functional intactness of mitochondria on the basis of the enzymatic reduction of this tetrazolium salt by the mitochondrial dehydrogenases in viable cells. The MTT was first applied to quantify cellular proliferation and it is now widely used for screening antitumoral and antiviral activities of a large number of synthetic and natural products. This assay has several advantages; it is easy to perform, the evaluations are objective, it can be automated using a personal computer and the cytotoxicity measurement can be made in parallel with antiviral activity evaluation.
The present test result, when compared to the study conducted by Krishna and Jayakumaran (2010), showed supportive information that all compounds revealed more cytotoxic activity when compared to the control sample. The researchers investigated the cytotoxic potential of Tectona grandis extracts by MTT assay. In their study, the extract which contained flavonoids exhibited very high activity against Chick Embryo Fibroblast (CEF) and Human Embryonic Kidney (HEK 293) cells with 87 and 95.3% inhibition, respectively.
Similarly, investigation by Gaire et al. (2013) revealed that the Terminalia chebula fruit extract to protect neuronal cells against ischemia and inflammation in rat pheochromocytoma cells (PCl2) using in vitro oxygen-glucose deprivation followed by reoxygenation (OGD-R) ischemia and hydrogen peroxide (H2O2) induced cell death. Cell survival was evaluated by a 2-(4, 5-dimethylthiazol- 2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. They found that T. chebula extract increases the survival of cells subjected to OGD-R by 68% and H2O2 by 91.4%; reduces NO production and death rate of microglia cells stimulated by lipopolysaccharide (LPS).
By comparing the obtained results of MTT assay with these literature survey reports, it could be concluded that the compound has the potential to protect the cells from damage and the possible mechanism might be the inhibition of oxidative and inflammatory processes.
Assessment of cytotoxicity is clearly an important part of the evaluation of a potential antiviral agent because a useful compound should be selective for virus-specific processes with no or few effects on cellular metabolism and not show toxicity against the host. In vitro cytotoxicity evaluation is usually performed by using cell viability assays, such as the uptake of a dye by nonviable cells or by alterations in the mitochondrial function, these endpoints have been established for many years in many cell types.
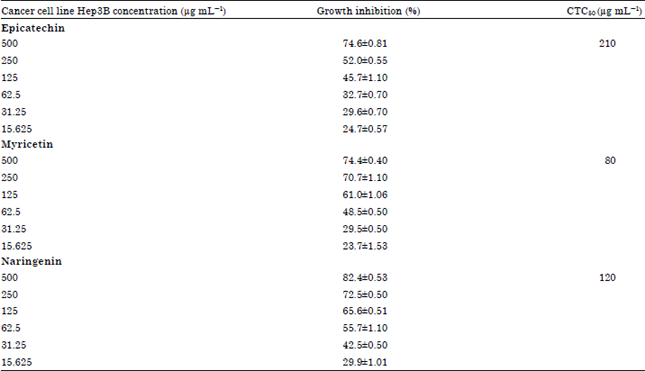
Table 1 shows the statistical analysis of cytotoxicity for the compounds epicatechin, myricetin, naringenin in Hep3B cell lines.
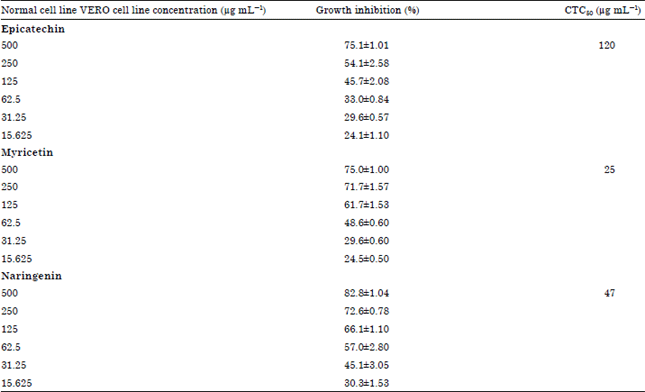
According to the results shown in Table 1, it was clear that the compound epicatechin gave the CTC50 value of 210 μg mL-1 which was the maximum value than myricetin (80 μg mL-1) and naringenin (120 μg mL-1) against Hep3B cell lines. In Table 2, VERO cell line epicatechin gave the CTC50 value of 125 μg mL-1 and in myricetin (25 μg mL-1) and naringenin (47 μg mL-1). Comparative analysis of CTC50 value of epicatechin, myricetin and naringenin revealed maximum growth inhibition of both VERO and Hep3B cell lines.
| Table 1: | Determination of cytotoxicity CTC50 for Hep3B cell lines |
 | |
| Table 2: | Determination of CTC50 for VERO cell lines |
 | |
Interestingly, maximum growth inhibition of the cancer cell line (Hep3B) was observed during the in vitro cytotoxicity analysis (Fig. 1 and 2).
Yamaguchi et al. (2002) observed that the compounds with free hydroxyls in their molecules showed toxicity to VERO cells. Robinetinidol (4R-6) gallocatechin (VI), which contains 10 free hydroxyls, was more cytotoxic than the other tested compounds. It was also verified that the compounds epicatechin (III), epigallocatechin (IV) and robinetinidol (4R-6) gallocatechin (VI) showed substantial effects against HSV-1 (29-R strain) at 107.14, 70.42 and 72.19 M, respectively. In relation to HSV-1 (KOS strain), the compounds IV and VI showed significant effects at 173.56 and 69.85 M, respectively. Epigallocatechin (IV) and robinetinidol (4R-6) gallocatechin (VI) demonstrated promising antiherpetic activity against both tested against HSV-1 (29-R strain).
Figure 3 shows the comparative analysis of the phytochemicals epicatechin, myricetin, naringenin against Hep3B cancer cell lines and VERO normal cell lines.
Bulus et al. (2011) showed a selective cytotoxicity and apoptosis inductive effect indicative of an anticancer activity of the 50% ethanol-water leaf extract of Polyalthia evecta on the human hepatoma (HepG2) cell line.
Panunto et al. (2011) showed that naringenin did not obviously inhibit tumor proliferation either in vitro or in vivo. These results suggest that naringenin does not kill tumor cells directly.
Liyanage et al. (2002) examined the effect of naringenin on immunosuppression related molecules and cells in vitro. The results revealed that naringenin significantly inhibited the production of TGF-β1 and IL-10. Furthermore, naringenin inhibited TGF-β1 induced Treg production in vitro.
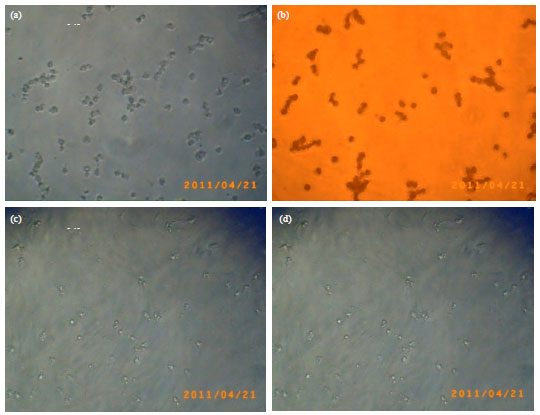 |
| Fig. 1: | Cytotoxicity effect of the compounds (a) Control, (b) Epicatechin, (c) Myricetin and (d) Naringenin against Hep3B cancer cell lines |
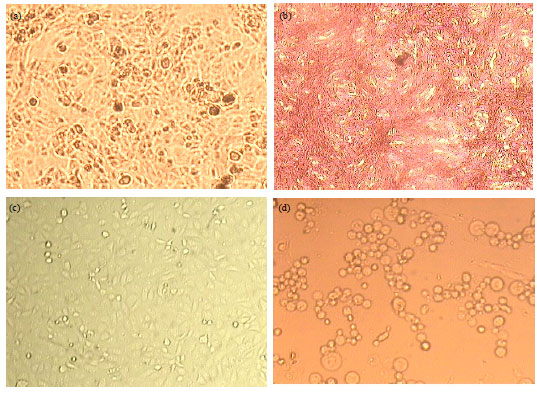 |
| Fig. 2: | Potential effect of the compounds (a) Control, (b) Epicatechin, (c) Myricetin and (d) Naringenin against VERO normal cell line |
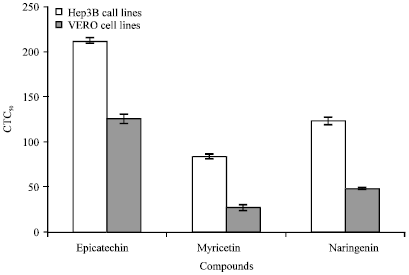 |
| Fig. 3: | Comparative analysis of CTC50 for Hep3B and VERO cell lines |
This data demonstrate that naringenin can reverse Treg related immunosuppression which in turn, increase the proportion of activated T cells.
Beyer and Schultze (2006) suggested that orally administered naringenin can inhibit the outgrowth of metastases after surgery via regulating host immunity. The antimetastatic properties and low toxicity of naringenin make it an ideal surgical adjuvant therapy for breast cancer.
Shenkier et al. (2004) stated that catechin and epicatechin that were isolated from Eysenhardtia subcoriacea demonstrated strong radical scavenging properties against diphenylpicrylhydrazil (DPPH). Similarly, Ko et al. (2005) reported that epigallocatechin-gallocatechin and epicatechin-Catechin inhibited DPPH by more than 95%.
Pulaski et al. (2000) concluded that naringenin has also been shown to reduce hepatitis C virus production by infected hepatocytes (liver cells) in cell culture. This seems to be secondary to naringenin’s ability to inhibit the secretion of very-low-density lipoprotein by the cells.
CONCLUSION
During the last decades, a global trend is again turned towards naturopathy and now days, green pharmaceuticals are matter of high concern due to minimum side effects and being cost effective. The present study showed that the compounds epicatechin, myricetin and naringenin had a high potential against cancer cells and less activity against normal cell lines. Thus, these compounds provide strong motivators for the development of the novel therapeutic drug against hepatocellular carcinoma.
REFERENCES
- Tan, J., B. Wang and L. Zhu, 2009. Regulation of survivin and Bcl-2 in HepG2 cell apoptosis induced by quercetin. Chem. Biodivers., 6: 1101-1110.
CrossRefDirect Link - Bulus, T., S.E. Atawodi and M. Mamman, 2011. Acute toxicity effect of the aqueous extract of Terminalia avicennioides on white albino rats. Sci. World J., 6: 1-4.
Direct Link - Stefani, E.D., P. Boffetta, H. Deneo-Pellegrini, M. Mendilaharsu, J.C. Carzoglio, A. Ronco and L. Olivera, 1999. Dietary antioxidants and lung cancer risk: A case-control study in uruguay. Nutr. Cancer, 34: 100-110.
CrossRefPubMedDirect Link - Feng, Q., T. Kumagai, Y. Torii, Y. Nakamura, T. Osawa and K. Uchida, 2001. Anticarcinogenic antioxidants as inhibitors against intracellular oxidative stress. Free Radical Res., 35: 779-788.
CrossRefPubMedDirect Link - Gaire, B.P., N. Jamarkattel-Pandit, D. Lee, J. Song and J.Y. Kim et al., 2013. Terminalia chebula extract protects OGD-R induced PC12 cell death and inhibits LPS induced microglia activation. Molecules, 18: 3529-3542.
CrossRefDirect Link - Ko, K., S. Yamazaki, K. Nakamura, T. Nishioka and K. Hirota et al., 2005. Treatment of advanced tumors with agonistic anti-GITR mAb and its effects on tumor-infiltrating Foxp3+CD25+CD4+ regulatory T cells. J. Exp. Med., 202: 885-891.
PubMedDirect Link - Krishna, M.S. and N.A. Jayakumaran, 2010. Antibacterial, cytotoxic and antioxidant potential of different extracts from leaf, bark and wood of Tectona grandis. Int. J. Pharm. Sci. Drug Res., 2: 155-158.
Direct Link - Zhong, Y., C. Krisanapun, S.H. Lee, T. Nualsanit, C. Sams, P. Peungvicha and S.J. Baek, 2010. Molecular targets of apigenin in colorectal cancer cells: Involvement of p21, NAG-1 and p53. Eur. J. Cancer, 46: 3365-3374.
CrossRefDirect Link - Liyanage, U.K., T.T. Moore, H.G. Joo, Y. Tanaka and V. Herrmann et al., 2002. Prevalence of regulatory T cells is increased in peripheral blood and tumor microenvironment of patients with pancreas or breast adenocarcinoma. J. Immunol., 169: 2756-2761.
CrossRefPubMedDirect Link - Menendez, J.A., L. Vellon and R. Lupu, 2005. Antitumoral actions of the anti-obesity drug orlistat (XenicalTM) in breast cancer cells: Blockade of cell cycle progression, promotion of apoptotic cell death and PEA3-mediated transcriptional repression of Her2/neu (erbB-2) oncogene. Ann. Oncol., 16: 1253-1267.
PubMedDirect Link - Mimeault, M., N. Jouy, P. Depreux and J.P. Henichart, 2005. Synergistic antiproliferative and apoptotic effects induced by mixed epidermal growth factor receptor inhibitor ZD1839 and nitric oxide donor in human prostatic cancer cell lines. Prostate, 62: 187-199.
CrossRefDirect Link - Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods, 65: 55-63.
CrossRefPubMedDirect Link - Murthy, N.S. and A. Mathew, 2004. Cancer epidemiology, prevention and control. Curr. Sci., 86: 518-527.
Direct Link - Panunto, W., K. Jaijoy, N. Lerdvuthisopon, N. Lertprasertsuke, N. Jiruntanat, N. Soonthornchareonnon and S. Sireeratawong, 2011. Acute and chronic toxicity studies of the water extract from dried fruits of Terminalia chebula Rezt. in rats. Int. J. Applied Res. Nat. Prod., 3: 36-43.
Direct Link - Pulaski, B.A., D.S. Terman, S. Khan, E. Muller and S. Ostrand-Rosenberg, 2000. Cooperativity of Staphylococcal aureus enterotoxin B superantigen, major histocompatibility complex class II and CD80 for immunotherapy of advanced spontaneous metastases in a clinically relevant postoperative mouse breast cancer model. Cancer Res., 60: 2710-2711.
PubMedDirect Link - Shenkier, T., L. Weir, M. Levine, I. Olivotto, T. Whelan and L. Reyno, 2004. Clinical practice guidelines for the care and treatment of breast cancer: 15. Treatment for women with stage III or locally advanced breast cancer. Can. Med. Assoc. J., 170: 983-994.
CrossRefPubMedDirect Link - Shukla, S. and S. Gupta, 2010. Apigenin: A promising molecule for cancer prevention. Pharmaceut. Res., 27: 962-978.
CrossRefDirect Link - Szliszka, E., Z.P. Czuba, K. Jernas and W. Krol, 2008. Dietary flavonoids sensitize HeLa cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). Int. J. Mol. Sci., 9: 56-64.
CrossRefDirect Link - Wakisaka, N., T. Yoshizaki, N. Raab-Traub and J. Pagano, 2005. Ribonucleotide reductase inhibitors enhance cidofovir-induced apoptosis in EBV-positive nasopharyngeal carcinoma xenografts. Int. J. Cancer, 116: 640-645.
CrossRefDirect Link - Yamaguchi, K., M. Honda, H. Ikigai, Y. Hara and T. Shimamura, 2002. Inhibitory effects of (-)-epigallocatechin gallate on the life cycle of human immunodeficiency virus type 1 (HIV-1). Antiviral Res., 53: 19-34.
CrossRefDirect Link - Young, I.S. and J.V. Woodside, 2001. Antioxidants in health and disease. J. Clin. Pathol., 54: 176-186.
CrossRefPubMedDirect Link