
M. Ikhwanuddin
Institute of Tropical Aquaculture, Universiti Malaysia Terengganu, 21030 Kuala Terengganu, Terengganu, Malaysia
S.N. Fatihah
Institute of Tropical Aquaculture, Universiti Malaysia Terengganu, 21030 Kuala Terengganu, Terengganu, Malaysia
A.H. Nurfaseha
Institute of Tropical Aquaculture, Universiti Malaysia Terengganu, 21030 Kuala Terengganu, Terengganu, Malaysia
M. Fathiah
School of Marine Sciences and Environment, Universiti Malaysia Terengganu, 21030 Kuala Terengganu, Terengganu, Malaysia
M. Effenddy
School of Marine Sciences and Environment, Universiti Malaysia Terengganu, 21030 Kuala Terengganu, Terengganu, Malaysia
A. Shamsudin
School of Marine Sciences and Environment, Universiti Malaysia Terengganu, 21030 Kuala Terengganu, Terengganu, Malaysia
A. Siti Aishah
School of Marine Sciences and Environment, Universiti Malaysia Terengganu, 21030 Kuala Terengganu, Terengganu, Malaysia
A.B. Abol-Munafi
School of Fisheries and Aquaculture Sciences, Universiti Malaysia Terengganu, 21030 Kuala Terengganu, Terengganu, Malaysia
Asian Journal of Cell Biology
Year: 2014 | Volume: 9 | Issue: 1 | Page No.: 1-13







ABSTRACT
A study was carried out to determine the ovarian maturation stages of 100 days and the effect on the embryonic development of mud spiny lobster, Panulirus polyphagus. For the ovarian development, treatments are involving water temperature (low temperature at 15-20°C and high temperature at 30-35°C) and control group at ambient condition of temperature at 26-29°C. Results showed that there were no significant difference (p>0.05) in all treatments in terms Gonadosomatic Index (GSI) and Oocyte Stage Index (OSI) however, there were significant difference (p<0.05) at oocyte diameters for all treatments. While, for the embryonic development, five berried females kept at temperature 25-28°C (control), 20°C (low temperature) and 30°C (high temperature). In control, the largest mean diameters were 0.668±0.037 mm (Day 11). However, in low temperature (R1 and R2), the largest mean diameters were 0.632±0.039 mm (Day 5) and 0.598±0.041 mm (Day 17) and in high temperature (R1 and R2), the largest mean diameter were 0.651±0.012 mm (Day 4) and 0.679±0.022 mm (Day 3). Therefore, ambient condition was found to be suitable to stimulate ovarian development as P. polyphagus could definitely adapt to the natural environment compared to extreme conditions and culturing the berried female in low temperature could cause mortality to the lobster and trigger the berried female to release the eggs unhatch due to stress.
PDF Abstract XML References Citation
Received: January 17, 2014;
Accepted: March 15, 2014;
Published: June 26, 2014
How to cite this article
M. Ikhwanuddin, S.N. Fatihah, A.H. Nurfaseha, M. Fathiah, M. Effenddy, A. Shamsudin, A. Siti Aishah and A.B. Abol-Munafi, 2014. Effect of Temperature on Ovarian Maturation Stages and Embryonic Development of Mud Spiny Lobster, Panulirus polyphagus. Asian Journal of Cell Biology, 9: 1-13.
DOI: 10.3923/ajcb.2014.1.13
URL: https://scialert.net/abstract/?doi=ajcb.2014.1.13
DOI: 10.3923/ajcb.2014.1.13
URL: https://scialert.net/abstract/?doi=ajcb.2014.1.13
INTRODUCTION
Spiny lobsters are captured and exploited in over 90 countries (Philips and Kittaka, 2000). Most fisheries are fully exploited or overexploited and one of the few ways to enlarge production is all the way through aquaculture. In the commercial culture of spiny lobsters, the most hurdles are difficulty in growing species throughout all their larval stages (Kittaka and Booth, 2000). Kittaka (1994) state that temperature affects phyllosoma growth and various species of spiny lobster require different temperature stages. Besides, reproductive cycle of crustacean species can be controlled by temperature and photoperiod since those are important environmental factors enhancing growth of organisms (Meusy and Payen, 1988).
Among the key constraints in understanding of fisheries management and development of mud spiny lobsters, Panulirus polyphagus is that paucity of information for fishery management purposes. Due to the increase in demand of this species in the market now-a-days, sufficient data on the distribution and reproduction is needed to fulfill the requirements of P. polyphagus. Studies on the effect of certain parameters on the ovarian maturation stages and early embryonic development are important as to widen the information on this species and can serve as the baseline which could be useful in the field of aquaculture and fishery management. The data analyzed in the ovarian maturation stages and early embryonic development in a given condition can be used to determine the optimum condition to shorten the the ovarian maturation stages and egg incubation period in the breeding technology of P. polyphagus. The main objectives of the present study are to determine the effects of temperature on the ovarian development of mud spiny lobster, P. polyphagus which is described based on the external and histological characteristics of the ovary and to determine different temperature regimes (15, 25 and 35°C) on the embryonic development and on egg incubation period of P. polyphagus.
MATERIALS AND METHODS
Effect of temperature on ovarian maturation stages of mud spiny lobster, Panulirus polyphagus: Female broodstocks of P. polyphagus with Carapace Length (CL) less than 65.9 mm were sourced from the fishermen from Sungai Rengit, Johor, Malaysia. Lobsters were individually weighed and measured for CL. The CL was measured from rear of the eye socket parallel to the centre line of the body shell to the rear of the body shell. Then, lobsters were transferred into 500 L fibre glass tank filled with seawater before being transferred into respective treatment tanks. Three broodstocks were sacrificed on the 1st day and the maturation stage of samples from wild environment was determined through histological assessment.
Another 3 broodstocks were introduced to each treatment tank of different temperature regime of control (T1), low temperature regime of 20-25°C (T2) and high temperature regime of 30-35°C (T3). Both treatment groups were sharing the same control sample organisms reared at ambient temperature of 26-29°C. Experiment was conducted within 100 days and broodstocks were fed twice daily with chunked squid at 100% biomass. Culture tanks were daily siphoned to remove any uneaten feed and faecal matters. The optimum conditions in culturing P. polyphagus are temperature based on preferred treatments, dissolved oxygen at 5.0-8.0 mg mL-1 pH 6.5-8.5 and ammonia level of 0.025 mg mL-1. After 100 days, P. polyphagus were measured, weighed and dissected to obtain the ovaries which to be used in order to determine the gonad colouration and development stages, Gonadosomatic Index (GSI) and Oocyte Size Index (OSI). Gonadosomatic Index (GSI) and Oocyte Size Index (OSI) were determined based on the following equation by Minagawa (1997):
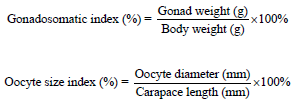 |
After weighing for GSI, ovaries were quickly fixed in Bouin’s solution for 24 h. Afterwards, gonads were dehydrated through an alcohol series. Purpose of embedding was to provide an easy handled solid block held in microtome. Cutting was done using rotary microtome at 4-7 μm thickness. Then, floating inside the water bath at 38-40°C was done to stretch the ribbon and placed on top of clean glass slide after ‘fishing’. Glass slides holding stretched ribbons were dried on hot plate at 60°C overnight. Haematoxylin and eosin method were used for staining purpose.
Diameters of 100 oocytes were randomly measured from the triplicates of each treatment using image analyzer (Nikon Eclipse 80i). All statistics were performed using SPSS version 20.0 for Windows. The Gonadosomatic Index (GSI), mean of oocyte diameter and Oocyte Size Index (OSI) were analyzed using One Way Analysis of Variance (ANOVA). The Independent t-test was used for further analysis of significant between treatments.
Effect of temperature on the embryonic development of mud spiny lobster, Panulirus polyphagus: Five berried females were kept at temperature 25-28°C (one berried female for control), 20°C (two berried females for low temperature, replicate 1 (R1) and replicate 2 (R2) and 30°C (two berried females for high temperature, replicate 1 (R1) and replicate 2 (R2). The berried female was fed with squid at 20% body weight. Every day, the eggs were recorded to identify the respective embryonic stages. Three tanks (5 tonnes capacity) were prepared to culture the broodstock; control tank, low temperature treatment tank and high temperature treatment tank. For low temperature, water chiller was used to maintain the temperature at 20°C. Meanwhile, for high temperature at 30°C was maintained using electric heater (30 W) with regulator. About 30 eggs were taken daily from the berried female by scratching the egg bundle gently with water dropper. Eggs samples from each water temperature berried female were kept in bottles and fixed with 5% formalin. Eggs for development were observed under Nikon Measuring Microscope MM-800. The diameter of egg was recorded by using data processing software E-MAX V5.20 while the development of the embryo was recorded as images by using NIS-Elements F2.30 SP3 imaging software and identified according to Tong et al. (2000). Each change in the stage of embryonic development such as changes in egg colour, presence of eye spot and chromatophore was identified.
RESULTS AND DISCUSSION
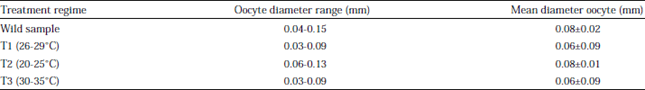
Effect of temperature on ovarian maturation stages of mud spiny lobster, Panulirus polyphagus: Ovarian maturation stages of P. polyphagus reared within 100 days at different temperatures treatments were classified based on the colouration, characteristic of the ovary and the oocyte diameters (Table 1, Fig. 1, 2). Results showed that there were no significant difference (p>0.05) noted between the treatment of GSI and OSI. As for temperature, T2 showed the highest value as compared to others in the same group in terms of oocyte diameter, mean CL, mean BW, GSI and OSI which were 0.08±0.01 mm, 59.83±5.81 mm, 202.62±35.86 g, 0.39±0.12% and 0.16±0.08%, respectively (Fig. 3-5).
| Table 1: | Ovarian maturation of Panulirus polyphagus reared within 100 days at different temperatures |
 | |
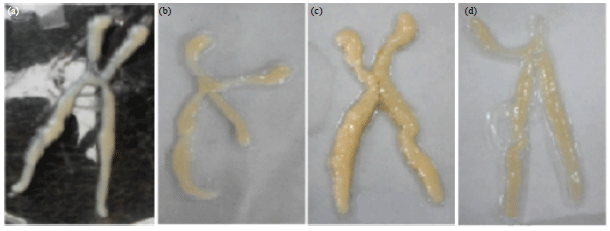 | |
| Fig. 1(a-d): | Morphology of ovaries (H-shape) of Panulirus polyphagus in different water temperature treatments reared within 100 days, (a) Wild sample, (b) Control of 26-28°C (T1), (c) Low temperature regime of 20-25°C (T2) and (d) High temperature regime of 30-35°C (T3) |
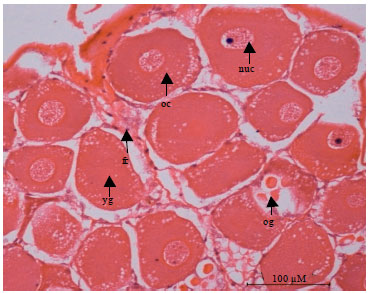 | |
| Fig. 2: | Histology of immature or non-vitellogenic ovary (stage 1) of mud spiny lobster, Panulirus polyphagus. The section oocyte (oc), nucleus (nc) and oogonia (og) in each lobe. Follicle cells (fc) around the oocytes and yolk globules (yg) on large oocytes |
Based on histological study, the ovaries which classified as the developing or latter stages in maturation were categorized as physiologically mature. In immature or non-vitellogenic phase, the ovary is transparent to translucent tissue that sometimes difficult to recognize. Histological sections showed basophilic oogonia and primary oocyte with large nuclei. The follicle cells are small and distributed around the periphery of the lobes and in an area among groups of oogonia and oocytes. The follicle cells gradually enclosed the oocyte. The nucleus of oocyte commonly contains nucleolus as well as dispersed chromatin.
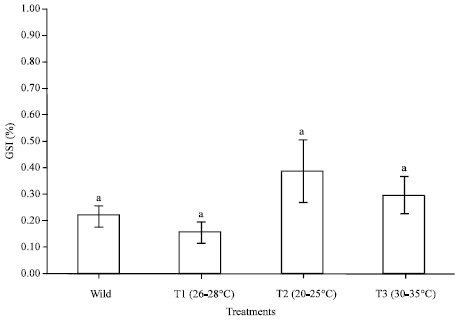 | |
| Fig. 3: | Mean of gonadosomatic index (GSI) in different water temperature treatments reared within 100 days of Panulirus polyphagus. Bars with the same letters were not significantly different (p>0.05) |
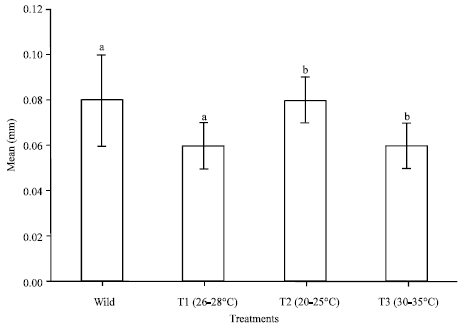 | |
| Fig. 4: | Mean oocyte diameter of different water temperature reared within 100 days of Panulirus polyphagus, bars with same letters are significantly different (p<0.05) |
Water temperature had a significant effect on the permissive stimulation of ovarian maturation by long day photoperiod in captive of American lobster, Homarus americanus (Aiken and Waddy, 1980; Quackenbush and Herrnkind, 1983). Endocrine control of ovarian maturation and the endocrine control of the moult cycle of somatic growth were integrated in response to changes in photoperiod and temperature (Quackenbush and Herrnkind, 1983). Temperature and photoperiod play an important role in regulating ovarian development and oviposition which induced under long photoperiod and high temperature conditions (Matsuda et al., 2002; Hamasaki et al., 2004).
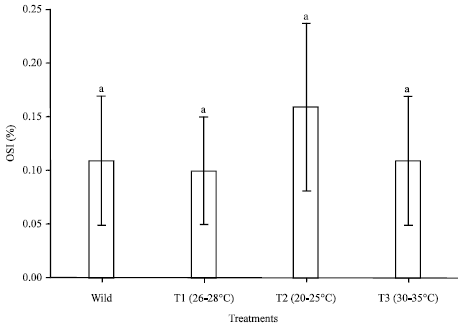 | |
| Fig. 5: | Mean Oocyte Size Index (OSI) of different water temperature treatment reared within 100 days of Panulirus polyphagus, bars with same letter are not significantly different (p>0.05) |
The particular combination of photoperiod and temperature inducing ovarian development varies with species or population because these factors depending on the natural environment and season of breeding (Matsuda et al., 2002).
Panulirus polyphagus usually found at muddy substrate, subtidal rock and coral reef area with temperature range of 20-29°C (UNEP, 2004) as this comply to the optimal temperature (20-25°C) used to achieve better ovarian growth as in present study. Other studies on Japanese spiny lobster, Panulirus japonicus showed that prevention of ovarian development at higher temperature under short photoperiod obviously demonstrates that increasing in water temperature alone did not affect ovarian development. Culture conditions at high temperature and short photoperiod which prevented ovarian development seem to activate moulting P. japonicus. A short photoperiod and high temperature are possible more efficient conditions for continuing growth of the lobsters due to high moulting rate and consequent rapid growth (Matsuda et al., 2002).
Ovarian development corresponded well in changes in GSI and ovigerous percentage (Chang et al., 2007). Colour changes with ovaries maturation and ovarian development stages (Islam et al., 2010). The cells that constitute the ovaries are of 3 main types: Oogonia, oocytes in different stages of development and follicle cells (Santos et al., 2009). Ovarian maturation cycle is differentiated into 7 stages which are immature, early maturing I, early maturing II, late maturing, mature, semispent and spent (Kagwade, 1988). Through microscopic examination, there is only 1 oogenesis stage which was at immature stage. At normal condition, the diameter of oocytes increased as the ovarian developmental stage progressed. According to Chang et al. (2007), a histological analysis of the developmental stages of oocyte is the most accurate method for determining sexual maturity because unberried but mature females are indistinguishable from external morphology.
Results of histological allowed affirming that the ovaries vary according to gonadal development stages (Santos et al., 2009).
 | |
| Fig. 6(a-c): | Change of egg colour at the pleopods of the Panulirus polyphagus broodstock (a) Early stage-bright orange, (b) Middle stage-brick red and (c) Prior to hatching-dark red |
| Table 2: | Chronology of embryonic development stages of Panulirus polyphagus at ambient temperature of control treatment |
 | |
Quinitio et al. (2007) stated that developing oocytes and the follicle cells or nurse cells are the major cell types found within the ovarian lobes of crustacean where it is agreed in this study of Panulirus polyphagus where the follicle cells are larger and more obvious in immature gonads. The colour of immature ovaries is translucent to yellow and becomes darker yellow to dark orange in mature ovaries due to accumulation of yolk in the oocytes. The yellow colour of the lipoprotein is due to β-carotene, astaxanthin and other unidentified carotenoids (Quinitio et al., 2007). According to Philips et al. (2000), in immature condition, the ovary is externally white or weakly yellow. The colour, size and texture of the ovary are closely related to its cellular development. The presence of immature and recovery stage ovaries were observed year round (Chang et al., 2007).
Effect of temperature on the embryonic development of mud spiny lobster, Panulirus polyphagus: The eggs appeared to be bright orange in the earlier stage, turning to brick red in colour and lastly dark red in colour prior to hatching (Fig. 6). The colour change of the eggs was same for all samples in every temperature treatments. The development of the embryo was shown in Table 2 and Fig. 7.
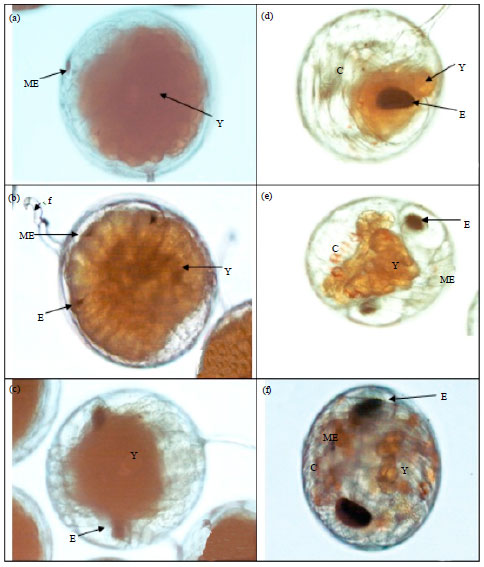 | |
| Fig. 7(a-f): | Embryonic development stages of Panulirus polyphagus berried female (a) Stage 1, (b) Stage 2, (c) Stage 3, (d) Stage 4, (e) Stage 5 and (f) Stage 6, ME: Median eye, Y: Egg yolk, f: Funiculus, E: Eyespot, C: Chromatophore |
Table 3 shows the time taken to reach next stage from the previous stage of the embryo. Only eggs from berried female in control treatment and high temperature treatment; (R1) that managed was observed from stage 1 until 6. Both berried female for control and high temperature (R1) incubate their eggs for about 264 h. In addition, 24 h was taken by the embryo from stage 6 to hatch. For berried female in high temperature (R2), only stage 5, 6 and time taken to hatch was obtained. The time taken from stage 5-6 and stage 6 to hatch was same for both replicate in high temperature which 72 and 24 h. For low temperature, only stage 4 and 5 embryo managed was observed in R1 while only stage 2 embryos managed was observed in R2. Berried female mortality was observed in berried female of R1 after 360 h incubating the eggs in low temperature. For berried female of R2 has released all her eggs unhatched after 480 h incubating the eggs.
In control, the R2 value was 0.992, showing that temperature at ambient level was really effective towards the development of egg diameter (Fig. 8a). The largest mean diameter recorded was on Day 11 (0.668±0.037 mm).
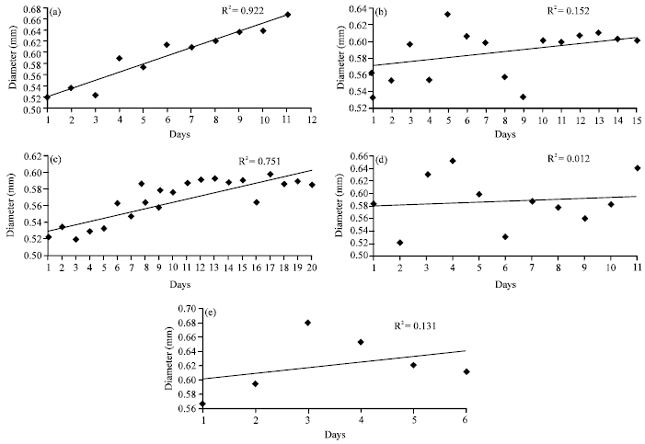 | |
| Fig. 8(a-e): | Change in mean diameter of eggs during embryonic development of Panulirus polyphagus for (a) Control, (b) Low temperature treatment of replicate 1, (c) Low temperature treatment of replicate 2, (d) High temperature treatment of replicate 1 and (e) High temperature treatment of replicate 2 |
| Table 3: | Time taken (hour) to reach next stage from previous stage of embryo development stages of Panulirus polyphagus. Low temperature and high temperature treatment have two replicates each of replicate 1 (R1) and replicate 2 (R2) |
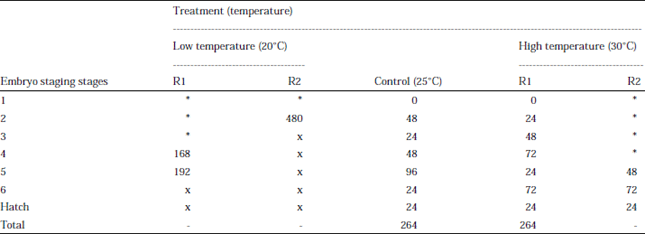 | |
| *Berried female when obtained already had embryo at later stages of development, x: Embryo either died or berried female released the eggs unhatched, -: No data can be calculated as the development was not observed from early stage | |
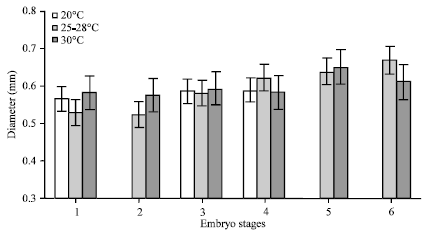 | |
| Fig. 9: | Change in mean egg diameter according to different embryonic development stages of Panulirus polyphagus for all treatments. Control (25-28°C, low temperature (20°C) and high temperature (30°C) |
The smallest mean diameter was recorded on Day 1 (0.520±0.023). In low temperature, the R2 value was 0.152 for R1 and 0.751 for R2 (Fig. 8a, b). The largest mean diameter recorded was on day 5 (0.632±0.039 mm) and the smallest mean diameter recorded was 0.535±0.042 mm. For low temperature of R2, increase in diameter throughout the incubation period was observed (R2 = 0.751) (Fig. 8b). The largest mean diameter observed was 0.598±0.041 mm on Day 17 and the smallest mean diameter was 0.520±0.028 mm on Day 3. In high temperature, the R2 value was 0.012 for R1 and 0.131 for R2 (Fig. 8d, e). The largest mean diameter recorded was 0.651±0.012 mm on Day 4 of observation and the smallest mean diameter was 0.522±0.018 mm on day 2 for high temperature of R1. For R2 of high temperature, the increase of the diameter was slow and the diameter was decreased from Day 3 until the end of incubation with the largest diameter recorded is 0.679±0.022 mm on Day 3 while the smallest mean diameter is 0.567±0.042 mm on the Day 1 (Fig. 8e). Figure 9 shows the change in egg diameter according to different stages for all treatments with growth of the embryo was observed throughout the incubation period.
In this study, the colour of the egg bundle at the pleopods of the berried female at early stage was bright orange, turning to brick red in colour and lastly dark red in colour few days prior to hatching. It was because of the absorption of the yellow yolk and development of dark pigment in the eyes and appearance of chromatophores on the abdomen (Samuel and Soundarapandian, 2010). Prescott (1988) explained that in the Solomon Islands, where average seawater temperature was 28-29°C, incubation period was for a period of approximately 3 weeks for P. penicillatus. In higher latitudes where water temperatures are reduced, incubation takes longer period. Adult female P. ornatus has an incubation period of approximately 3 weeks before the developed larvae hatch when held at ambient summer temperatures (Jones et al., 2003). In present study, Panulirus polyphagus took 264 h (11days) to incubate and hatch. This data was gained from berried female in control treatment and high temperature treatment of Replicate 1. The differences in the period of incubation for Panulirus polyphagus and lobsters in previous studies could be due to the original habitat and regional differences. In marine lobsters, the incubation period between egg fertilization and egg hatch was temperature dependant (Tong et al., 2000). Incubation period was negatively correlated with temperature and can vary widely, for example, the incubation period of Eastern Rock Lobster, Sagmariasus verreauxi can range between 55 days (20°C) and 130 days (13°C) depending on the incubation temperature (Samuel and Soundarapandian, 2010). Previously study have shown that the development rate of the embryos and the egg incubation time of the spiny lobster J. edwardsii is directly related to water temperature and this has allowed the development of a useful quantitative staging technique, based on an eye index, that can accurately predict hatching time (Tong et al., 2000).
In present study, only egg bundle from broodstock cultured in control (25-28°C) and high temperature (30°C) condition managed to hatch. Mortality and the behaviour of releasing egg unhatch from the pleopods is observed for berried female cultured in low temperature (20°C). This shows that Panulirus polyphagus from this region prefer higher temperature for the development of the embryo stage. The present study was started with berried female that already have later stage, which cause some of data was insufficient from earlier stage or until hatching moments. The time taken to reach next stage from previous stage of the present study was compared by stage that synchronizes which stage 4 and 5. To reach stage 4 from stage 3, longer time was taken for the embryo to develop in low temperature (20°C) with 168 h, followed by high temperature (30°C) with 72 h and control (25-28°C) with only 48 h. To reach stage 5 from stage 4, low temperature still took the longest time (192 h) compared to the other temperature treatment at 96 h for control, 24 h for high temperature of R1 and 48 h for high temperature of R2.
One of the most interesting features of lobster embryonic development stages occurs when 80% of embryonic development time has elapsed. However, if an embryo reaches 80% development and the temperature was not high enough, they cease to develop and remain in stasis at the 80% development plateau until the ambient temperature increases to a suitable level (Helluy and Beltz, 1991). For some of the replicates (R2 of low temperature and R1 of the high temperature), the mean diameter of the eggs was not uniformly increased throughout the incubation period and abruptly increased and decreased because the egg in the bundle actually did not develop at the same rate. However, most of the replicates, increase in diameter was observed throughout the incubation period which shows that the embryo did undergo growth in size.
The colour of the egg appeared to be bright orange in the earlier stage, turning to brick red in colour and lastly dark red in colour few days prior to hatching. The developing embryo was defined in this study based on daily progress. The time taken for the incubation period only gained from broodstock in control and high temperature (R1) which was 264 h (11 days). Culturing the berried female in low temperature could cause mortality to the lobster and trigger the berried female to release the eggs unhatch due to stress. There was increase in the size of the embryo throughout the incubation period but some are not uniform because of the rate of development of the embryo in the same egg bundle is different. From this study, the optimum temperature that can promote healthy growth of the embryo during incubation period was 25-30°C.
ACKNOWLEDGMENTS
This study had been funded by Malaysia’s Ministry of Higher Education under Exploratory Research Grant Scheme (Vot. No. 55051). Our great appreciation to all staffs of Institute of Tropical Aquaculture, Universiti Malaysia Terengganu who’s involved directly or indirectly during this study.
REFERENCES
- Chang, Y.J., C.L. Sun, C. Yong, S.Z. Yeh and W.C. Chiang, 2007. Reproductive biology of the spiny lobster, Panulirus penicillatus, in the southeastern coastal waters off Taiwan. Mar. Biol., 151: 553-564.
CrossRefDirect Link - Hamasaki, K., H. Imai, N. Akiyama and K. Fukunaga, 2004. Ovarian development and induced oviposition of the overwintering swimming crab Portunus trituberculatus (Brachyura: Portunidae) reared in the laboratory. Fish. Sci., 70: 988-995.
CrossRefDirect Link - Helluy, S.M. and B.S. Beltz, 1991. Embryonic development of the American lobster (Homarus americanus): Quantitative staging and characterization of an embryonic molt cycle. Biol. Bull., 180: 355-371.
Direct Link - Islam, M.S., K. Kodama and H. Kurokura, 2010. Ovarian development of the mud crab Scylla paramamosain in a tropical mangrove swamps, Thailand. J. Sci. Res., 2: 380-389.
Direct Link - Kagwade, P.V., 1988. Reproduction in the spiny lobster Panulirus polyphagus (Herbst). J. Mar. Biol. Assoc. India, 30: 37-46.
Direct Link - Kittaka, J., 1994. Culture of phyllosomas of spiny lobster and its application to studies of larval recruitment and aquaculture. Crustaceana, 66: 258-270.
Direct Link - Tong, L.J., G.A. Moss, T.D. Pickering and M.P. Paewai, 2000. Temperature effects on embryo and early larval development of the spiny lobster Jasus edwardsii and description of a method to predict larval hatch times. Mar. Freshwater Res., 51: 243-248.
Direct Link - Matsuda, H., T. Takenouchi and T. Yamakawa, 2002. Effects of photoperiod and temperature on ovarian development and spawning of the Japanese spiny lobster Panulirus japonicus. Aquaculture, 205: 385-398.
CrossRef - Meusy, J.J. and G.G. Payen, 1988. Review: Female reproduction in Malacostracan crustacea. Zool. Sci., 5: 217-265.
Direct Link - Minagawa, M., 1997. Reproductive cycle and size-dependent spawning of female spiny lobsters (Panulirus japonicas) off Oshima Island, Tokyo, Japan. Mar. Freshwater Res., 48: 869-874.
Direct Link - Quackenbush, L.S. and W.F. Herrnkind, 1983. Regulation of the molt cycle of the spiny lobster, Panulirus argus: Effect of photoperiod. Comp. Biochem. Phys. A, 76: 259-263.
CrossRef - Quinitio, E.T., J. De Pedro and F.D. Parado-Estepa, 2007. Ovarian maturation stages of the mud crab Scylla serrata. Aquacult. Res., 38: 1434-1441.
CrossRefDirect Link - Samuel, N.J. and P. Soundarapandian, 2010. Embryology of commercially important portunid crab Scylla serrata (Forskal). Asian J. Exp. Biol. Sci., 1: 178-182.
Direct Link