
N. Yogananth
Department of Biotechnology, Mohamed Sathak College of Arts and Science, Sholinganallur, Chennai, India
LiveDNA: 91.2037
R. Bhakyaraj
Department of Biosciences, SNMV College of Arts and Science, Coimbatore, India
LiveDNA: 91.4419
M. Syed Ali
Department of Biotechnology, Mohamed Sathak College of Arts and Science, Sholinganallur, Chennai, India
LiveDNA: 91.17314
R. Muthezhilan
Department of Marine Biotechnology, AMET University, Kanathur, Chennai, Tamil Nadu, India
LiveDNA: 91.1730
Asian Journal of Biological Sciences
Year: 2019 | Volume: 12 | Issue: 2 | Page No.: 278-283







ABSTRACT
Background and Objective: Kaempferol is the important polyphenolic flavonoids that is used to the remedy for cancers, cardiovascular disease, neuron disorder, cholesterol and serve as antioxidant and anti-inflammatory. Materials and Methods: The present investigation was the comparative analysis of kaempferol from field grown plant parts and in vitro callus and also the enhancement of kaempferol by the influence of yeast elicitor were determined. In the present study, at the concentration of 2.0 mg L–1 2,4–D+0.5 mg L–1 KIN gave the maximum callus growth for both stem and leaf explants (3.47±0.50e and 3.56±0.44e, respectively). Results: The maximum content of kaempferol in three type of samples was observed in this order; yeast elicitor treated samples > in vitro raised callus samples > field grown samples. The highest amount of kaempferol was fount in 1 mL yeast elicitor treated with stem callus sample (0.986 mg g–1 dw). Conclusion: Therefore, the present finding it was proved that the highest accumulation of kaempferol in elicitor treated in vitro callus is provides resource for preparation of drugs against various diseases by the pharmaceutical industries.
PDF Abstract XML References Citation
Received: December 01, 2018;
Accepted: January 31, 2019;
Published: March 15, 2019
How to cite this article
N. Yogananth, R. Bhakyaraj, M. Syed Ali and R. Muthezhilan, 2019. Effect of Yeast Elicitor on the Enhancement of Kaempferol from in vivo and in vitro Callus Cultures of Dregea volubilis Benth. Asian Journal of Biological Sciences, 12: 278-283.
URL: https://scialert.net/abstract/?doi=ajbs.2019.278.283
URL: https://scialert.net/abstract/?doi=ajbs.2019.278.283
INTRODUCTION
Recent days the production of secondary metabolites using plant cells has been the subject of wide investigate. Many of these metabolites are valued for their pharmacological needs and industrial or agricultural contents which improved the commercial value of medicinal herbs1, 2. Phyto constituents are sometimes very less or not measurable in field grown plant parts and de-differentiated cells such as callus tissues or suspension cultured cells.
Micropropagation is a potential source for the cultivation of essential secondary metabolites. The major trouble to arrive at a cost-effective process is the low productivity due to lack of differentiation in the cultured cells. Various approaches have been used to maximize the yield of phytochemicals produced by cultured plant cells i.e., selection of high-producing strains, addition of precursors, biotransformation, elicitor treatment, application of immobilized cells and product secretion into the culture media3. Elicitation is also one of the most successful methods employed in the induction of secondary metabolite products. The strategy of elicitation facility based on the accumulation of many phytoconstituents in plants is part of the defense responses of plants against pathogen and environmental stimuli and elicitors were referred as the agents to induce plant defense responses4.
Dregea volubilis (Asclepiadaceae) is a very well known folk medicine for antifebrile and emetic5. Also, this plant has used as a remedy to treat , abdominal tumours, aphrodisiac, carminative, sore throat, haematemesis, appetizer, heals carbuncles, eczema, asthma and antidote for poison6. The stems and leaves contain a pigment taraxerol, a triterpenoid, kaempferol, drevogenin A, drevogenin P, D-cymarose, L-oleandrose and saponins7. Kaempferol is one of the important polyphenolic flavonoids of Dregea volubilis. It is a natural plant product with potentially useful pharmacological and nutraceutical activities. It is common in vegetable, fruits, plant and herbal medicines. Over the years, studies have shown that can help treatment of cancers, cardiovascular disease, neuron disorder, cholesterol and serve as anti-oxidant and anti-inflammatory8.
The aim of the present work, to induce the callus formation from leaf and stem explants of Dregea volubilis and the comparative analysis of kaempferol from field grown plant parts and in vitro callus and also the enhancement of kaempferol by the influence of yeast elicitor were determined.
MATERIALS AND METHODS
Callus induction: Explants of Dregea volubilis were collected from medicinal plant garden, JJ College of Arts and Science, Pudukkottai and its young stem and leaves were used for establishing callus. The explants (1-2 cm) were washed thoroughly under running tap water and then treated with a few drops of Tween-80 and 1% savlon for 10 min with constant shaking. This followed by successive three washing with distilled water to make the material free from savlon. Again the explants were washed with 70% ethyl alcohol for few seconds and washed with distilled water for 3-4 times. After that, the explants were transferred to laminar air flow chamber and disinfected with 0.1% HgCl2 for 2 min and washed with sterile distilled water for 5-7 times. Then, the explants were placed in sterile Petri plates before inoculation.
The sterilized leaves and stem were wounded all over the surface and were cultured on Murashige and Skoog medium supplemented with different concentrations of 2, 4-D (1.0, 1.5, 2.0, 2.5 and 3.0 mg L–1) along with cytokinins like KIN (0.5 mg L–1). Each experiment/treatments was repeated thrice. Analysis of variance was carried out and the differences between the treatments were determined by Duncan’s Multiple Range test.
Estimation of kaempferol: One gram of air-dried stem, leaf, root and pericarp of the field grown plants and 1 g of 2.0 mg L–1 2,4-D+0.5 mg L–1 KIN callus of the stem and leaf was accurately weighed and extracted with methanol. Each methanol extract was filtered through 0.2 μm PTFE membrane filter before injection into HPLC column.
About 10 g of the yeast extract was dissolved in 50 mL to distilled water and after addition of 80% v/v ethanol. It was incubated at 6°C for 4 days and then supernatant was decanted. This process was repeated and the final precipitate was dissolved in 75 mL of distilled water and the solution of final concentrations of 0.25 and 0.8 mg mL–1 were added to the cultures9.
About 1 g of dry powdered stem, leaf, root, pericarp of D. volubilis containing 31 mL of distilled water inoculated tubes were incubated for 240 h at 25-25°C at 200 rpm. About 2 mL of the sample were collected at 240 h and extracted with 10 mL methanol. The extract was analyzed by HPLC. The stem and leaf derived callus were sub-cultured on MS media supplemented with 2,4-D 2.0 and 0.5 mg L–1 KIN. The media were impregnated with yeast extract (1, 2 and 3 mL L–1) as an elicitor was tested on the same MS media. The calli were left to grow for a period of two weeks. After 14 days, the callus cultures were harvested and 200 mg of fresh wet tissues were extracted in 2 mL of absolute methanol for 24 h. The extracts were centrifuged at 10.000 rpm for 15 min and collect the supernatant.
The HPLC analysis of the different powdered samples i.e., field grown plant culture, callus culture and elucidated culture were carried out with JASCO HPLC, using a C-18 μ Bondapak column and acetonitrile: Water (50:50)+0.1% phosphoric acid as solvent system at a flow rate 0.8 mL min–1 and detector set at 350 nm. The kaempferol was identified by comparison of its retention time with those of authentic kaempferol samples (received from Sigma Aldrich, Bangalore, India) and quantified using a standard calibration curve. Kaempferol yields were calculated using the total dry weight of tissue and expressed in terms of percentage of kaempferol content.
RESULTS
Callus induction was observed in MS media containing different concentrations and combination of 2,4-D with Kin. Within 2 weeks of incubation the explants of stem and leaves depending upon the concentrations and combination of hormone were induced calli.
There was a range of variation in percentage of callus formation and average fresh weight of callus. The stem explants showed highest percentage of callus induction (85.03±0.78d) in MS medium containing 2.5 mg L–1 2,4-D and 0.5 mg L–1 Kin and followed by 82.66±0.33d in MS medium containing 3.0 mg L–1 2,4-D and 0.5 mg L–1 Kin. But, the leaf explants showed the highest callus growth in terms of fresh weight (3.56±0.44 g) and dry weight (0.33±0.03e g) was observed in MS medium fortified with 2.0 mg L–1 2,4-D and 0.5 mg L–1 Kin (Table 1).
Among the four field grown plant parts tested, the stem extract contained maximum content of kaempferol (0.528 mg g–1 dw) and minimum was observed in pericarp (0.088 mg g–1 dw). The content of isolated flavonoids in various plant parts was observed in this order; stem>leaf> root>pericarp (Table 2, Fig. 1).
The kaempferol analysis of callus, the maximum kaempferol content was observed in stem derived callus extracts (0.947 mg g–1 dw) whereas the kaempferol content in the leaf derived callus extracts were 0.586 mg g–1 dw (Table 3, Fig. 2). From the results of callus induction, the concentration of 2.0 mg L–1 2,4-D and 0.5 mg L–1 Kin gave the maximum callus growth to both stem and leaf explants of Dregea volubilis. Hence these callus cultures used for enhancement experiment of kaempferol by using yeast elicitor.
Table 4 was shown the result of kaempferol enhancement from field grown and callus cultures of Dregea volubilis. In yeast elicitor treatment, the stem callus produced the highest quantity of kaempferol (0.986 mg g–1 dw) in 1 mL yeast elicitor treatment and the minimum quantity of kaempferol was observed in Pericarp+1 mL elicitor treatment (0.345 mg g–1 dw) (Fig. 3).
DISCUSSION
The media containing high auxin and low cytokinin concentrations promote cell proliferation resulting in callus formation10. In this present study, callus cultures were established from young leaf and stem explants of D. volubilis Benth. cultured on MS medium+B5 vitamins with varying concentrations of 2, 4-D (1.0-3.0 mg L–1) along with BAP (0.5 mg L–1). Generally, the 2,4-D can react with the plant cell RNA metabolism by inducing the transcription of messenger RNA capable of coding proteins required for the growth and therefore, promoting a chaotic cell proliferation, i.e., callus formation11.
Secondary metabolites are mainly responsible for the pharmacological properties of plants2. The secondary metabolite production was mainly restricted to particular species or genus and to particular stage of plant growth. It also depends on external factors such as soil fertility and seasonal variations12 besides seasonal variations, majority of plant based pharmaceutically important secondary metabolites are produced in higher quantities under controlled conditions using tissue culture techniques13.
| Table 1: | Effect of 2,4-D with KIN on callus induction of Dregea volubilis |
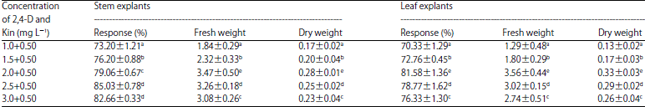 | |
| ±: Standard error, Means followed by the same letter not significantly different by the DMRT at p<0.05 level of significance | |
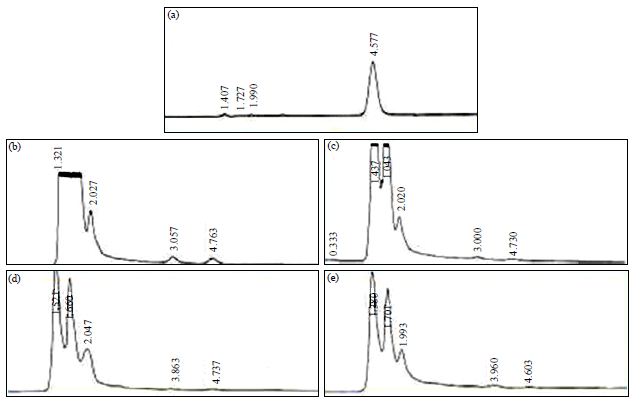 | |
| Fig. 1(a-e): | HPLC chromatogram of kaempferol standard and field grown plant parts (a) STD, (b) Stem, (c) Leaf, (d) Root and (e) Pericarp |
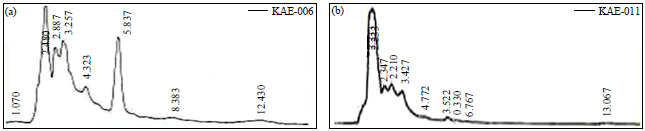 | |
| Fig. 2(a-b): | HPLC chromatogram of (a) Stem callus and (b) Leaf callus extracts |
| Table 2: | Kaempferol production from field grown plant parts |
 | |
| Table 3: | Kaempferol estimation from callus extracts |
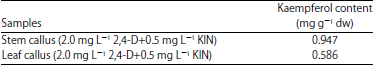 | |
In the present study, the estimation of field grown plant parts and in vitro raised callus showed a difference in kaempferol yield. In field grown plants, the stem explants gave maximum amount of kaempferol yield than other parts tested. In vitro raised callus showed highest kaempferol when compared to in vivo developed plant parts.
| Table 4: | Effect of yeast elicitor on kaempferol production from field grown and callus cultures of D. volubilis |
 | |
Similarly, Sudria et al.14 reported that auxin and cytokinin incorporated into culture medium have a marked influence on the production of secondary metabolites. Yogananth et al.15 reported solasodine production in Solanum nigrum on MS medium supplemented with different concentrations of IAA, NAA and BAP. They found that solasodine content was higher in callus cultures than field grown leaves.
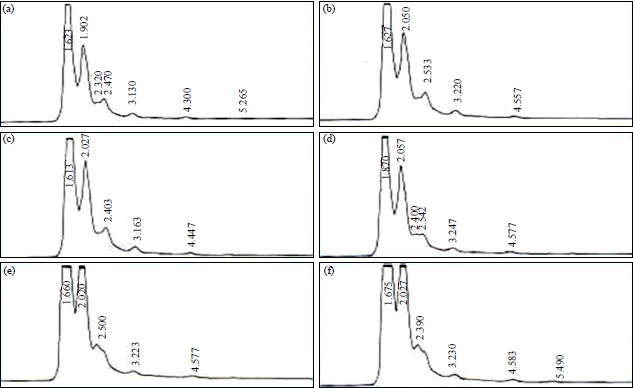 | |
| Fig. 3(a-f): | HPLC chromatogram of elicitor treated field grown plant parts and callus cultures (a) Stem, (b) Leaf, (c) Root, (d) Pericarp, (e) Stem callus and (f) Leaf callus |
In the present findings, the treatment of plant powder and callus suspensions with a crude extract of yeast elicitor improved production of kaempferol when compared to field grown plant parts and in vitro raised callus. The present study is supported by the findings of previous workers, plumbagin production from Drosera sp. by using the yeast elicitor16 and accumulations of silymarin in Silybum marianum by using yeast extract17 reported that the production of berberine by Thalictrum rugosum cells with yeast elicitor was increased up to four times and the production of alkaloids by Eschscholtzia californica was increased about 30 times.
CONCLUSION
The kaempferol analysis in three different ways, the elicitor treated callus culture gave maximum amount of kaempferol when compared to without elicitor treated in vitro raised callus and field grown plant parts. It indicates that yeast might be used as a permeabilizing agent to acquire kaempferol on a large scale without sacrificing the cells. However, the further studies regarding the identification of signaling component of yeast extract in biosynthesis could be a very effective approach for large scale augmentation of alkaloid yield of pharmaceutical importance.
REFERENCES
- Bingham, S.A., C. Atkinson, J. Liggins, L. Bluck and A. Coward, 1998. Phyto-oestrogens: Where are we now? Br. J. Nutr., 79: 393-406.
CrossRefDirect Link - Paganga, G., M. Miller and C.A. Rice, 1999. The polyphenolic content of fruit and vegetables and their antioxidant activities. What does a serving constitute? Free Radical Res., 30: 153-162.
CrossRefDirect Link - Zhou, L.G. and J.Y. Wu, 2006. Development and application of medicinal plant tissue cultures for production of drugs and herbal medicinals in China. Natl. Prod. Report, 23: 789-810.
PubMed - Panda, N., N.B. Mandal, S. Banerjee, N.P. Sahu and K. Koike et al., 2003. Polyhydroxy pregnanes from Dregea volubilis. Tetrahedron, 59: 8399-8403.
CrossRefDirect Link - Sahu, N.P., N. Panda, N.B. Mandal, S. Banerjeea, K. Koike and T. Nikaido, 2002. Polyoxypregnane glycosides from the flowers of Dregea volubilis. Phytochemistry, 61: 383-388.
CrossRefDirect Link - Venkata-Rao, D. and E. Venkata-Rao, 1971. Pregnane derivatives of Marsdenia volubilis T. cook. Cur. Sci., 40: 193-194.
Direct Link - Chen, H., F. Chena, F.C.K. Chiu and C.M.Y. Lo, 2001. The effect of yeast elicitor on the growth and secondary metabolism of hairy root cultures of Salvia miltiorrhiza. Enzyme Microbial Technol., 28: 100-105.
CrossRefDirect Link - Oksman-Caldentey, K.M. and D. Inze, 2004. Plant cell factories in the post-genomic era: New ways to produce designer secondary metabolites. Trends Plant Sci., 9: 433-440.
CrossRefPubMedDirect Link - Wang, C., J. Wu and X. Mei, 2001. Enhanced taxol production and release in Taxus chinensis cell suspension cultures with selected organic solvents and sucrose feeding. Biotechnol. Prog., 17: 89-94.
CrossRefDirect Link - Sudria, C., M.T. Pinol, J. Palazon, R.M. Cusido and R. Vila et al., 1999. Influence of plant growth regulators on the growth and essential oil content of cultured Lavandula dentata plantlets. Plant Cell Tissue Organ Cult., 58: 177-184.
CrossRefDirect Link - Yogananth, N., R. Bhakyaraj, A. Chanthuru, S. Parvathi and S. Palanivel, 2009. Comparative analysis of solasodine from in vitro and in vivo cultures of Solanum nigrum Linn. Kathmandu Univ. J. Sci. Eng. Technol., 5: 99-103.
CrossRefDirect Link - Sanchez-Sampedro, M.A., J. Fernandez-Tarrago and P. Corchete, 2005. Yeast extract and methyl jasmonate-induced silymarin production in cell cultures of Silybum marianum (L.) Gaertn. J. Biotechnol., 119: 60-69.
CrossRefDirect Link