
A.A. Wahizatul
Department of Biological Sciences, Faculty of Science and Technology, Universiti Malaysia Terengganu, 21030 Kuala Terengganu, Terengganu, Malaysia
J.T. Jennings
School of Earth and Environmental Sciences, The University of Adelaide, South Australia 5005, Australia
Asian Journal of Biological Sciences
Year: 2011 | Volume: 4 | Issue: 5 | Page No.: 428-444







ABSTRACT
The principal objective of this study was to compare differences in species richness and abundance of aquatic invertebrate communities between natural willow roots and artificial aluminium wire mesh substrates. A field experiment was conducted at Deep Creek in the Mount Lofty Ranges (South Australia) to investigate colonisation of willow roots by aquatic invertebrates. Two different types of substrate with differing degrees of complexity were used. Willow roots with only primary roots left intact; lateral and primary roots left intact; and no roots were cut down and aluminium wire mesh of three different mesh sizes: coarse (15 mm), medium (9 mm) and fine (1.5 mm), were allowed to colonise for 30 and 90 days. Aquatic invertebrates were significantly more abundant on willow roots than on aluminium wire mesh substrates. However, there was no significant difference in species richness. Habitat complexity and period of colonisation are important determinants of invertebrate community structure on each substrate type. Scrapers (primarily gastropods) often constituted the greatest abundance on willow roots. The introduced hydrobiid snail, Potamopyrgus antipodarum, was significantly more abundant especially on the most complex of willow root structures and after a longer period of colonisation (90 days). Findings of this study suggested that willow roots have habitat value for aquatic invertebrates, especially the hydrobiid snail and decrease in structure associated with willow roots resulted in a lower abundance of invertebrates. Carefully management of willow removal should take into account the sudden removal of willow roots which may disrupt the invertebrate communities which utilise the roots as habitat. It is also evident that a long period of colonisation is needed to obtain a stable invertebrate community, especially during revegetation efforts.
PDF Abstract XML References Citation
Received: March 03, 2011;
Accepted: May 26, 2011;
Published: July 16, 2011
How to cite this article
A.A. Wahizatul and J.T. Jennings, 2011. Impact of Exotic Willow Roots (Salix spp.) as Habitat for Aquatic Invertebrate Communities in South Australian Stream. Asian Journal of Biological Sciences, 4: 428-444.
URL: https://scialert.net/abstract/?doi=ajbs.2011.428.444
URL: https://scialert.net/abstract/?doi=ajbs.2011.428.444
INTRODUCTION
Since the early 1900s, thousands of kilometres of the riparian zones of many south-eastern Australia have become dominated by the exotic willow trees (Holland and Davies, 2007). They have been mainly used for erosion control, bank stabilization, boat navigation and as ornaments (Ladson et al., 1997; Holland and Davies, 2007). Since then, as more willows have become established, greater changes in hydrologic regimes, problems with water quality, decline in native species and establishment of exotic weeds have been reported by Merigiano (1996) and Patten (1998).
The ability of willow roots to spread into river and stream banks can have long term effects on channel morphology. Willow roots trap silt and layer new roots over old roots in stream channels, building up the streambed and creating a broad shallow stream (Bunn et al., 1999; Ladson et al., 1997). Willow roots can be associated with altering bank morphology, changing channel depth and width, flow and sediment conditions (Bunn et al., 1993; Lester et al., 1994a; Ladson et al., 1997). Willow roots are also presumed to be an important component responsible for a decline in native biodiversity by decreasing habitat heterogeneity for fish, aquatic plants and aquatic invertebrates (Collier, 1995; Schulze and Walker, 1997; Al-Sherif et al., 2009). Willows are now often being poisoned, removed from accessible river banks and the areas replanted with native vegetation. However, such programs have accelerated bank erosion, leading to water quality problems and allowing exotic weeds to become established.
The removal of willows directly destroys the thick mats of willow roots. Thick willow roots mats can play an important role in the formation and maintenance of habitat for aquatic fauna. Adult stream fish have been reported to use the roots as a habitat source (Pusey et al., 1998), whilst exposed roots have been used as a spawning substrate and larval habitat (Pusey et al., 2001). However, little is known about the value of willow roots as suitable habitat for aquatic invertebrate communities. Some studies suggest that willows may support high numbers of aquatic invertebrates by providing suitable habitat. For example, Glova and Sagar (1994) found greater species richness and diversity in benthic invertebrate communities in reaches lined by willows in three New Zealand streams. Jayawardana et al. (2006) found P. antipodarum were commonly associated with willow root habitats in central Victorian streams in Australia. Willow root mats have greater surface texture, area and provide a variety of microhabitats for invertebrates colonisation. The rough-fibrous texture of willow roots is known to affect colonisation by aquatic invertebrate communities and provides a suitable habitat for invertebrate reproduction and protection from predators (Lester et al., 1994a).
In this study, the principal objective was to compare differences in species richness and abundance of aquatic invertebrate communities between natural willow roots and artificial aluminium wire mesh substrates. Artificial substrates have been used to sample benthic macroinvertebrates in similar habitats in lotic systems (Souter and Williams, 2001; Souter, 2004; White and White, 2005). Aluminium wire mesh was chose as an artificial substrate to mimic microhabitats found in willow roots because it is manufactured in various mesh sizes, is easy to manipulate and when placed on the bottom of a stream, is relatively stable in currents. Generally, heavier or bottom-sited artificial substrates are recommended because they are considered more reliable than artificial floating substrates which tend to be selectively colonised by beetles, mayflies and caddisflies. Furthermore, artificial floating substrates (e.g., leaf packs/bags) are usually positioned near the stream bed and often float for a time before sinking to the bottom which may affect the representation of invertebrate communities (Boulton and Boon, 1991). Although Souter (2004) used bricks to weigh down his artificial substrates, we used metal stakes to hold down the artificial substrates which will also avoid the possibility of invertebrates using bricks as habitat.
This study was aimed to investigate differences in aquatic invertebrates colonisation on two different substrate types (willow roots vs. aluminium wire meshes), each with three different habitat complexities and for colonisation periods of 30 and 90 days. Previous studies found that natural substrates usually supported higher densities and diversity of aquatic invertebrates than artificial substrates. For example, Casey and Kendall (1996) found that the quantity of organic material and epilithic cover was greater on natural substrates than artificial substrates. Other study done by Zorriasatein et al. (2009) found greatest numbers of species of benthic fauna were recorded at 1st station in Gorganrood River, South Eastern Caspian Sea. They suggested that water quality and natural habitat heterogeneity play important role to support higher density and diversity of benthic organisms.
Therefore, willow roots were predicted would be preferred over artificial aluminium wire mesh substrates, with a greater abundance and species richness of aquatic invertebrates, as natural willow roots provide both more heterogeneous microhabitats and organic material for invertebrates colonisation.
MATERIALS AND METHODS
Site description: In situ experiments were conducted in Deep Creek in the Mt. Lofty Ranges, South Australia. Deep Creek is located at around 350 m elevation with latitude 34°56’N and longitude 138°46’E. This small fourth order stream is approximately 8.5 km long, is 2.0-4.5 m in width, 0.5-2.5 m in depth, is slow to sometimes fast flowing and the catchment area is ~13 km2. The stream bed is almost debris free, mainly comprising sand, pebbles, cobbles and small boulders. Aquatic vegetation is mainly Carex sp., Juncus sp. and Rorrippa sp. The major riparian vegetation in the study area consists of willows, Eucalyptus viminalis, E. obliqua, Acacia retinoides and A. melanoxylon. Generally, the upper reaches of the stream are exposed to direct sunlight where the water is deep and slow moving and the bed mostly sandy. The lower reaches are partly cleared and dominated by willows.
The climate in the Mount Lofty Ranges catchment area is hot Mediterranean. The average annual rainfall in the catchment areas is approximately 900 mm, with average maxima of 25°C (soaring into the 40 sec) during dry summers and with maximum temperatures averaging 12 to 15°C during cool, wet winters. Both study catchments include a mixture of rural residential, horticulture, orchards and grazing land. During the study period, the mean water temperature ranged from 15.70-23.60°C, pH ranged from 8.15-9.84, dissolved oxygen ranged from 10.81-16.25 mg L-1, conductivity ranged from 271-422 μS cm-1 and total dissolved solids ranged from 102-275 ppm (AHNRC, 2009).
Experimental designs: The experimental design was a 2x6x2 level factorial design, with 4 replicates per sub-class. Treatments were two types of substrate (willow roots vs. aluminium wire mesh), six different habitat complexity and two periods of colonisation (30 and 90 days). Fresh willow roots were collected from Deep Creek and air dried at ambient temperatures for 2-3 weeks. Three different habitat complexities were prepared for the willow root treatments: all fine and lateral roots were cut down with only primary roots left intact (WR1); only fine roots were cut down and lateral and primary roots were left intact (WR2); and no roots were cut down (WR3). For the artificial substrate, aluminium wire mesh of three different mesh sizes ranging from: coarse (15 mm; AS1), medium (9 mm; AS2) and fine (1.5 mm; AS3), were used as the three treatments. Replicate of the substrates were constructed from bundles of willow roots or aluminium wire mesh of 10-15 cm long and 5-10 cm diameter (total surface area 50-150 cm2). Each replicate was placed in a galvanized wire mesh cage (mesh size: 3 mm, 15x10 cm) to avoid being washed away from high flows. As most of invertebrates quickly leave the cages when it was removed, each substrate was wrapped with nylon onion bags before placing in the cages. Thus, all the invertebrates could be retrieved without the invertebrates escaping. There were four replicates of each treatment and a total of 48 cages were placed randomly in a uniform section of Deep Creek with an open canopy. Cages were anchored by metal stakes and marked to facilitate recovery.
Cages were collected twice to observe variations in colonisation by invertebrates from 30 days and 90 days. Between 1200 and 1700 h on 19th December 2008 (Day 30) and again on 21st February 2009 (Day 90), four cages of each different substrates complexity were randomly selected and removed from the stream (n = 24 cages). For each cage, samples were collected by placing a fine-meshed net (250 μm) directly downstream of the cage and quickly placing the cage into the net. The cage and net contents were transferred to individual plastic bags filled with 80% ethanol and returned to the laboratory for analysis.
The cage substrate was washed carefully to remove any attached sediment particles and invertebrates into 250 ìm mesh screen, fixed with 80% ethanol for sorting and enumeration. All invertebrates were sorted into functional feeding groups and then identified as far as possible using keys of Williams (1980), McCafferty (1981), Merritt and Cummins (1996), Watts (1998), Gooderham and Tsyrlin (2002), Dean et al. (2004) and Theischinger and Hawking (2006).
Statistical analysis: Two-way nested factorial ANOVAs were used to analyse differences in community distribution between substrate type (2 levels, fixed), habitat complexity (six levels, random, nested within substrate types) and days of colonisation (two levels, random). Data were log(x+1) transformed to ensure normality in calculations of means and ANOVAs. Pair-wise comparisons, using Bonferroni post hoc tests were used to compare means to determine between which treatments were different. The same model of two-way ANOVAs was applied to estimate differences in the abundance of functional feeding groups among types of habitat complexity and days of colonisation. All of the analyses were performed using the Prism statistical program (PRISM, 2007). Non-parametric multivariate analysis of variance was used to analyse differences in the invertebrate community assemblages among substrate type, habitat complexity and days of colonisation, using two-way factorial permutation-based nonparametric MANOVA (PerMANOVA; Anderson and Braak, 2003). The Sorensen Bray-Curtis test was used as a distance measure and 4999 permutation of raw data units were used for each analysis (McCune and Mefford, 2006). Detrended Correspondence Analysis (DCA) which ordinates both species and samples simultaneously was used to identify the species assemblage ordination which could be then related to measure how much the species distribution differed along types of substrate. One-way and two-way cluster analysis using Sorenson’s Coefficient method was used to sort species of invertebrate communities into groups to reveal the degree of associations between generic composition and the effects of habitat complexity and days of colonisation (Kovach, 1999). This method sort cases in groups or cluster, so that the degree of association is strong between members of the same cluster and weak between members of different clusters. PerMANOVA, DCA and clustering analyses were performed using the statistical program of PC-ORD version 5.13 developed by McCune and Mefford (2006).
RESULTS
Effect of substrate type: A total of 9935 individuals from 29 species belonging to 25 families of aquatic invertebrates were identified from the different treatments of substrate types in Deep Creek (December 2008 to February 2009). There were significant differences in the total abundance between willow roots and aluminium wire mesh substrates treatments (F1,47 = 15.275, p<0.001) (Table 1). Total abundance was significantly higher on willow root substrates (7564 individuals) compared with aluminium wire mesh substrates (2371 individuals). However, there was no significant difference in species richness of aquatic invertebrates between substrates (F1,47 = 14.771; p = 0.2462).
| Table 1: | Results of two-way ANOVAs on total abundance and species richness of aquatic invertebrates at different substrate type, habitat complexity and periods of colonisation |
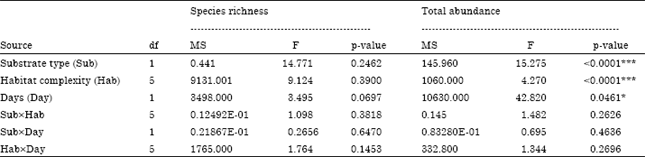 | |
| df: Degree of freedom, MS: Mean squares, ***p<0.0001; *p<0.05 | |
| Table 2: | Results of two-way factorial PerMANOVAs on aquatic invertebrate community assemblages at different types of substrate, substrate complexity and periods of colonisation |
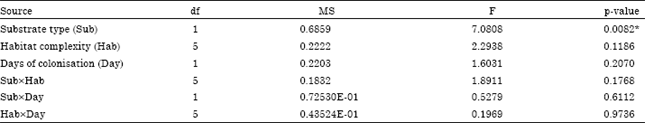 | |
| df: Degree of freedom, MS: Mean squares, *p<0.05 | |
Similar trends in relative abundances (%) for the major taxonomic groups of aquatic invertebrates were observed on both substrates. Gastropoda (mostly the introduced hydrobiid snail, P. antipodarum) was the most dominant group, making up about half of the total abundance but more were collected on willow roots than aluminium wire mesh substrates. Plecoptera was the second most abundant group while Trichoptera and Ephemeroptera were third and fourth highest on both substrates. More Coleoptera, Diptera and Oligochaeta were recorded on willow roots than on aluminium wire mesh substrates. However, slightly more Odonata were found on aluminium wire mesh than on willow roots. Other groups such as Bivalvia, Ostracoda and Hirudinea were uncommon on both substrates. However, the effect of substrate type on invertebrate community assemblages did not vary significantly among habitat complexity or for colonisation periods (Sub x Hab: F5,47 = 1.8911, p = 0.1768; Sub x Day: F1,47 = 0.5279; p = 0.6112) (Table 2).
Of the 29 species found, 13 species were classified as collector-gatherers, 7 species were shredders and 8 species each for predators and scrapers. Only 4 species were identified as collector-filterers. However, some of the species can be included into two or three functional feeding groups (e.g., Oecetis sp., Simsonia leai) (Table 3). Scrapers were dominated by the introduced hydrobiid snails, P. antipodarum, in all treatments and were significantly greater on willow roots than on the aluminium wire mesh substrates (F5,47 = 0.491, p<0.0001). There were clear differences among other feeding groups, with the shredders and collector-gatherers colonising rapidly on the willow roots while predators and collector-filterers colonised more slowly on both types of substrates. The abundance of shredders (e.g., D. evansi, L. aurata) was greater on willow roots than on the aluminium wire mesh substrates. However, the abundance of predators (e.g., chironomids, Hemicorduliidae) was greater on the aluminium wire mesh substrates compared with willow roots. Collector-gatherers and collector-filterers were in very low numbers in all substrates.
Effect of habitat complexity: The total abundance of aquatic invertebrates varied significantly from one habitat complexity to another (F5,47 = 4.270, p<0.0001) (Table 1). Further analysis using Bonferroni multiple comparison tests showed that the total abundance of invertebrates on aluminium wire mesh substrates (AS3, AS2 and AS1) was significantly lower than on willow roots (WR3, WR2 and WR1) at p<0.01. However, habitat complexity did not influence the invertebrate community assemblages for different periods of colonisation (F5,47 = 0.1969, p = 0.9736) (Table 2).
More invertebrates were found on soft, fibrous willow roots (WR1, WR2 and WR3) than on firm, smooth artificial aluminium wire mesh (AS1, AS2 and AS3). Invertebrates were more abundant on intact willow roots (WR3; 3874 individuals) compared with roots where fibrous fine roots had been removed (WR2; 2246 individuals and WR1; 1444 individuals). On aluminium wire mesh, quite high numbers of invertebrates were recorded on the fine aluminium wire mesh (AS3; 1727 individuals), although lower numbers were found on medium and coarse mesh sizes (AS2; 474 individuals and AS1; 170 individuals).
There were no significant differences between species richness and habitat complexity (F5,47 = 9.124; p = 0.3900) (Table 1). Similarly, no interaction for species richness was detected between habitat complexity and periods of colonisation (F5,47 = 1.764; p = 0.1453). AS3 (fine size of aluminum wire mesh) was found to support the most diverse invertebrate community as 23 species were identified (Table 3). A total of 18 species were found on AS2 (medium mesh size) and 15 species on AS1 (coarse mesh size). Willow roots (WR3, WR2 and WR1) were represented by 19 species in each treatment.
Both habitat complexity and periods of colonisation significantly affected the number of scrapers, shredders and predators. Scrapers were much more abundant on soft, fibrous willow roots (WR3, WR2 and WR1) than on the firm, smooth aluminum wire mesh. Scrapers were significantly more abundant on willow roots on aluminum wire mesh. Of the eight species of scrapers recorded, P. antipodarum was the dominant species in all willow root treatments, indicating to a strong preference to use willow roots. Shredders were dominated by D. evansi, L. aurata and hydroptilid caddisfly larvae. Dinotoperla evansi and hydroptilids were found on willow root treatments WR2 and WR3, whereas L. aurata which have, in general, similar feeding habits, occurred mostly on AS3. Higher numbers of predators were recorded on AS3, especially the damselfly nymphs (Pseudagrion sp.) and the chironomids. Of the three common species of predatory chironomids, Procladius sp. were found more on AS3, whereas Polypedilum sp. and Paramerina sp. appeared to be more on willow roots. However, their abundance was not statistically significant. Collector-gatherers (e.g., K. inconspicua and T. tillyardi) were more abundant on all aluminium wire mesh treatments, although no significant differences were detected. Collector-filterers, like the bivalves and chironomids, were very low in numbers in most treatments but more of them were collected on AS3.
Species abundances and species assemblage ordinations showed marked variation in relation to the types of substrate and habitat complexity (Fig. 1). The most abundant species was P. antipodarum which was significantly more abundant on all willow root treatments, especially on WR3. It was followed by gripopterygid stonefly larvae (D. evansi) and hydroptilid caddisfly larvae which were also significantly more abundant on willow root treatments. Other major species, such as lephtophlebiid mayfly larvae (K. inconspicua), caenid mayfly larvae (T. tillyardi), conoesucid caddisfly larvae (L. aurata) and physid snails (Ph. acuta) were very commonly found on all types of substrate complexity. In willow root treatments, the coenagrionid damselfly nymphs (Pseudagrion sp.) and leptocerid caddisfly larvae (Oecetis sp.) appeared to be particularly uncommon but more were found on aluminium wire mesh substrates (AS3 and AS2). Other species (e.g., S. leai, Isidorella sp.) were considered as minor since less than 49 individuals of each species were recorded.
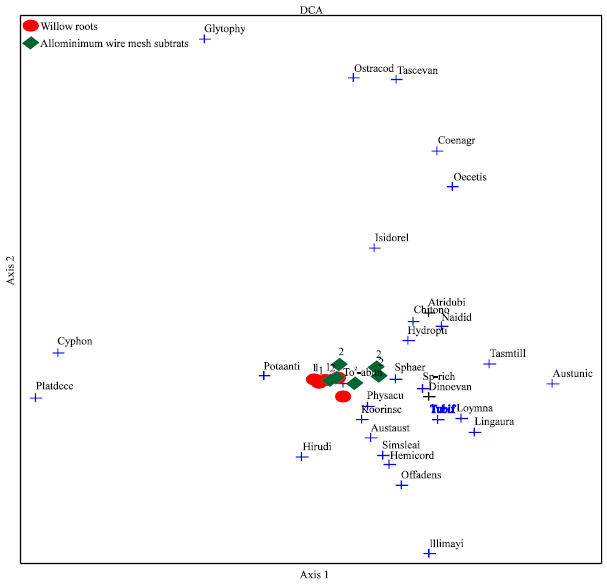 | |
| Fig. 1: | Detrended correspondence analysis (DCA) plots showing species assemblage ordinations which related to the types of substrate |
The least common species were telephebiid dragonfly nymphs (Austroaeschna unicornis), lymnaeid snails, ostracods and amphipods, each represented by less than 5 individuals throughout the study.
Effect of periods of colonisation: There were significant differences in the total abundance of aquatic invertebrates between periods of colonisation (F1,47 = 42.820, p<0.05) (Table 1). As predicted, more invertebrates were found after 90 days than after 30 days of colonisation. After 90 days, invertebrates were markedly higher on WR3 with 2397 individuals and considerably higher in other willow root treatments (WR2 = 1477 individuals; WR1 = 1339 individuals). Relatively high levels of abundance occurred on AS3 with 1316 individuals but abundance was drastically reduced on AS2 (355 individuals) and AS1 (105 individuals). The highest abundance after 30 days was observed on WR3 with 1220 individuals, followed by WR2 and WR1, with 769 individuals and 362 individuals respectively. However, significantly lower total abundances were recorded in aluminium wire mesh substrates after 30 days of colonisation.
| Table 3: | List of aquatic invertebrates, their functional feeding groups and species abundance in relation to the type of substrates and substrate complexity |
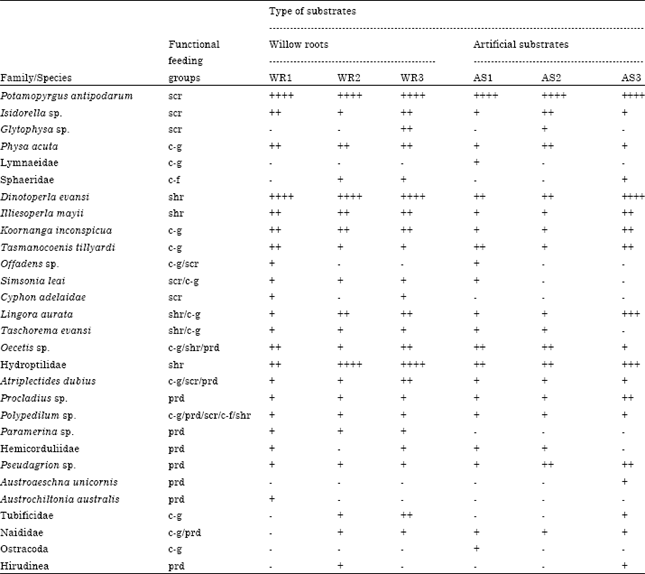 | |
| scr: Scraper, c-g: Collector-gatherer, c-f: Collector-filterer, shr: Shredder, prd: Predator, ++++: Very abundant (>100 individuals), +++: Abundant (50-99), ++: Common (10-49 individuals), +: Rare (1-9 individuals), -: Absent (none) | |
The total abundance on AS3 was the highest (411 individuals) and the abundance declines on AS2 (119 individuals) and reached its lowest level on AS1 (66 individuals).
In terms of species richness, a slightly richer and more diverse fauna were found after 90 days than 30 days. However, habitat complexity and days of colonisation did not influence the species richness of the invertebrate fauna (F5,47 = 1.764; p = 0.1453) (Table 1). The fine aluminium wire mesh substrate (AS3) had a richer fauna than other treatments with 23 species recorded after 90 days. Interestingly, only 19 species colonised each of the willow root treatments by 90 days, although many species were highly abundant (Table 3). Again, the fine aluminium wire mesh substrate (AS3) recorded the richest fauna after 30 days with 21 species, whereas other treatments were considerably lower with the lowest diversity being on AS2 with only 11 species.
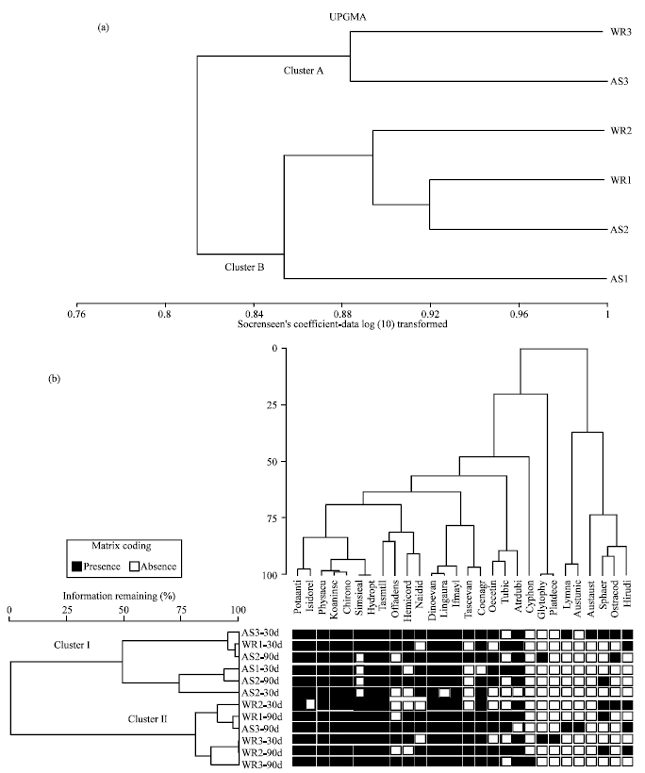 | |
| Fig. 2: | Dendrograms using UPGMA Sorensen’s Coefficient method for one-way (a) and two-way (b) clustering analysis of individuals and generic composition in different substrates, habitat complexity and days of colonisation. (WR: Willow roots, AS: Artificial substrates, 30d: 30 days of colonisation, 90d: 90 days of colonisation, Note that not all taxa are included. Taxa with fewer than 5 individuals, such as bivalves and amphipods, were excluded) |
Sorensen’s coefficient method for one-way and two-way clustering analysis was used to examine differences in habitat complexity and time of colonisation and is represented by the dendograms (Fig. 2). The importance of habitat complexity on the composition of the invertebrates is illustrated by one-way clustering analysis (Fig. 2a) in which all samples from WR3 and AS3 fall into Cluster A. Platynectes decempunctatus, Au. unicornis and Cyphon adelaidae were only found on more complex habitats (WR3 and AS3; Cluster A). Cluster B contains the samples from less complex habitats (WR2, WR1, AS2 and AS1). However, AS1 was well separated from the other samples in Cluster B, probably due to its lowest total abundance and species richness compared with other samples.
The interaction between habitat complexity and periods of colonisation, however, differ with the dendogram of two-way cluster analysis (Fig. 2b), in which two main groups were found. Most samples from the aluminium wire mesh substrates (AS1, AS2 and AS3) of each colonisation period (30 and 90 days; except for AS3) fall into Cluster I together with the sample least complex of willow root habitats after 30 days (WR1). The remaining samples, willow root treatments (WR1, WR2 and WR3) from each colonisation period (30 and 90 days; except for WR1) and the sample of fine mesh size of aluminium wire substrate (AS3) after 90 days form the other group (Cluster II).
Generally, invertebrates among the treatments were similarly distributed as the average Sorenson similarity coefficient values were medium to high, ranging from 0.50 to 0.95. Sorenson's coefficient is an index of community similarity, ranges from 0 to 1 with higher values indicating a higher degree of similarity. Thus, a coefficient of 1 means complete overlap of species between two communities and 0 means no overlap. In this study, similarity was determined within types of habitat complexity (i.e., WR1, AS1, etc.) and between days of colonisation (30 and 90 days). The samples from willow roots (WR1, WR2 and WR3) are not more distinct from the samples of aluminium wire mesh substrates (AS1, AS2 and AS3) (Fig. 2a). This was due to the similar distribution of certain dominant species among all treatments (e.g., P. antipodarum, Ph. acuta, D. evansi, T. tillyardi).
However, Cluster I was well separated from Cluster II (Fig. 2b). This separation is mainly due to the presence/absence of several species. For example, baetid mayfly larvae (Offadens sp.), elmids beetle larvae (S. leai), conoesucid caddisfly larvae (Taschorema evansi) and tubificids were absent from most samples in Cluster I. Whereas, some species such as telephebiid dragonfly nymphs (Au. unicornis) and scirtid beetle larvae (Cy. adelaidae) were only present in Cluster II. In terms of periods of colonisation, most species were more abundant on all substrates after 90 days than after 30 days. The more complex of willow root habitats (WR2 and WR3) and aluminium wire mesh habitat (AS3) exhibit a greater similarity to each other in terms of frequent species composition than the less complex habitats (WR1, AS1 and AS) after 30 and 90 days.
DISCUSSION
Overall, willow roots supported significantly higher total numbers of aquatic invertebrates by providing a better habitat than the aluminium wire mesh substrates (Table 1). As well, more species were recorded on willow roots than on aluminium wire mesh substrates but the latter difference was not significant (Table 1). No consistent differences were detected in the mean number of species among the treatments tested (Table 2). Thus, our first hypothesis was partially supported: the abundance of aquatic invertebrates was higher on willow roots than on aluminium wire mesh but this was not the case for species richness or species differences.
Willow roots have habitat value for aquatic invertebrate communities. Willow root mats are fine, rough, fibrous texture and extend into the water column and provide a suitable habitat for invertebrate reproduction, protection from predators and food supply. Willow roots can be colonised by beneficial bacteria and naturally occurring food organisms (biofilms) and these biofilms and epilithics material (including algae, bacteria, fungi, detritus, sand and silt) are known to affect colonisation by aquatic invertebrate communities (Mackay, 1992). Davoudi (2011) reported that willow root is recommended as an effective technique for preventing superficial sliding in Iran. In this study, most species were greater in number on rough, fibrous substrates than on smooth, firm substrates, indicating that inorganic or artificial aluminium wire mesh is less suitable as a site of attachment and less able to provide a food source than natural willow roots. Willow root treatments accumulated a much thicker slimy microbial biofilm and epilithics material compared with that on aluminium wire mesh. To date, there have been no studies on the amount of periphyton growth or detritus accumulation on the willow roots or aluminium wire mesh substrates, although Casey and Kendall (1996) found that densities of benthic macroinvertebrates and the quantity of organic material were greater on a natural substratum than on artificial substrate types (single particles of natural rock or clay brick). They suggested that the development of an invertebrate community on a substrate is linked to both the mobility of different species and the accumulation of food sources, i.e., periphyton and organic detritus, on the substrates. Other investigators who studied artificial substrates (Souter and Williams, 2001; Souter, 2004) did not measure organic material (e.g., biofilms, epilithics material, etc.) on the substrates. Thus, for future research, investigators should consider the potential effect of the natural epilithic cover of substratum particles might have on colonisation by invertebrates when choosing the type of artificial substrates.
In terms of relative abundance of the major taxonomic groups, we found the gastropods (primarily scrapers) were significantly greater (p<0.0001) on willow roots than on aluminium wire mesh, although this was primarily due to a single species, P. antipodarum which contributed more than half of the total abundance. Jayawardana et al. (2006) also found that willow root habitats facilitate the high abundance of P. antipodarum in three streams in Central Victoria. This alien species has rapidly invaded Australian freshwater systems and potentially impacts on native invertebrate assemblages include domination of preferred habitat, physical covering of egg sites and egg masses, harbouring and of molluscivorous predators and attraction of fish predators (Ponder, 1988; Bowler, 1991). Usually, invasive species always caused negative impacts to the environment and the most serious threat may be the resource competition with native organisms (Gab-Alla, 2007; Ndimele et al., 2011). Other commonly collected taxa, mostly on all substrates, included stonefly larvae (D. evansi), mayfly larvae (T. tillyardi), caddisfly larvae (hydroptilids) and midge larvae (Chironomidae). However, no significant differences and no correlation in relative abundance were detected.
In most cases, the abundance of the functional feeding groups was greater on willow roots treatment. Scrapers (primarily P. antipodarum) were the most dominant colonist on willow roots. This was probably caused by the physical heterogeneity and quantity of organic material provided by willow roots compared with the aluminium wire mesh. The high numbers of shredders (ephemeropterans, plecopterans and trichopterans) observed on willow roots suggest that most colonists are drawn from the drift and include those which disperse from low or high flows over short distances, by actively swimming or crawling over the substratum. In this study, ephemeropterans (Leptophlebiidae and Caenidae) are strong swimmers and crawlers, as are plecopterans such as D. evansi and I. mayii and were more abundant on willow root habitats. Predator species were also more abundant on willow roots, probably due to prey availability and refuge requirements. In contrast, other functional feeding groups such as collector-gatherers and collector-filterers, showed no consistent differences in abundance among substrate types. It might be expected that they would not be influenced by either structural heterogeneity or the presence of organic material but for most feeding groups, abundance was consistent with the prediction that invertebrate assemblages would be greater on the willow roots than on the aluminium wire mesh.
The hypothesis that habitat complexity would have a significant influence on the total abundance of aquatic invertebrate assemblages was supported (Fig. 1). Slightly more invertebrates were found on soft, fibrous willow roots especially where none of the willow roots were cut (WR3). When the structure of willow roots were less complex (WR2 and WR1), we found that the invertebrates were significantly less abundant. This lower abundance may be due to more homogeneous habitats as more fibrous fine and lateral willow roots were removed. These results suggest that willow roots are an adequate refuge for the invertebrates but major decreases in structural complexity results in a lower abundance of invertebrates.
The firm, smooth aluminium wire mesh substrates had significantly fewer aquatic invertebrates than willow roots. However, the attempt to mimic the structure of willow roots provides interesting findings into the effect of habitat complexity. Among the aluminium wire mesh substrates, invertebrates were more abundant on the fine mesh size (AS3) than on medium (AS2) and coarse (AS1) mesh sizes. Although more species and a greater diversity was also found on the fine mesh (AS3; 23 species), these differences were not significant. The complex habitat provided by aluminium wire mesh could support a diverse invertebrate fauna. It implies the fine aluminium wire mesh (AS3) may providing a better habitat compared with the other aluminium wire mesh treatments but further testing is necessary to confirm this. Not all invertebrate communities on the aluminium wire mesh had a high diversity and it is therefore not always possible to correlate total number of species with habitat complexity. The low diversity of invertebrates on medium (AS2) and coarse (AS1) aluminium mesh might indicate a poorer microhabitat for the invertebrates compared with willow roots.
There may be some mechanisms that might explain the differences observed in invertebrate abundance and species richness between artificial aluminium wire mesh and natural willow roots. The differences in total abundance and species richness were probably caused by several factors, such as physical heterogeneity of the substrata, quantity of organic material and biofilms covered on the artificial substrates. A study done by Moghaddasi et al. (2009) suggested that substrate seemed to be the most important environmental factors controlling the distribution of benthic foraminifera in the Northern Oman Sea. But, the high numbers of invertebrates on the willow roots was probably caused by greater surface area and physical characteristics of these substrata. However, more richness and diverse of invertebrates were found on fine size of aluminium wire mesh (AS3) indicating that artificial substrates may have greater impact on uncommon taxa. Indeed, a greater number of uncommon species such as the native lymnaied snails, baetid mayfly larvae (Offadens sp.), telephebiid dragonfly nymphs (Au. unicornis) and caenid amphipods (Austrochiltonia australis) were present on the fine size of aluminium wire mesh (AS3). Further research should focus on the biofilms which develop on them as probably, the presence of organic materials might be responsible to the occurrence of these uncommon species on fine aluminium wire mesh (AS3).
One-way cluster analysis showed that habitat complexity is probably the main factor in determining the distribution of invertebrates (Fig. 2). The importance of the effect of habitat complexity on the composition of the invertebrates is illustrated by the fact that all samples from less complex habitats (WR1, WR2, AS2 and AS1) fall into Cluster A and samples of high habitat complexity (WR3 and AS3) were grouped together in Cluster B. This indicate that substrate complexity is important, as more species were found in significantly greater densities on rough, fibrous and/or more complex substrates (WR3 and AS3) than smooth and less complex of substrates (WR1, WR2, AS2 and AS1). Four uncommon species were identified to be found strictly on more these complex habitats. They were the telephebiid dragonfly nymphs (Au. unicornis), caenid amphipods (Aus. australis), scirtid beetle (Cy. adelaidae) and dystiscid beetle (Platynectes decempunctatus).
Physical heterogeneity alone did not necessarily cause the differences we observed in the fauna between natural willow roots and aluminium wire mesh. Several observers have suggested that organic material affects colonisation by some of invertebrate taxa as it provides either a source of food or a suitable habitat (Suter, 1990; Casey and Kendall, 1996; Souter and Williams, 2001). The fine sediment we observed on willow roots probably included a natural biofilm and fine detritus that would provide food for some invertebrates especially the scrapers. Willows are an important food source for many aquatic invertebrates (Lester et al., 1994a) and Jayawardana et al. (2006) found more scrapers (Ph. acuta, P. antipodarum) and organic pollution-tolerant taxa (Cura sp., Megadrilii sp.) under willow roots, when willows had a higher Coarse Particulate Organic Matter (CPOM) content. We suspect that willow roots are attractive and may have similar nutritional value to willow leaves and possibly supply additional nutrition from increased microbial conditioning (Collier and Winterbourn, 1986), or perhaps from the removal of harmful secondary compounds which directly affect the invertebrate feeding (Lester et al., 1994b).
Differences in habitat complexity of willow roots may also influence the refuge selection behaviour of organisms. Levin et al. (2000) suggested that habitat may affect the frequency of interspecific or intraspecific behavioural interactions of the organisms. We suspect that habitat complexity may modify the effect on invertebrate functional feeding groups by providing refugia. We found more scrapers on fibrous, fine willow roots than on the aluminium wire mesh. Scrapers are probably colonist of willow roots, as willow roots provide a more stable and better habitat from high currents. The low number of other functional feeding groups may be due to competitive interactions, where there might be competition especially between the scrapers and collector-filterers for fine particulate organic matter on the willow roots. However, for the aluminium wire mesh, more shredders were found on fine (AS3) compared with medium (AS2) and coarse (AS1) mesh sizes. The higher abundance of invertebrates on more complex habitats (WR3 and AS3) might be related to the shelter provided from high currents. This is supported by the high abundance of some drift organisms such as D. evansi, L. aurata and hydroptilid caddisfly larvae which were highly associated with more complex habitats during high stream flows in spring.
Other functional feeding groups such as collector-gatherers (e.g., K. inconspicua and T. tillyardi) and collector-filterers (e.g., bivalves and chironomids) were greater in number on all willow root treatments, although no significant differences were detected. Predators such as chironomids and odonate nymphs were consistently associated with willow roots. Therefore, this study not only supports the hypothesis that habitat complexity is an important factor influencing the distribution of aquatic invertebrates but it also indicates the relatively large degree of change in the faunal composition and functional feeding groups in the stream.
There were significant increases in the abundance of aquatic invertebrates on the willow root treatments (WR3, WR2 and WR1) after 90 days of colonisation compared with 30 days of colonisation. Similar results were also found for the aluminium wire mesh substrates (AS3, AS2 and AS1) (Table 3). A long colonisation period is crucially important to obtain a stable invertebrates community because it allows development of biofilms and epilithics material which is known to affect colonization by aquatic invertebrates. A stable invertebrate community and colonisation dynamics is complex and generally not well understood (Rosenberg and Resh, 1993). Generally, short colonisation periods (<2 weeks) on artificial substrates cannot represent a stable community because the habitat will still be changing relatively rapidly. Casey and Kendall (1996) found that the macroinvertebrate assemblages colonising both artificial and natural substrate types were higher after 29 days of exposure. We found species composition was more diverse and abundant after 90 days than after 30 days of colonisation on all substrates. Some species such as baetid mayfly larvae (Offadens sp.), elmids beetle larvae (S. leai), conoesucid caddisfly larvae (Taschorema evansi) and tubificids were more abundant on complex habitats (WR2, WR3 and AS3) after 90 days of colonisation, probably due to the long exposure time and food source availability (e.g., biofilms and organic material).
We also found a more diverse and richer invertebrate fauna on fine aluminium wire mesh (AS3) after 90 days of colonisation (25 species), suggesting that aluminium wire mesh substrates may provide suitable habitats as well as food availability but that a longer period of colonisation is essential to obtain this richer and more diverse invertebrate community. The greatest abundance of invertebrates was found on WR3 after 90 days of colonisation (2397 individuals). However, habitat complexity and periods of colonisation did not influence invertebrate community assemblages. Consistent patterns in total abundances and species richness on each substrate type were found after each colonisation period. The classification of the samples obtained by one-way and two-way cluster analyses, is related to habitat complexity and days of colonisation. The results not only support the hypothesis that habitat complexity is an important factor influencing the distribution of aquatic invertebrates in a stream but also indicate that the period of colonisation may change the faunal composition (i.e., the longer the period, the better the colonisation).
For habitat complexity, the relative abundance of species was similar for all willow root treatments, except for the least complex willow roots (WR1) after 30 days of colonisation. This suggests that willow roots tend to be selectively colonised by certain species which may explain why the most complex willow roots (WR3) had a lower diversity compared with fine size of aluminium wire mesh (AS3). Generally, gastropods (primarily scrapers) constituted the greatest abundance in willow root treatments. The introduced hydrobiid snails, P. antipodarum, were significantly more abundant, especially on more complex habitat structures and over a longer period of colonisation (e.g., WR3 after 90 days). This may help explain why willow root treatments were grouped together. The rapid colonisation and utilisation of much or all of the available space by P. antipodarum on the willow roots had excluded other invertebrate species that might utilise the willow roots as a refuge.
The abundance of shredders and predators showed significant differences among the degree of habitat complexity and days of colonisation. Of the shredders, D. evansi, T. tillyardi and hydroptilid caddisflies were numerically dominant on all substrate types but their density was affected by the interaction between habitat complexity and days of colonisation. The only possible species preference noted was the colonisation of the telephebiid dragonfly nymphs (Au. unicornis) and scirtid beetle larvae (Cy. adelaidae) on aluminium wire mesh and their complete absence on the willow roots. Future research could concentrate on colonisation by individual species and their interactions with other species which might lead to a greater insight into colonisation patterns.
CONCLUSION
Overall, soft, fibrous willow roots have habitat value for aquatic invertebrates by providing a better surface texture and area and variety of microhabitats, for invertebrate colonisation. This study also indicates that artificial willow roots (fine aluminium wire mesh) can be used to stimulate invertebrate community and may be particularly useful especially in sites where willows have been removed. In the large scale removal of willows, especially in a small stream, at least some of the willow roots should be left in such an operation, as the sudden removal of their roots may disrupt the communities, including many native invertebrates which utilise the roots as habitat. As a relatively long period of colonisation is needed to obtain a complex and stable invertebrate community approaching what might be considered ‘normal’ and further examination of the amount of detritus and periphyton growth on willow roots and aluminium wire mesh should be carried out to decide which of these substrates is better for invertebrate colonisation during revegetation efforts. However, the most serious threat may be the resource competition between exotic P. antipodarum and native invertebrates.
ACKNOWLEDGMENTS
We thank Chris Madden (Freshwater Macroinvertebrates) for assistance during field work, species identification and valuable comments on the manuscript, Peter Goonan (Environment Protection Authority) for valuable advice and critiques on the manuscript, Katrina Warner (Sixth Creek Catchment Group) for helping with sites and field work, the Government of Malaysia for a scholarship to Wahizatul A. Azmi and the Nature Conservation Society of SA Inc. for a Conservation Biology Grant.
REFERENCES
- Davoudi, M.H., 2011. Influence of willow root density on shear resistance parameters in fine grain soils using in situ direct shear tests. Res. J. Environ. Sci., 5: 157-170.
CrossRefDirect Link - Moghaddasi, B., S.M.B. Nabavi, G. Vosoughi, S.M.R. Fatemi and S. Jamili, 2009. Abundance and distribution of benthic foraminifera in the Northern Oman Sea (Iranian side) continental shelf sediments. Res. J. Environ. Sci., 3: 210-217.
CrossRefDirect Link - Zorriasatein, N., B. Dehzad, G. Vossoughi, M. Shapoori and S. Jamili, 2009. Benthic fauna and water quality in Southern Caspian Sea Estuary: A case study on Gorganrood River. Res. J. Environ. Sci., 3: 599-603.
CrossRefDirect Link - Gab-Alla, A.A.F.A., 2007. Ecological study on community of exotic invasive seaweed Caulerpa prolifera in suez canal and its associated macro invertebrates. J. Applied Sci., 7: 679-686.
CrossRefDirect Link - Al-Sherif, E.A., W. Amer, S.E.A. Khodary and W. Azmy, 2009. Ecological studies on Salix distribution in Egypt. Asian J. Plant Sci., 8: 230-234.
CrossRefDirect Link - Anderson, M.J. and C.J.F.T. Braak, 2003. Permutation tests for multi-factorial analysis of variance. J. Stat. Comput. Simul., 73: 85-113.
Direct Link - Boulton, A.J. and P.I. Boon, 1991. A review of methodology used to measure leaf litter decomposition in lotic environments: Time to turn over an old leaf? Aust. J. Mar. Freshwater Res., 42: 1-43.
CrossRef - Bunn, S.E., P.M. Davies and T.D. Mosisch, 1999. Ecosystem measures of river health and their response to riparian and catchment clearing. Freshwater Biol., 41: 333-346.
CrossRef - Casey, R.J. and S.A. Kendall, 1996. Comparison among colonization of artificial substratum types and natural substratum by benthic macroinvertebrates. Hydrobiologia, 341: 57-64.
CrossRef - Collier, K.J. and M.J. Winterbourn, 1986. Processing of willow leaves in two suburban streams in Christchurch, New Zealand. N. Z. J. Mar. Freshwater Res., 20: 575-582.
CrossRef - Collier, K.J., 1995. Environmental factors affecting the taxonomic composition of aquatic macroinvertebrate communities in lowland waterways of Northland, New Zealand. N. Z. J. Mar. Freshwater Res., 29: 453-465.
CrossRef - Glova, G.J. and P.M. Sagar, 1994. Comparison of fish and macroinvertebrate standing stocks in relation to riparian willows (Salix spp.) in three New-Zealand streams. N. Z. J. Mar. Freshwater Res., 28: 255-266.
CrossRef - Jayawardana, C., M. Westbrooke, M. Wilson and C. Hurst, 2006. Macroinvertebrate communities in willow (Salix spp.) and reed beds (Phragmites australis) in central Victorian streams in Australia. Mar. Freshwater Res., 57: 429-439.
CrossRefDirect Link - Lester, P.J., S.F. Mitchell and D. Scott, 1994. Effects of riparian willow trees (Salix fragilis) on macroinvertebrate densities in two small Central Otago, New Zealand streams. N. Z. J. Mar. Freshwater Res., 28: 267-276.
Direct Link - Lester, P.J., S.F. Mitchell and D. Scott, 1994. Willow leaf and periphyton chemical-composition and the feeding preferences of Olinga feredayi (Trichoptera, Conoesucidae). N. Z. J. Mar. Freshwater Res., 28: 13-18.
CrossRef - Levin, P.S., N. Toimieri, M. Nicklin and P.F. Sale, 2000. Integrating individual behaviour and population ecology: the potential for habitat-dependent population regulation in a reef fish. Behav. Ecol., 11: 565-571.
CrossRef - Mackay, R.J., 1992. Colonization by lotic macroinvertebrates: A review of process and patterns. Can. J. Fishes Aquat. Sci., 49: 617-628.
Direct Link - Ndimele, P.E., C.A. Kumolu-Johnson and M.A. Anetekhai, 2011. The invasive aquatic macrophyte, water hyacinth {Eichhornia crassipes (Mart.) Solm-Laubach: Pontedericeae}: Problems and prospects. Res. J. Environ. Sci., (In Press).
Direct Link - Patten, D.T., 1998. Riparian ecosystems of semi-arid North America: diversity and human impacts. Wetlands, 18: 498-512.
CrossRef - Pusey, B.J., A.H. Arthington and M.G. Read, 1998. Freshwater fishes of the Burdekin River, Australia: Biogeography, history and spatial variation in assemblage structure. Environ. Biol. Fish., 53: 303-318.
CrossRef - Pusey, B.J., A.H. Arthington, J.R. Bird and P.G. Close, 2001. Reproduction in three species of rainbowfish (Melanotaeniidae) from rainforest streams in northern Queensland, Australia. Ecol. Freshwater Fish., 10: 75-87.
CrossRef - Schulze, D.J. and K.F. Walker, 1997. Riparian eucalypts and willows and their significance for aquatic invertebrates in the River Murray, South Australia. Regulated Rivers Res. Manage., 13: 557-577.
CrossRef - Souter, N.J. and W.D. Williams, 2001. A comparison of macroinvertebrate communities in three South Australian streams with regard to reintroduction of the platypus. Trans. Royal Soc. South Aust., 125: 71-82.
Direct Link - White, J. and T.C.R. White, 2005. Macro-invertebrates captured in artificial substrates in the restored water valley wetlands in South Australia. Trans. Royal Soc. South Aust., 129: 25-38.
Direct Link