
Imam A.A. Mekkawy
Department of Zoology, Faculty of Science, Assiut University, Assiut, Egypt
LiveDNA: 20.3952
U.M. Mahmoud
Department of Zoology, Faculty of Science, Assiut University, Assiut, Egypt
Sara Salah
Department of Zoology, Faculty of Science, Assiut University, Assiut, Egypt
American Journal of Biochemistry and Molecular Biology
Year: 2017 | Volume: 7 | Issue: 1 | Page No.: 21-40







ABSTRACT
Background and Objective: Atrazine is considered the most common aquatic herbicide used. So, the present study aimed at studying atrazine-induced changes in some biochemical characteristics during early embryonic stages of Clarias gariepinus . These characteristics include two metabolic enzymes, lactate hydrogenase (LDH) and glucose-6-phosphate dehydrogenase (G6PDH), two oxidative enzymes, glutathione reductase (GSH) and superoxide dismutase (SOD), two thyroid hormones, triiodothyronine (T3) and thyroxine (T4), lipid peroxidation (LPO), DNA damage and patterns of protein fractions of different early developmental stages. Material and Methods: Gametes were collected from mature specimens of the African catfish, C. gariepinus from the river Nile. The fertilized eggs were incubated in dechlorinized tap water (pH = 7.7, dissolved oxygen 88-94% saturation, temperature 27-29EC and photoperiod 12:12 (light:dark). Under atrazine influences, different measurements of enzyme activities, electrophoretic protein analysis, lipid peroxidation, DNA fragmentation, Total Protein (TP) and hormones were carried out. Results: The atrazine-induced significant fluctuation in the activity of G6PDH and LDH with a sharp decrease in 24 h-PFS after hatching. There is no significant variations in GSH activity under atrazine doses except in 24 and 96 h-PFS with a decrease in all developmental stages except at 72 h-PFS. Insignificant fluctuation in SOD activity in all developmental stages of C. gariepinus except 4, 20 and 24 h-PFS in comparison to control. Atrazine induced significant fluctuation in DNA damage and LPO/TP ratio in all developmental stages. Insignificant fluctuation in T3 activity but significant decrease in T4 activity in all developmental stages of C. gariepinus under exposure of sublethal doses of atrazine. Conclusion: One could conclude that atrazine significantly disturbe the metabolic parameters of embryonic and developmental stages of fishes and in turn most of these parameters could be used as indicators of environmental aquatic pollution.
PDF Abstract XML References Citation
Received: September 05, 2016;
Accepted: October 25, 2016;
Published: December 15, 2016
How to cite this article
Imam A.A. Mekkawy, U.M. Mahmoud and Sara Salah, 2017. Atrazine-induced Changes in some Biochemical Parameters of the Early Developmental Stages of the African Catfish Clarias gariepinus (Burchell, 1822). American Journal of Biochemistry and Molecular Biology, 7: 21-40.
DOI: 10.3923/ajbmb.2017.21.40
URL: https://scialert.net/abstract/?doi=ajbmb.2017.21.40
DOI: 10.3923/ajbmb.2017.21.40
URL: https://scialert.net/abstract/?doi=ajbmb.2017.21.40
INTRODUCTION
In Egypt, waste water and agriculture drains containing pesticides are discharged directly into rivers and lakes from industrial plans, aerial spraying of field and water drainage system1. These pesticides are stored under conditions that rarely meet internationally accepted standards2 leading to magnification of their contamination problem. Previous studies3-10 have reported that the exposure of fish to pesticides resulted in several changes many biochemical parameters including enzyme systems and hormones.
All chemical reactions in the cell are catalyzed by enzymes and controlled by hormones especially in embryonic and adult stages11,12. Introduction of foreign chemicals and pesticides in the cell generally disturb many enzyme and hormone functions13-20. Atrazine is one of the most common and significant pesticides that belongs to the s-triazine family of herbicides.
The natural antioxidant activity in animal body is a potential mechanism which contributes to the prevention of a variety of oxidative damages, toxicity and other diseases21-23. The antioxidant defense has become an important aspect of investigation in mammals and animals24,25. In this concern, fish are used as bio-indicators because the effects of exposure to xenobiotics can be measured in terms of biochemical or physiological responses26. Changes in age and species distribution in a fish population are also general indicators of water pollution but there are also responses specific to a single pollutant or a group of xenobiotics27.
African catfish Clarias gariepinus widely distributed throughout Africa28 has become one of the most important fishes used as bioindicator in terms of its biochemical and physiological characters9,28-36. Early developmental stages are considered to be one of the most sensitive stages in the fish life cycle to the toxic effects of chemical contaminants29,33,37-41.
Accordingly, the present study aimed at studying atrazine-induced changes in some biochemical characteristics during early embryonic stages of Clarias gariepinus to discover new discover environmental bioindicators for atrazine pollution . These characteristics include 2 metabolic enzymes, lactate hydrogenase and glucose-6-phosphate dehydrogenase, 2 oxidative enzymes, glutathione reductase and superoxide dismutase, 2 thyroid hormones, triiodothyronine (T3) and thyroxine (T4), lipid peroxidation, DNA damage and patterns of protein fractions of different early developmental stages.
MATERIALS AND METHODS
Gamete collection: Mature specimens of the African catfish, C. gariepinus (weight of 900-1500 g) were collected from the river Nile at Assuit, Egypt (during spawning seasons from April-July) and transported to the Fish Lab., Zoology Department, Assuit University.
The criteria applied for the selection of spawners were those described by De Graaf and Janssen42. For collection of semen, male fishes were anaesthetized with 200 mg L–1 Ms_222 (tricaine methane sulphonate, Crescent Research Chemicals, Phoenix, Arizona, USA) buffered with 800 mg L–1 sodium bicarbonate and one of the testes was removed surgically. Alternatively the fish were killed and the whole gonads were removed. Testes were cleaned from the blood by surgical towels. The sperms from the testes were pressed through a mesh fabric into a sterile dry petri dish and used directly for dry fertilization after ensuring the healthy and motility of sperms. For collection of eggs, ovulation was artificially induced. Females were injected intra-peritoneal with pellets (gonadotropin-releasing hormone analogue, GnRHa, D-Ala6, Pro9 Net) containing 2.5-3.0 mg of water soluble dopamine antagonist metoclopramide (Interfish Ltd., Hungary) dissolved in 0.65% NaCl. One pellet was used per kg body weight. After injection (10-11 h), the fish were stripped and the eggs were collectedin clean dry plastic containers, dry fertilization was considered.
Source of atrazine: A stock solution of herbicide atrazine (90% purity) provided by syngenta, Bazal, Switzerland (commercial name: Gesaprim 90 WG) was prepared by dissolving 1.10 g of the substance in 1 L of dechlorinated water.
Experimental setup and sampling: The fertilized eggs were incubated in dechlorinized tap water (pH = 7.7, dissolved oxygen 88-94% saturation, temperature 27-29°C and photoperiod 12:12 (light:dark). Exposure started 4 h after fertilization (4 h-PFS) then at intervals, 16, 24, 48, 72, 96 and 120 h-PFS for triiodothyronine (T3), thyroxine (T4), Glutathione Reductase (GR) and superoxide dismutase (SOD) in one experiment. The embryos were divided into 4 groups: One control and three concentrations 9, 18 and 27 mg L–1 of commercial herbicide atrazine. All working solutions were made immediately prior to use. Exposure took place in 12 petridishes (14 cm in diameter and 2.5 cm high), 3 petri dish replicates for each group (treated water level was 2 cm) for each developmental stage.
In another experiment, exposure started 4 h after fertilization then at intervals, 8, 12, 16 and 24 h-PFS for lactate dehydrogenase (LDH), glucose-6-phosphate dehydrogenase (G6PDH), lipid peroxidation (LPO) and DNA damage using three sublethal doses of atrazine (9, 18 and 27 mg L–1) in comparison to the control. The fertilized eggs were divided into eight groups: One control and seven concentrations 0.1, 1, 5, 10, 20, 100 and 200 mg L–1 of commercial herbicide atrazine. Exposure took place in 24 petri dishes (14 cm in diameter and 2.5 cm high), 3 petri dish replicates for each group (treated water level was 2 cm) for each developmental stage.
Measurements of enzyme activities: Because of the small size of the embryos, whole body homogenates were used for the measurements of the enzyme activities37. Homogenization of the whole animal is a reliable method that provides a reasonable index of total enzyme activity in individuals whose body mass is predominantly composed of muscle tissue43,44. The embryos were pulverized under liquid nitrogen and 100 mg of ground tissue powder was added to 2 volumes of buffer [50 mM tris, pH 7.4, 1 mM ethylene diamine tetra-acetic acid (EDTA) and 2 mM Mg Cl2]. Tissue was homogenized briefly with an Ultra-T URRAX (temperature was maintained at 4°C during homogenization). The homogenate was centrifuged for 15 min at 10,000xg and 4°C and the supernatant was used for the enzyme activity assay. Activities were determined in the supernatant with a spectrophotometer (Micro Lab 200 Vital Scientific, Dieren, The Netherlands) at a wavelength of 340 nm and at 37°C using kits, Stanbio LDH (UV-Rate) procedure No. 0940 USA for the quantitative determination of lactate dehydrogenase45 and RANDOX Laboratories Ltd., PD410, UK BT294QY for the quantitative determination of glucose-6-phosphate dehydrogenase46 G6PDH. The calculation of the catalytic activity content for the selected enzymes was in accordance with the recommendations of the French Society of Clinical Biology47. The principle of the assay consists of the kinetic determination of LDH activity, based on the rate of NADH oxidation. We determined the oxidation rate, which is directly proportional to LDH activity by measuring the decrease in absorbance at 340 nm. To standardize the expressions of enzyme activity, the units of enzyme activity have been defined as the quantity of enzyme that catalyzes the reaction of 1 mmol of substrate min–1. The catalytic concentration was expressed as U L–1.
In addition to determine the antioxidant enzymes at wavelength 340 nm and 37°C using kits, biodiagnostic (UV method) for the determination of glutathione reductase48 which is essential for the glutathione redox cycle that maintains adequate levels of reduced cellular GSH. The GSH serves as an antioxidant, reacting with free radicals and organic peroxides, in amino acid transport and as asubstrate for the glutathione peroxidases and glutathione S-transferases in the detoxification of organic peroxide and metabolism of xenobiotics,respectively. And at wavelength 560 nm for the determination of superoxide dismutase49 which is metalloenzyme that catalyze the dismutation of the superoxide anion to molecular oxygen and hydrogen peroxide and thus form acrucial part of the cellular antioxidant defense mechanism.
Protein analysis by polyacrylamide gel electrophoresis (SDS–PAGE): Embryos (~0.1 g fresh weight) of each treatment in addition to control were suspended in 1.0 mL lysing buffer, heated at 100°C for 5 min centrifuged at 10,000 rpm for 30 min and 50 μL of each extracted protein treatment was used for protein analysis using SDS-PAGE according to Laemmli50 in the first dimension. The low-molecular weight standards (Pierce, USA) were run concurrently and the protein molecular mass was determined using Gel-Pro analyzer package version 3.1 for Windows XP/NT (Media Cybernetica 1993-97).
Lipid peroxidation and total protein measurements: Total protein contents were determined according to the Biuret method51 using bovine serum albumin (E. Merck-Darmstadt, Germany) as a standard. Lipid peroxidation (LPO) in the embryos was determined by the procedure of Utley et al.52. The absorbance of each aliquot was measured at 535 nm. The rate of lipid peroxidation was expressed as nmol of thiobarbituric acid reactive substance (TBARS) formed per h per milligram of protein using a molar extinction coefficient53 of 1.56 M–1 cm–1.
DNA fragmentation measurement: The DNA fragmentation was determined by the procedure of Kurita-Ochiai et al.54 using spectrophotometer (Micro lab 200 vital scientific Dieren the Netherlands) at 575 or 600 nm against reagent blank. The percentage of fragmented DNA was estimated by the following formula:
Hormones measurements: According to the method of Cobb et al.55, the triiodothyronine (T3) and total thyroxine (T4) in embryos after homogenated in phosphate buffer saline were measured according to the method of Skelley et al.56 using ELISA kits (with Cat# BC-1005 and BC-1007 for T3 and T4, respectively). These kits were obtained from BioCheck, Inc. 323 Vintage Park Dr., Foster city, CA94404.
Statistical analysis: The basic statistics, means, standard errors and ranges were estimated. The patterns of variation due to developmental stages and atrazine doses and their interaction were studied by two way analysis of variance using the SPSS package57 at the 0.05 significance level. Levene’s test of equality of error variance of the dependent variables was applied, with rejection of the null hypothesis for raw, log-transformed and SQRT-transformed data. So, the homogeneity of variance was assumed for raw data. The Tukey-HSD test was considered for multiple comparisons. Moreover, the Dunnett, t-test was applied, measuring the control against other treatments in each developmental stage.
RESULTS
Metabolic enzymes: G6PDH and LDH: The atrazine doses have different adverse impacts on 2 metabolic enzymes (G6PDH and LDH) during early embryonic development of Clarias gariepinus. The atrazine-induced activity of G6PDH (Table 1, Fig. 1) was fluctuated significantly with developmental progress (p<0.001) with a peak in 12 h-PFS and a sharp decrease in 24 h-PFS directly after hatching. Such sharp decrease may refer to the fact that zygotic gene of G6PDH is activated during this later stage. Similarly, significant fluctuation (p<0.001) in G6PDH activity was evident under atrazine doses with no special trends towards decrease or increase in each developmental stage studied.
The atrazine-induced activity of LDH (Table 2, Fig. 2) was fluctuated significantly in some cases and increased in others with developmental progress (p<0.001) with a sharp decrease in 24 h-PFS directly after hatching. Such sharp decrease may refer to the fact that zygotic gene of LDH is activated during this later stage.
| Table 1: | Effect of different doses of atrazine on the activity of G6PDH (Mean±SE) during early developmental stages of the African catfish Clarias gariepinus |
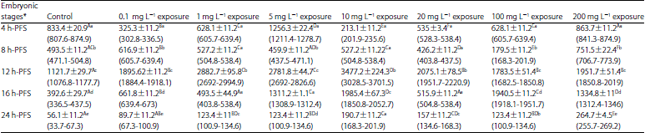 | |
Values are in units per litter, *Embryonic stages showing similar lower case letters are insignificant within the doses at 0.05 level (vertical comparison), stages showing similar capital letters are insignificant within the embryonic stages at 0.05 levels (horizontal comparison), hatching process started 20 h after fertilization | |
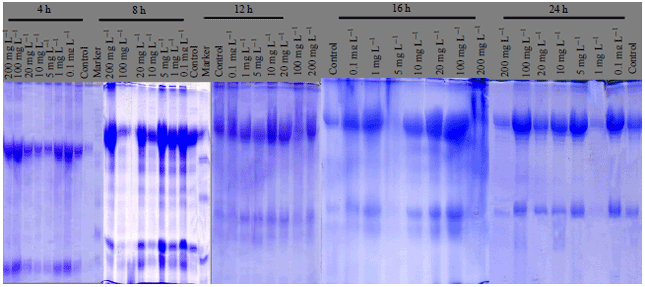 |
| Fig. 1: | Protein fractions identified in different larval stages of C. gariepinus under different doses of atrazine in comparison with control *h-PFS: Hours post fertilization stages |
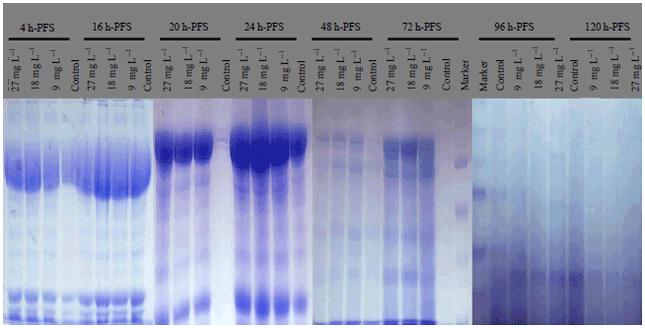 |
| Fig. 2: | Protein fractions identified in different larval stages of C. gariepinus under different doses of atrazine in comparison with control, h-PFS: Hours post fertilization stages |
| Table 2: | Effect of different doses of atrazine on the activity of LDH (Mean±SE) during early developmental stages of the African catfish Clarias gariepinus |
 | |
Values are in units per litter, *Embryonic stages showing similar lower case letters are insignificant within the doses at 0.05 level (vertical comparison), stages showing similar capital letters are insignificant within the embryonic stages at 0.05 levels (horizontal comparison), hatching process started 20 h after fertilization | |
Similarly, significant fluctuation (p<0.001) in LDH activity was evident under atrazine doses with no special trends towards decrease or increase in each developmental stage studied. However, LDH activity increased in higher atrazine doses and in old stages studies (Table 2) till 24 h-PFS.
Oxidative enzymes: GR and SOD: In each of the developmental stages studies (4-120 h-PFS) of C. gariepinus, there is no significant variations in GR activity under atrazine sublethal doses except in 24 and 96 h-PFS stages (Table 3). Such variations (p>0.05) were directed towards decrease in all developmental stages except 72 h-PFS which show fluctuation in enzyme activity under stress. Except for the control group (p<0.05), the GR activity exhibited insignificant variations among developmental stages.
Atrazine-induced SOD activity insignificantly (p>0.05) fluctuated in all developmental stages of C. gariepinus except 4, 20 and 24 h-PFS in comparison to the control. Except for the control and 27 mg L–1 groups (p<0.05), the SOD activity exhibited insignificant variations among developmental stages under the stress of other sublethal doses (Table 4).
DNA damage: Atrazine-induced DNA damage significantly (p<0.05) fluctuated in all developmental stages of C. gariepinus except 4 h-PFS in comparison to the control. Except for the control, 0.1 and 10 mg L–1 groups (p>0.05), the DNA damage exhibited significant variations among developmental stages under the stress of other sublethal and lethal doses (Table 5) (p<0.0001). Significant interaction between atrazine doses and embryonic stages was evident.
Lipid peroxidation relative to total protein (LPO/TP): Atrazine-induced LPO/TP significantly (p<0.0001) fluctuated in all developmental stages of C. gariepinus. The LPO/TP exhibited also significant variations among developmental stages under the stress of sublethal and lethal doses (Table 6) (p<0.0001). Significant interaction between atrazine doses and embryonic stages was evident.
| Table 3: | Effect of three doses of atrazine on the activity of glutathione reductase (U L–1) (Mean±SE) during early developmental stages of the African catfish Clarias gariepinus |
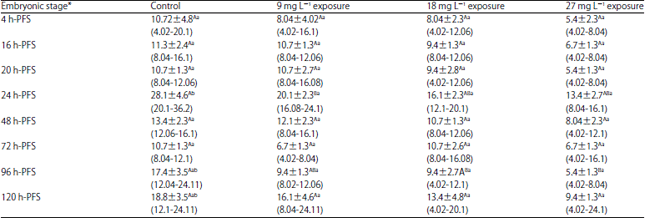 | |
*Embryonic stages showing similar lower case letters are insignificant within the doses at 0.05 level (vertical comparison), stages showing similar capital letters are insignificant within the embryonic stages at 0.05 levels (horizontal comparison), hatching process started 23 h after fertilization | |
| Table 4: | Effect of three doses of atrazine on the activity of Superoxide dismutase (U mg–1 tissue), (Mean±SE) during early developmental stages of the African catfish Clarias gariepinus |
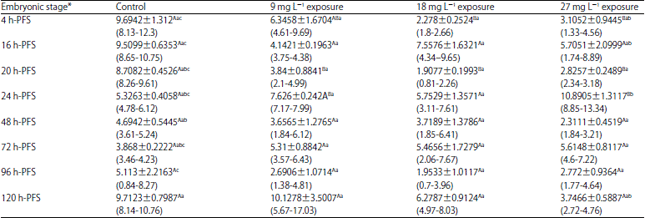 | |
*Embryonic stages showing similar lower case letters are insignificant within the doses at 0.05 level (vertical comparison), stages showing similar capital letters are insignificant within the embryonic stages at 0.05 levels (horizontal comparison), hatching process started 23 h after fertilization | |
| Table 5: | Effect of different doses of atrazine on the percentage of DNA damage (%) (Mean±SD) during early developmental stages of the African catfish Clarias gariepinus |
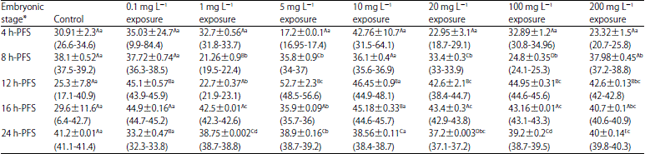 | |
*Embryonic stages showing similar lower case letters are insignificant within the doses at 0.05 level (vertical comparison), stages showing similar capital letters are insignificant within the embryonic stages at 0.05 levels (horizontal comparison), hatching process started 20 h after fertilization | |
Thyroid hormones: T3 and T4: Atrazine-induced T3 variations insignificantly (p>0.05) fluctuated in all developmental stages of C. gariepinus. Except for the control, 0.1 and 27 mg L–1 groups (p<0.0001), T3 activity exhibited insignificant variations among developmental stages under the stress of other sublethal doses (Table 7) (p>0.05).
| Table 6: | Effect of different doses of atrazine on the lipid peroxidation/total protein ratio (Mean±SE) (nmol mg–1) during early developmental stages of the African catfish Clarias gariepinus |
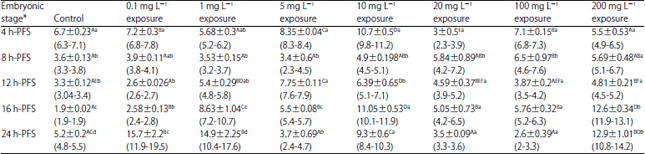 | |
*Embryonic stages showing similar lower case letters are insignificant within the doses at 0.05 level (vertical comparison), stages showing similar capital letters are insignificant within the embryonic stages at 0.05 levels (horizontal comparison), the hatching process started 23 h after fertilization | |
| Table 7: | Effect of three doses of atrazine on the activity of triiodothyronine (T3) (ng mL–1), (Mean±SE) during early developmental stages of the African catfish Clarias gariepinus |
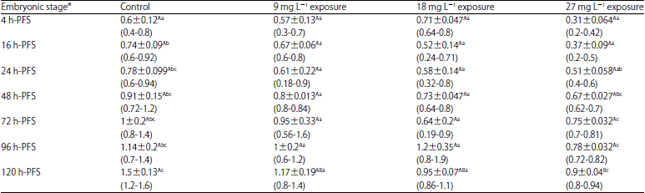 | |
*Embryonic stages showing similar lower case letters are insignificant within the doses at 0.05 level (vertical comparison), stages showing similar capital letters are insignificant within the embryonic stages at 0.05 levels (horizontal comparison), hatching process started 23 h after fertilization | |
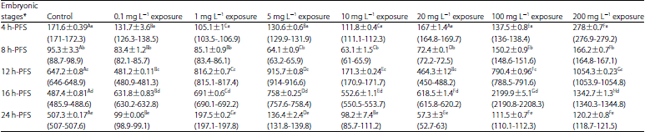
| Table 8: | Effect of three doses of atrazine on the activity of thyroxine (T4) (µg dL–1), (Mean±SE) during early developmental stages of the African catfish Clarias gariepinus |
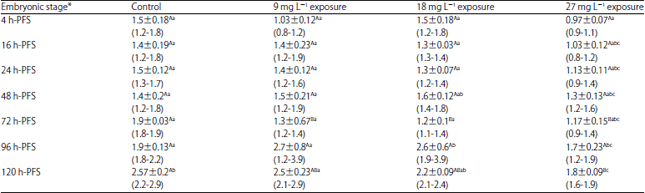 | |
*Embryonic stages showing similar lower case letters are insignificant within the doses at 0.05 level (vertical comparison), stages showing similar capital letters are insignificant within the embryonic stages at 0.05 levels (horizontal comparison), hatching process started 23 h after fertilization | |
No significant impacts of atrazine sublethal doses on the activity of T3. The variations in the T3 activity may be present in the treated group as well as the control one. Only in the older early embryonic stages (72 and 120 h-PFS), the atrazine sublethal doses led to significant decreased change in T4 activity in comparison to the control (Table 8). Except for 9 mg L–1, within each treatment, T4 activity exhibited significant variations among developmental stages. Such variations may not be related to atrazine doses but to the stage of development.
Protein electrophoresis: The protein fractions of different developmental (4, 8, 12, 16, 20, 24, 48, 72, 96 and 120 h) stages used in the present study were identified in terms of their molecular weight and concentration under atrazine (0.1, 1, 5, 9, 10, 18, 20, 27, 100 and 200 mg L–1 ) stress in comparison with the control. Variability in these 2 parameters and number of bands were evident reflecting variations in protein gene expression under atrazine stress, developmental stages and their interactions (Table 9-12).
| Table 9: | Protein fractions (in percent ) identified in different larval stages of C. gariepinus under different doses of atrazine in comparison with control |
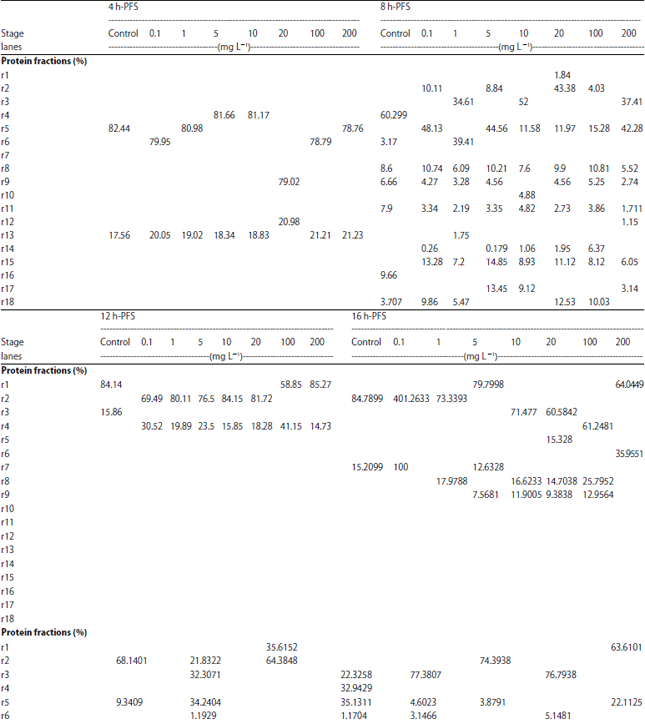  | |
| h-PFS: Hours post fertilization stages | |
| Table 10: | Molecular weight (in kda) identification of protein fractions in different larval stages of C. gariepinus under different doses of atrazine in comparison with control |
 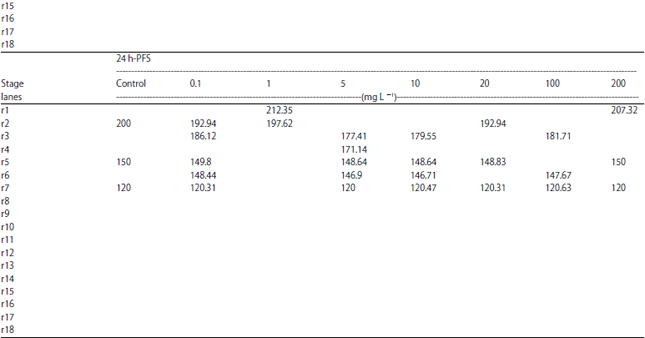 | |
h-PFS: Hours post fertilization stages | |
| Table 11: | Protein fractions (in percent) identified in developmental stages of C. gariepinus under effect of different doses atrazine in comparison with control |
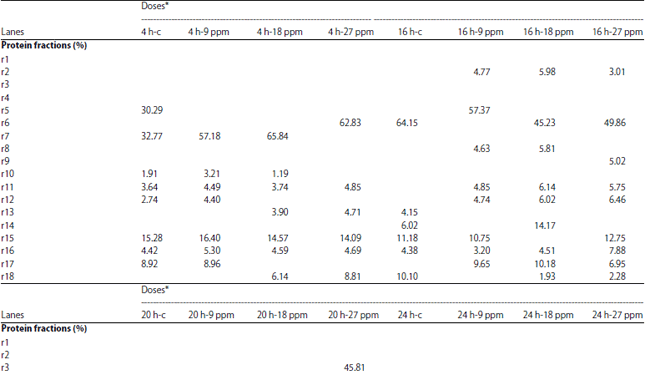 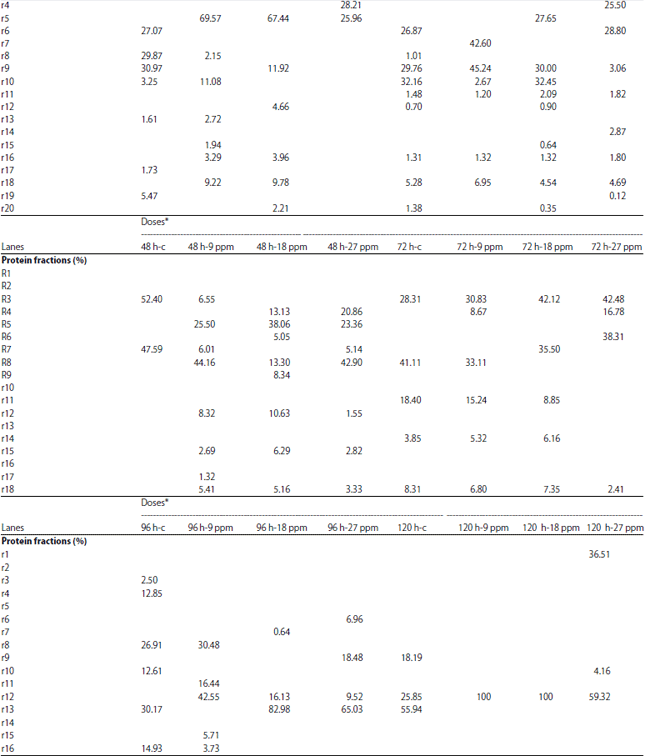 | |
h-PFS: Hours post fertilization stages | |
| Table 12: | Molecular weight (in kda) identification of protein fractions in developmental stages of C. gariepinus under effect of different doses of atrazine in comparison with control |
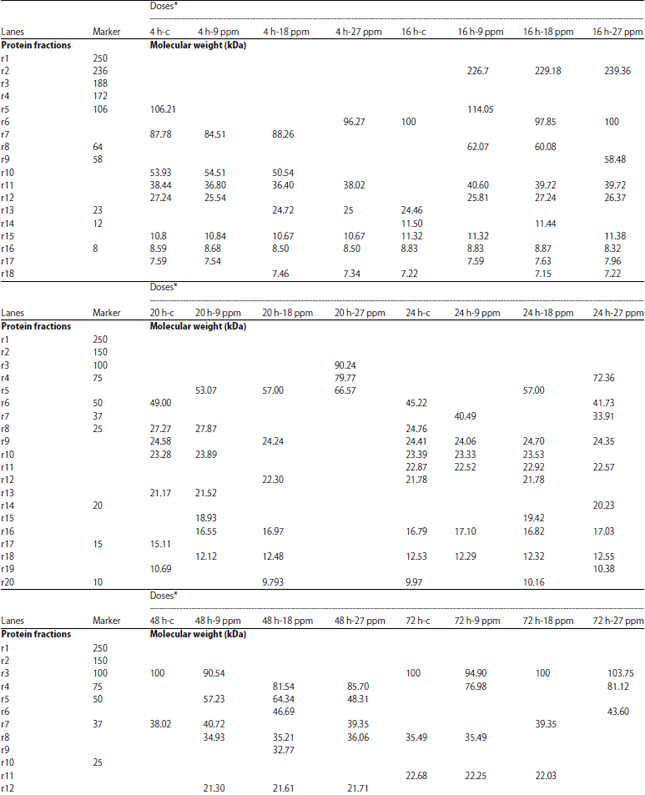 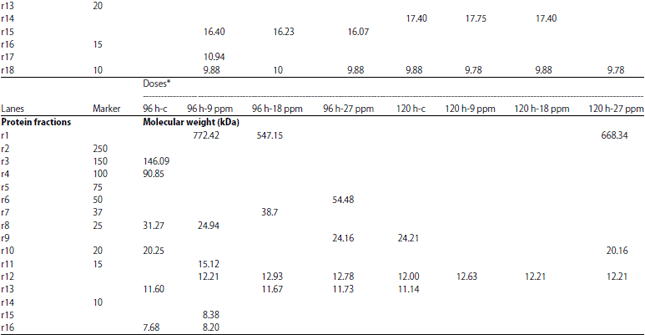 | |
| *h-PFS: Hours post fertilization | |
DISCUSSION
The contamination of aquatic ecosystems by xenobiotics has gained increasing attention58-62 and several recent studies have demonstrated the toxicity and effects to fish under field and laboratory conditions4,63-69. Atrazine is one of the most widely used herbicides and because of its considerable persistence and mobility in soil and water, it is considered a common terrestrial and aquatic contaminant70. Atrazine treatment increased hydrogen peroxide (H2O2) content, while reducing gene expression and enzymatic activities related to two major H2O2-detoxification pathways71.
The uptake kinetic of atrazine exhibited a three-compartment characteristic. It had already reached the embryo within 10s of exposure time and is rapidly absorbed in the first 6 h, then followed steady state conditions between 6 and 24 h. Atrazine uptake is also influenced by elimination process since such process was more effective between 24 and 48 h72. In the present study, such pattern of atrazine uptake was evident by its adverse fluctuated effect on different biochemical parameters considered.
The activities of enzymes such as LDH and G6PDH have been used as bioindicator of stress in catfish Clarias garepinus9,29,32,33,73. The variations in these two metabolic enzyme systems of the embryonic stages of Ctenopharyngodon idellus and Clarias gariepinus were studied by Mekkawy and Lashein40, Mekkawy et al.74 and Osman et al.33. These researchers discussed the developmental time of activation and inactivation of the maternal and zygotic genes through those early developmental stages. The switching time (i.e., blastula, gastrula and organogenesis) from maternal to zygotic LDH and G6PDH-genes was determined in the control and stressed developmental stages. The pattern of LDH and G6PDH activities in the present study was also determined directly before 24 h-PFS. Interaction between atrazine doses and developmental stages was evident with fluctuating variations in the enzyme activities. Decrease in G6PDH activities under pesticide stress is well documented in literature75,76. Both the decreased concentration of G6PDH and the increased activity of LDH can be explained as a consequence of stress-based especially hepatic tissue77. The SOD and GSH provide the first line of defense against oxygen derived free radicals4,78-80. Different studies deals with these enzymes under different types of stress81-86 especially herbicides3-5 in fishes. Increase in the SOD activities is probably a response towards increased ROS generation in pesticide toxicity87. Generally exposure of animals to xenobiotics increases SOD activity in various tissues87,88. The St. Dimitrova et al.89 reported a simultaneous induction response in SOD activities in carp Cyprinus carpio exposed to zinc. Different researchers referred to increase90-92 or decrease76,93-96 in GSH activities under stress in fishes. Fluctuations GSH and SOD activities under atrazine stress was evident in most developmental stages with interaction between atrazine doses and progress in early development. The pesticide toxicity results from the metabolic conversion processed in the endoplasmic reticulum of cells in concern directly affecting the morphology of these cells97,98. Oxidative stress may be due to the depletion of GSH which prevents the conjugation of xenobiotics like atrazine to GSH and thus enables them to freely combined covalently with cell proteins99. Jayachandran et al.90 reported that the reduced levels of GSH in the atrazine administered Labeo rohita could be the result of either decreased utilization of GSH for induced toxicity. Envionmental stress has been demonstrated to cause an increase in the oxidative stress, an imbalance in the antioxidant status100. Nammalwar101 reported that the change in glutathione redox ratio and hence a decrease in reduced GSH levels, the glutathione-mediated detoxification process may also be affected. This might be a factor responsible for the lack of elimination of toxic compounds that enter the fish and thus result in their accumulation, aggravating oxidative stress. Yildirim et al.78 concluded that a decline in tissue GSH content during exposure to pollution may be due to an increased utilization of GSH, which can be converted into oxidized glutathione and in-efficient GSH regeneration. However, severe oxidative stress may suppress GSH levels due to the impairment of adaptive mechanisms102.
Exposure of fishes to pesticides and other xenobiotics is known to induce lipid peroxidation in various tissues especially liver ones, which is responsible for the adverse biological effects103-105. Previous investigations have reported the induction of LPO by pesticides such as deltamethrin106,107, alachlor108, malathion109 and butachlor5 in fish. Parthasarathy and Joseph93 demonstrated that λ-cyhalothrin as a type of pesticides might have stimulated lipid peroxidation by influencing a variety of reactions in freshwater tilapia Oreochromis mossambicus. Nwani et al.4 also reported that the elevated level of lipid peroxidation in the liver of Channa punctatus in response to the exposure to atrazine resulting from reactive oxygen species which may lead to cell apoptosis.
The degree of lipid peroxidation is associated with toxic doses of chemicals, time of exposure, species, developmental stages and their interactions4,10,33,40,74. Most of these findings were evident in the present study by Atrazine-induced changes in LPO in early embryonic stages of C. gariepinus. Singh et al.110 reported that reactive oxygen species, such as superoxide anions, hydroxyl radical and hydrogen peroxide enhance the oxidative process and induce peroxidative damage to membrane lipids. So, the fluctuation in current pattern of LPO may be one of the molecular and adaptation mechanisms involved in the atrazine-induced toxicity on early embryonic stages.
Different investigations emphasized on DNA damage under chemical stresses in fishes29,33. Many of these investigations have shown that ROS is the major source of DNA damage by causing strand breaks, removal of nucleotides and various modifications of the nucleotide bases111. Variability of DNA damage were found to be related to species, age, chemical doses and time of their exposure and their interactions. In the present study, pattern of variations in DNA damage was evident in concern with the early embryonic stages of C. gariepinus and atrazine doses.
The sensitivity of thyroid gland to many toxicants have gained much attention and a number of endocrine disrupting compounds have been30,112. The thyroid hormones are known for their osmoregulatory and metabolic effects113-115 and are involved in the regulatory act of stress tolerance in fish116-118. These hormones also control development, growth and reproduction of fishes113,117,119-128. It is possible that the elevated plasma levels of thyroid hormones in broodstock during reproduction are necessary to ensure an adequate concentration of hormones in the developing oocyte127,129. Moreover, several studies have demonstrated the beneficial effects of increased levels of maternal T3 on subsequent larval development and survival, e.g., striped sea bass and the rabbitfish Siganus guttatus128,130. In many species studied, there is a clear evidence that the concentration of T4 and T3 decreases as embryogenesis proceeds. The biological significance of such evidence is still unclear131,132.
Different pesticides have been found to change the pattern of secretion of thyroid hormones in adult and embryonic stages of fishes30,133,134. Exposure of catfish Heteropneutes fossilis and Clarias batrachus to malathion and endosulfan caused changes in circulatory thyroid hormones133,134. A decrease in T3 has been reported in rainbow trout exposed to acidic water135 and starvation136-140. A change in T3 and T4 hormones have been reported in adult catfish Clarias gariepinus exposed to 4-nonylphenol41. In the present study, fluctuations in T3 and T4 activities in the developmental stages of C. gariepinus were insignificant in most cases and referred to the fact that it may not be related to atrazine doses but to the stage of development.
In the present study, the Variability in the molecular weight, concentration and number of bands were evident reflecting variations in protein gene expression under atrazine stress, developmental stages of C. gariepinus and their interactions. Our results are consistent with those of Mekkawy et al.29 who found that the protein fractions in fish embryos exposed to UV-A have fluctuations in their protein concentration and their molecular weight in comparison with control embryos.
CONCLUSION
It is concluded that atrazine induces oxidative stress, DNA damage, lipid peroxidation and endocrine disruption on the developmental stages of African catfish Clarias gariepinus. Most of these parameters could be used as indicators of environmental aquatic pollution. However, fluctuations in T3 and T4 activities in the developmental stages of C. gariepinus were insignificant in most cases and referred to the fact that it may not be related to atrazine doses but to the stage of development.
SIGNIFICANCE STATEMENT
Discovering new indicators to assess environmental risks is an important issue especially with the increase of use of herbicides like atrazine. So this study and the related others are a must especially in concern with hormone disruptors and genotoxicity. The sensitivity of such situations is related to the chemicals as well as fish species and the aquatic communities.
REFERENCES
- Qian, H., W. Chen, G.D. Sheng, X. Xu, W. Liu and Z. Fu, 2008. Effects of glufosinate on antioxidant enzymes, subcellular structure and gene expression in the unicellular green alga Chlorella vulgaris. Aquat. Toxicol., 88: 301-307.
CrossRefDirect Link - Nwani, C.D., W.S. Lakra, N.S. Nagpure, R. Kumar, B. Kushwaha and S.K. Srivastava, 2010. Toxicity of the herbicide atrazine: Effects on lipid peroxidation and activities of antioxidant enzymes in the freshwater fish Channa punctatus (Bloch). Int. J. Environ. Res. Public Health, 7: 3298-3312.
CrossRefDirect Link - Farombi, E.O., Y.R. Ajimoko and O.A. Adelowo, 2008. Effect of Butachlor on antioxidant enzyme status and lipid peroxidation in fresh water African Catfish, (Clarias gariepinus). Int. J. Environ. Res. Public Health, 5: 423-427.
CrossRefPubMedDirect Link - Samprath, K., S. Velamnial, I.J. Kennedy and R. James, 1993. Haematological changes and their recovery in Oreochromis mossambicus as a function of exposure period and sublethal levels of Ekalux. Acta Hydrobiol., 35: 73-83.
Direct Link - Adedeji, O.B., O.K. Adeyemo and S.A. Agbede, 2009. Effects of diazinon on blood parameters in the African catfish (Clarias gariepinus). Afr. J. Biotechnol., 8: 3940-3946.
Direct Link - Paskova, V., K. Hilscherova and L. Blaha, 2011. Teratogenicity and embryotoxicity in aquatic organisms after pesticide exposure and the role of oxidative stress. Rev. Environ. Contamin. Toxicol., 211: 25-61.
CrossRefDirect Link - Devlin, R.H. and Y. Nagahama, 2002. Sex determination and sex differentiation in fish: An overview of genetic, physiological and environmental influences. Aquaculture, 208: 191-364.
CrossRefDirect Link - Almeida, J.A., E.L.B. Novelli, M. Dal pPai Silva and R. Alves Jr., 2001. Environmental cadmium exposure and metabolic responses of the Nile tilapia, Oreochromis niloticus. Environ. Pollut., 114: 169-175.
CrossRefDirect Link - Tongo, I., L. Ezemonye and U. Ochei, 2012. Diazinon mediated biochemical changes in the African toad (Bufo regularis). J. Xenobiot., 2: 18-23.
CrossRefDirect Link - Manning, T., 2005. Endocrine-disrupting chemicals: A review of the state of the science. Aust. J. Ecotoxicol., 11: 1-52.
Direct Link - Luskova, V., M. Svoboda and J. Kolarova, 2002. The effect of diazinon on blood plasma biochemistry in carp (Cyprinus carpio L.). Acta Veterinaria Brno, 71: 117-123.
Direct Link - Zaki, M.S., O.M. Fawzi, S.O. Mostafa, I. Awad and M. Fawzy, 2010. Biochemical studies on Tilipia nilotica exposed to climate change and cadmium sulphate (0.50p.p.m.). N. Y. Sci. J., 3: 90-95.
Direct Link - Sobha, K., A. Poornima, P. Harini and K. Veeraiah, 2007. A study on biochemical changes in the fresh water fish, Catla catla (Hamilton) exposed to the heavy metal toxicant cadmium chloride. Kathmandu Univ. J. Sci. Eng. Technol., 1: 1-11.
CrossRefDirect Link - Uttara, B., A.V. Singh, P. Zamboni and R.T. Mahajan, 2009. Oxidative stress and neurodegenerative diseases: A review of upstream and downstream antioxidant therapeutic options. Curr. Neuropharmacol., 7: 65-74.
CrossRefPubMedDirect Link - Fang, Y.Z., S. Yang and G. Wu, 2002. Free radicals, antioxidants, and nutrition. Nutrition, 18: 872-879.
CrossRefDirect Link - Hsu, C.H., Z.H. Wen, C.S. Lin and C. Chakraborty, 2007. The zebrafish model: Use in studying cellular mechanisms for a spectrum of clinical disease entities. Curr. Neurovasc. Res., 4: 111-120.
CrossRefDirect Link - Garcia-Alonso, F.J., A. Guidarelli and M.J. Periago, 2007. Phenolic-rich juice prevents DNA single-strand breakage and cytotoxicity caused by tert-butylhydroperoxide in U937 cells: The role of iron chelation. J. Nutr. Biochem., 18: 457-466.
CrossRefDirect Link - Wang, C., Y. Zhao, R. Zheng, X. Ding, W. Wei, Z. Zuo and Y. Chen, 2006. Effects of tributyltin, benzo[a]pyrene and their mixture on antioxidant defense systems in Sebastiscus marmoratus. Ecotoxicol. Environ. Saf., 65: 381-387.
CrossRefPubMedDirect Link - Mondon, J.A., S. Duda and B.F. Nowak, 2001. Histological, growth and 7-ethoxyresorufin O-deethylase (EROD) activity responses of greenback flounder Rhombosolea tapirina to contaminated marine sediment and diet. Aquat. Toxicol., 54: 231-247.
CrossRefDirect Link - Siroka, B., J. Huttova, L. Tamas, M. Simonovicova and I. Mistrik, 2004. Effect of cadmium on hydrolytic enzymes in maize root and coleoptile. Biologia, 59: 513-517.
Direct Link - Nguyen, L.T.H. and C.R. Janssen, 2002. Embryo-larval toxicity tests with the African catfish (Clarias gariepinus): Comparative sensitivity of endpoints. Arch. Environ. Contam. Toxicol., 42: 256-262.
CrossRefDirect Link - Mekkawy, I.A.A., U.M. Mahmoud, A.G. Osman and A.E.D.H. Sayed, 2010. Effects of ultraviolet A on the activity of two metabolic enzymes, DNA damage and lipid peroxidation during early developmental stages of the African catfish, Clarias gariepinus (Burchell, 1822). Fish physiol. Biochem., 36: 605-626.
CrossRefPubMedDirect Link - Sayed, A.E.D.H., U.M. Mahmoud and I.A. Mekkawy, 2012. Reproductive biomarkers to identify endocrine disruption in Clarias gariepinus exposed to 4-nonylphenol. Ecotoxicol. Environ. Saf., 78: 310-319.
CrossRefPubMedDirect Link - Sayed, A.E.D.H., I.A.A. Mekkawy and U.M. Mahmoud, 2011. Effects of 4-nonylphenol on metabolic enzymes, some ions and biochemical blood parameters of the African catfish Clarias gariepinus (Burchell, 1822). Afr. J. Biochem. Res., 5: 287-297.
Direct Link - Sayed, A.E.D.H., A.T. Ibrahim, I.A.A. Mekkawy and U.M. Mahmoud, 2007. Acute effects of ultraviolet-a radiation on African catfish Clarias gariepinus (Burchell, 1822). J. Photochem. Photobiol. B: Biol., 89: 170-174.
CrossRefPubMedDirect Link - Osman, A.G.M., I.A. Mekkawy, J. Verreth and F. Kirschbaum, 2007. Effects of lead nitrate on the activity of metabolic enzymes during early developmental stages of the African catfish, Clarias gariepinus (Burchell, 1822). Fish Physiol. Biochem., 33: 1-13.
CrossRefDirect Link - Richter, C.J.J., W.J.A.R. Viveen, E.H. Eding, M. Sukkel and A.J. Rothuis et al., 1987. The significance of photoperiodicity, water temperature and an inherent endogenous rhythm for the production of viable eggs by the African catfish, Clarias gariepinus, kept in subtropical ponds in Israel and under Israeli and Dutch hatchery conditions. Aquaculture, 63: 169-185.
CrossRefDirect Link - Osman, A.G.M., M. Koutb and A.E.D.H. Sayed, 2010. Use of hematological parameters to assess the efficiency of quince (Cydonia oblonga Miller) leaf extract in alleviation of the effect of ultraviolet-A radiation on African catfish Clarias gariepinus (Burchell, 1822). J. Photochem. Photobiol. B: Biol., 99: 1-8.
CrossRefPubMedDirect Link - Peter, M.C.S., J. Leji, V. Rejitha, J. Ignatius and V.S. Peter, 2008. Physiological responses of African catfish (Clarias gariepinus) to water-borne ferric iron: Effects on thyroidal, metabolic and hydromineral regulations. J. Endocrinol. Reprod., 12: 24-30.
Direct Link - Osman, A.G.M., S. Wuertz, I.A. Mekkawy, H.J. Exner and F. Kirschbaum, 2007. Lead induced malformations in embryos of the African catfish Clarias gariepinus (Burchell, 1822). Environ. Toxicol., 22: 375-389.
CrossRefPubMedDirect Link - Weis, J.S. and P. Weis, 1987. Pollutants as developmental toxicants in aquatic organisms. Environ. Health Perspect., 71: 77-85.
CrossRefDirect Link - Mahmoud, U.M., I.A.A. Mekkawy and A.E.D.H. Sayed, 2009. Ultraviolet radiation-A (366 Nm) induced morphological and histological malformations during embryogenesis of Clarias gariepinus (Burchell, 1822). J. Photochem. Photobiol. B: Biol., 95: 117-128.
CrossRefPubMedDirect Link - Mekkawy, I.A.A. and F.E. Lashein, 2003. The effect of lead and cadmium on LDH and G-6-PDH isozyme patterns exhibited during the early embryonic development of the teleost fish, Ctenopharyngodon idellus with emphasis on the corresponding morphological variations. Proceeding of the 26th Annual Larval Fish Conference, July 22-26, 2002, Bergen, Norway.
- Lemos, D., M. Salomon, V. Gomes, V.N. Phan and F. Buchholz, 2003. Citrate synthase and pyruvate kinase activities during early life stages of the shrimp Farfantepenaeus paulensis (Crustacea, Decapoda, Penaeidae): Effects of development and temperature. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol., 135: 707-719.
CrossRefDirect Link - Berges, J.A. and J.S. Ballantyne, 1991. Size scaling of whole-body maximal enzyme activities in aquatic crustaceans. Can. J. Fish. Aquat. Sci., 48: 2385-2394.
CrossRefDirect Link - Aellig, A., A. Albert, G. Blin, J. Buret and E. Daubrosse et al., 1982. Societe francaise de biologie clinique. Section of physiopathology. Commission reference values. Utilisation of reference values. (Document J, stage 3, version 1). Ann. Biol. Clin. (Paris), 40: 697-708.
PubMedDirect Link - Nishikimi, M., N.A. Rao and K. Yagi, 1972. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun., 46: 849-854.
CrossRefPubMedDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Gornall, A.G., C.J. Bardawill and M.M. David, 1949. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem., 177: 751-766.
CrossRefPubMedDirect Link - Utley, H.G., F. Bernheim and P. Hochstein, 1967. Effect of sulfhydryl reagents on peroxidation in microsomes. Arch. Biochem. Biophys., 118: 29-32.
CrossRefDirect Link - Buege, J.A. and S.D. Aust, 1978. Microsomal Lipid, Peroxidation. In: Methods in Enzymology, Vol. 52, Flesicher, S. and L. Packer (Eds.)., Academic Press, New York, pp: 302-310.
CrossRefDirect Link - Kurita-Ochiai, T., K. Fukushima and K. Ochiai, 1999. Lipopolysaccharide stimulates butyric acid-induced apoptosis in human peripheral blood mononuclear cells. Infect. Immunity, 67: 22-29.
Direct Link - Cobb, W.E., R.P. Lamberton and I.M. Jackson, 1984. Use of a rapid, sensitive immunoradiometric assay for thyrotropin to distinguish normal from hyperthyroid subjects. Clin. Chem., 30: 1558-1560.
PubMedDirect Link - Skelley, D.S., L.P. Brown and P.K. Besch, 1973. Radioimmunoassay. Clin. Chem., 19: 146-186.
PubMedDirect Link - Baxter, L., R.A. Brain, A.J. Hosmer, M. Nema, K.M. Muller, K.R. Solomon and M.L. Hanson, 2015. Effects of atrazine on egg masses of the yellow-spotted salamander (Ambystoma maculatum) and its endosymbiotic alga (Oophila amblystomatis). Environ. Pollut., 206: 324-331.
CrossRefDirect Link - Ji, Q., J. Lee, Y.H. Lin, G. Jing and L.J. Tsai et al., 2016. Atrazine and malathion shorten the maturation process of Xenopus laevis oocytes and have an adverse effect on early embryo development. Toxicol. In vitro, 32: 63-69.
CrossRefDirect Link - Liu, Z., Y. Wang, Z. Zhu, E. Yang, X. Feng, Z. Fu and Y. Jin, 2016. Atrazine and its main metabolites alter the locomotor activity of larval zebrafish (Danio rerio). Chemosphere, 148: 163-170.
CrossRefDirect Link - MacLoughlin, C., I.S. Canosa, G.R. Silveyra, L.S.L. Greco and E.M. Rodriguez, 2016. Effects of atrazine on growth and sex differentiation, in juveniles of the freshwater crayfish Cherax quadricarinatus. Ecotoxicol. Environ. Saf., 131: 96-103.
CrossRefDirect Link - Liu, Z., Z. Fu and Y. Jin, 2017. Immunotoxic effects of atrazine and its main metabolites at environmental relevant concentrations on larval zebrafish (Danio rerio). Chemosphere, 166: 212-220.
CrossRefDirect Link - Farah, M.A., B. Ateeq, M.N. Ali, R. Sabir and W. Ahmad, 2004. Studies on lethal concentrations and toxicity stress of some xenobiotics on aquatic organisms. Chemosphere, 55: 257-265.
CrossRefDirect Link - Pandey, S., R. Kumar, S. Sharma, N.S. Naghpure, S.K. Srivastava and M.S. Verma, 2005. Acute toxicity bioassays of mercuric chloride and malathion on air-breathing fish Channa punctatus (Bloch). Toxicol. Environ. Saf., 61: 114-120.
CrossRefDirect Link - Ayoola, S.O., 2008. Toxicity of glyphosate herbicide on Nile tilapia (Oreochromis niloticus) juvenile. Afr. J. Agric. Res., 3: 825-834.
Direct Link - Costa, M.J., D.A. Monteiro, A.L. Oliveira-Neto, F.T. Rantin and A.L. Kalinin, 2008. Oxidative stress biomarkers and heart function in bullfrog tadpoles exposed to roundup original®. Ecotoxicology, 17: 153-163.
CrossRefDirect Link - Do Carmo Langiano, V. and C.B.R. Martinez, 2008. Toxicity and effects of a glyphosate-based herbicide on the neotropical fish Prochilodus lineatus. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 147: 222-231.
CrossRefDirect Link - Lushchak, O.V., O.I. Kubrak, M.J. Storey, B.K. Storey and I.V. Lushchak, 2009. Low toxic herbicide roundup induces mild oxidative stress in goldfish tissues. Chemosphere, 76: 932-937.
CrossRefDirect Link - Kumar, A., M.B. Tyagi and P.N. Jha, 2004. Evidences showing ultraviolet-B radiation-induced damage of DNA in cyanobacteria and its detection by PCR assay. Biochem. Biophys. Res. Commun., 318: 1025-1030.
CrossRefDirect Link - Oulmi, Y., R.D. Negele and T. Braunbeck, 1995. Segment specificity of the cytological response in rainbow trout (Oncorhynchus mykiss) renal tubules following prolonged exposure to sublethal concentrations of atrazine. Ecotoxicol. Environ. Saf., 32: 39-50.
CrossRefPubMedDirect Link - Ramel, F., C. Sulmon, M. Bogard, I. Couee and G. Gouesbet, 2009. Differential patterns of reactive oxygen species and antioxidative mechanisms during atrazine injury and sucrose-induced tolerance in Arabidopsis thaliana plantlets. BMC Plant Biol., Vol. 9.
CrossRefDirect Link - Wiegand, C., E. Krause, C. Steinberg and S. Pflugmacher, 2001. Toxicokinetics of atrazine in embryos of the zebrafish (Danio rerio). Ecotoxicol. Environ. Saf., 49: 199-205.
CrossRefPubMedDirect Link - Kumaran, S.S., C. Kavitha, M. Ramesh and T. Grummt, 2011. Toxicity studies of nonylphenol and octylphenol: Hormonal, hematological and biochemical effects in Clarias gariepinus. J. Applied Toxicol., 31: 752-761.
CrossRefDirect Link - Mekkawy, I.A.A., U.M. Mahmoud, E.T. Wassif and M. Naguib, 2011. Effects of cadmium on some haematological and biochemical characteristics of Oreochromis niloticus (Linnaeus, 1758) dietary supplemented with tomato paste and vitamin E. Fish Physiol. Biochem., 37: 71-84.
CrossRefPubMedDirect Link - Thapar, A., R. Sandhir and R. Kiran, 2002. Acephate induced oxidative damage in erythrocytes. Indian J. Exp. Biol., 40: 963-966.
PubMed - Singh, M., R. Sandhir and R. Kiran, 2006. Erythrocyte antioxidant enzymes in toxicological evaluation of commonly used organophosphate pesticides. Indian J. Exp. Biol., 44: 580-583.
PubMed - Yildirim, N.C., F. Benzer and D. Danabas, 2011. Evaluation of environmental pollution at Munzur river of tunceli applying oxidative stress biomarkers in Capoeta trutta (Heckel, 1843). J. Anim. Plant Sci., 21: 66-71.
Direct Link - Trenzado, C., M.C. Hidalgo, M. Garcia-Gallego, A.E. Morales and M. Furne et al., 2006. Antioxidant enzymes and lipid peroxidation in sturgeon Acipenser naccarii and trout Oncorhynchus mykiss. A comparative study. Aquaculture, 254: 758-767.
CrossRefDirect Link - Velkova-Jordanoska, L., G. Kostoski and B. Jordanoska, 2008. Antioxidative enzymes in fish as biochemical indicators of aquatic pollution. Bulgarian J. Agric. Sci., 14: 235-237.
Direct Link - Borkovic-Mitic, S.S., T.B. Kovacevic, B.R. Perendija, S.G. Despotovic, J.P. Gavric, S.Z. Pavlovic and Z.S. Saicic, 2011. Superoxide dismutase and catalase activities in the digestive gland and gills of the freshwater bivalve Unio pictorum from the Sava River. Arch. Biol. Sci., 63: 185-192.
CrossRefDirect Link - An, A., Y. Li, X. Niu and H. Yu, 2008. Responses of antioxidant enzymes in catfish exposed to liquid crystals from E-waste. Int. J. Environ. Res. Public Health, 5: 99-105.
Direct Link - Vinodhini, R. and M. Narayanan, 2009. Biochemical changes of antioxidant enzymes in common carp (Cyprinus carpio L.) after heavy metal exposure. Turk. J. Vet. Anim. Sci., 33: 273-278.
CrossRefDirect Link - Modesto, K.A. and C.B.R. Martinez, 2010. Effects of roundup transorb on fish: Hematology, antioxidant defenses and acetylcholinesterase activity. Chemosphere, 81: 781-787.
CrossRefDirect Link - Giguere, A., P.G.C. Campbell, L. Hare and C. Cossu-Leguille, 2005. Metal bioaccumulation and oxidative stress in yellow perch (Perca flavescens) collected from eight lakes along a metal contamination gradient (Cd, Cu, Zn, Ni). Can. J. Fish. Aquat. Sci., 62: 563-577.
CrossRefDirect Link - Gad, N.S., 2011. Oxidative stress and antioxidant enzymes in Oreochromis niloticus as biomarkers of exposure to crude oil pollution. Int. J. Environ. Sci. Eng., 1: 49-58.
Direct Link - John, S., M. Kale, N. Rathore and D. Bhatnagar, 2001. Protective effect of vitamin E in dimethoate and malathion induced oxidative stress in rat erythrocytes. J. Nutr. Biochem., 12: 500-504.
CrossRefPubMedDirect Link - Datta, C., J. Gupta, A. Sarkar and D. Sengupta, 1992. Effects of organophosphorus insecticide phosphomidon on antioxidant defence components of human erythrocyte and plasma. Indian J. Exp. Biol., 30: 65-70.
PubMedDirect Link - St. Dimitrova, M., V. Tishinova and V. Velcheva, 1994. Combined effect of zinc and lead on the hepatic superoxide dismutase-catalase system in carp (Cyprinus carpio). Comp. Biochem. Physiol. Part C: Pharmacol. Toxicol. Endocrinol., 108: 43-46.
CrossRefDirect Link - Jayachandran, K., K. Pugazhendy, M. Susiladevi and C. Jayanthi, 2010. Protective role of atropine against atrazine toxicity on antioxidant enzymes in the fingerlings of Labeo rohita (Hamilton). Int. J. Rec. Scient. Res., 5: 15-20.
Direct Link - Haluzova, I., H. Modra, J. Blahova, M. Havelkova, Z. Siroka and Z. Svobodova, 2011. Biochemical markers of contamination in fish toxicity tests. Interdisciplin. Toxicol., 4: 85-89.
CrossRefDirect Link - Ferreira, D., T.C. Unfer, H.C. Rocha, L.C. Kreutz, G. Koakoski and L.J.G. Barcellos, 2012. Antioxidant activity of bee products added to water in tebuconazole-exposed fish. Neotrop. Ichthyol., 10: 215-220.
CrossRefDirect Link - Parthasarathy, R. and J. Joseph, 2011. Studies on the hepatic antioxidant defense system in λ cyhalothrin-induced oxidative stress in fresh water tilapia (Oreochromis mossambicus). Afr. J. Environ. Sci. Technol., 5: 530-534.
CrossRefDirect Link - Ceyhun, S.B., M. Senterk, D. Ekinci, O. Erdogan, A. Ciltas and E.M. Kocaman, 2010. Deltamethrin attenuates antioxidant defense system and induces the expression of heat shock protein 70 in rainbow trout. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 152: 215-223.
CrossRefPubMedDirect Link - Dorval, J. and A. Hontela, 2003. Role of glutathione redox cycle and catalase in defense against oxidative stress induced by endosulfan in adrenocortical cells of rainbow trout (Oncorhynchus mykiss). Toxicol. Applied Pharmacol., 192: 191-200.
CrossRefPubMedDirect Link - Prasanthi, K., Muralidhara and P.S. Rajini, 2005. Fenvalerate-induced oxidative damage in rat tissues and its attenuation by dietary sesame oil. Food Chem. Toxicol., 43: 299-306.
CrossRefDirect Link - Rodrigues, E.D.L. and E. Fanta, 1998. Liver histopathology of the fish Brachydanio rerio Hamilton-Buchman after acute exposure to sublethal levels of the organophosphate Dimethoate 500. Revista Brasileira Zoologia, 15: 441-450.
CrossRefDirect Link - Machado, M.R. and E. Fanta, 2003. Effects of the organophosphorous methyl parathion on the branchial epithelium of a freshwater fish Metynnis roosevelti. Braz. Arch. Biol. Technol., 46: 361-372.
CrossRefDirect Link - Yamano, T. and S. Morita, 1995. Effects of pesticides on isolated rat hepatocytes, mitochondria and microsomes II. Arch. Environ. Contam. Toxicol., 28: 1-7.
CrossRefDirect Link - Yildirim, N.C., M. Yurekli and N. Yildirim, 2010. Investigation of some antioxidant enzymes activities depending on adrenomedullin treatment and cold stress in rat liver tissue. Turk. J. Biochem., 35: 140-144.
Direct Link - Zhang, J., H. Shen, X. Wang, J. Wu and Y. Xue, 2004. Effects of chronic exposure of 2,4-dichlorophenol on the antioxidant system in liver of freshwater fish Carassius auratus. Chemosphere, 55: 167-174.
CrossRefDirect Link - El-Demerdash, F.M., M.I. Yousef, F.S. Kedwanyand and H.H. Baghdadi, 2004. Role of α-tocopherol and β-carotene in ameliorating the fenvalerate-induced changes in oxidative stress, hemato-biochemical parameters, and semen quality of male rats. J. Environ. Sci. Health B, 93: 443-459.
PubMedDirect Link - Sharma, Y., S. Bashir, M. Irshad, T.C. Nagc and T.D. Dogra, 2005. Dimethoate-induced effects on antioxidant status of liver and brain of rats following subchronic exposure. Toxicology, 215: 173-181.
CrossRefPubMedDirect Link - Kamboj, A., R. Kiran and R. Sandhir, 2006. Carbofuran-induced neurochemical and neurobehavioral alterations in rats: Attenuation by N-acetylcysteine. Exp. Brain Res., 170: 567-575.
CrossRefDirect Link - Atif, F., S. Parvez, S. Pandey, M. Ali and M. Kaur et al., 2005. Modulatory effect of cadmium exposure on deltamethrin-induced oxidative stress in Channa punctata Bloch. Arch. Environ. Contam. Toxicol., 49: 371-377.
CrossRefDirect Link - Koprucu, S.S., E. Yonar and E. Seker, 2008. Effects of deltamethrin on antioxidant status and oxidative stress biomarkers in freshwater mussel, Unio elongatulus eucirrus. Bull. Environ. Contam. Toxicol., 81: 253-257.
CrossRefDirect Link - Peebua, P., P. Kosiyachinda, P. Pokethitiyook and M. Kruatrachue, 2007. Evaluation of alachlor herbicide impacts on Nile Tilapia (Oreochromis niloticus) using biochemical biomarkers. Bull. Environ. Contam. Toxicol., 78: 138-141.
CrossRefDirect Link - Chandra, S., 2008. Toxic effect of malathion on acetylcholinesterase activity of liver, brain and gills of freshwater catfish Heteropneustes fossilis. Environ. Conserv., 9: 45-52.
Direct Link - Singh, M., R. Sandhir and R. Kiran, 2011. Effects on antioxidant status of liver following atrazine exposure and its attenuation by vitamin E. Exp. Toxicol. Pathol., 63: 269-276.
CrossRefDirect Link - Cooke, M.S., M.D. Evans, M. Dizdaroglu and J. Lunec, 2003. Oxidative DNA damage: Mechanisms, mutation and disease. FASEB J., 17: 1195-1214.
CrossRefPubMedDirect Link - Schmutzler, C., I. Gotthardt, P. Hofmann, B. Radovic and G. Kovacs et al., 2007. Endocrine disruptors and the thyroid gland-a combined in vitro and in vivo analysis of potential new biomarkers. Environ. Health Perspect., 115: 77-83.
CrossRefDirect Link - Leatherland, J.F., 1994. Reflections on the thyroidology of fishes: From molecules to humankind. Guelph Ichthyol. Rev., 2: 1-67.
Direct Link - Peter, M.C.S., R.A.C. Lock and S.E.W. Bonga, 2000. Evidence for an osmoregulatory role of thyroid hormones in the freshwater mozambique Tilapia Oreochromis mossambicus. Gen. Comp. Endocrinol., 120: 157-167.
CrossRefDirect Link - Oommen, O.V., P. Sreejith, R.S. Beyo, L. Divya, A.S. Vijayasree and M. Manju, 2006. Thyroid hormones regulate mitochondrial respiration as well as antioxidant defense in teleosts too! J. Endocrinol. Reprod., 10: 96-105.
Direct Link - Leji, J., G.S. Babitha, V. Rejitha, A.S. Vijayasree and J. Ignatius et al., 2007. Thyroidal and osmoregulatory responses in tilapia (Oreochromis mossambicus) to the effluents of coconut husk retting. J. Endocrinol., 11: 23-30.
Direct Link - Peter, V.S. and M.C.S. Peter, 2007. Influence of coconut husk retting effluent on metabolic, interrenal and thyroid functions in the air-breathing perch, Anabas testudineus Bloch. J. Endocrinol. Reprod., 11: 62-68.
Direct Link - Blanton, M.L. and J.L. Specker, 2007. The Hypothalamic-Pituitary-Thyroid (HPT) axis in fish and its role in fish development and reproduction. Crit. Rev. Toxicol., 37: 97-115.
CrossRefDirect Link - Eales, J.G., 1985. The peripheral metabolism of thyroid hormones and regulation of thyroidal status in poikilotherms. Can. J. Zool., 63: 1217-1231.
CrossRefDirect Link - Grau, E.G., 1988. Environmental influences on thyroid function in teleost fish. Am. Zool., 28: 329-335.
CrossRefDirect Link - Power, D.M., L. Llewellyn, M. Faustino, M.A. Nowell and B.T. Bjornsson et al., 2001. Thyroid hormones in growth and development of fish. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 130: 446-459.
CrossRefPubMedDirect Link - Mylonas, C.C., C.V. Sullivan and J.M. Hinshaw, 1994. Thyroid hormones in brown trout (Salmo trutta) reproduction and early development. Fish Physiol. Biochem., 13: 485-493.
CrossRefDirect Link - Reddy, P.K., C.L. Brown, J.F. Leatherland and T.J. Lam, 1992. Role of thyroid hormones in tilapia larvae (Oreochromis mossambicus): II. Changes in the hormones and 5'-monodeiodinase activity during development. Fish Physiol. Biochem., 9: 487-496.
CrossRefDirect Link - Weber, G.M., D.K. Okimoto, N.H. Richman III and E.G. Grau, 1992. Patterns of thyroxine and triiodothyronine in serum and follicle-bound oocytes of the tilapia, Oreochromis mossambicus, during oogenesis. Gen. Comp. Endocrinol., 85: 392-404.
CrossRefPubMedDirect Link - Ayson, F.G. and T.J. Lam, 1993. Thyroxine injection of female rabbitfish (Siganus guttatus) broodstock: Changes in thyroid hormone levels in plasma, eggs and yolk-sac larvae and its effect on larval growth and survival. Aquaculture, 109: 83-93.
CrossRefDirect Link - Tagawa, M., T. Ogasawara, T. Sakamoto, T. Miura, K. Yamauchi and T. Hirano, 1994. Thyroid hormone concentrations in the gonads of wild chum salmon during maturation. Fish Physiol. Biochem., 13: 233-240.
CrossRefDirect Link - Brown, C.L., S.I. Doroshov, M.D. Cochran and H.A. Bern, 1989. Enhanced survival in striped bass fingerlings after maternal triiodothyronine treatment. Fish Physiol. Biochem., 7: 295-299.
CrossRefDirect Link - Leatherland, J.F., L. Lin, N.E. Down and E.M. Donaldson, 1989. Thyroid hormone content of eggs and early developmental stages of five Oncorhynchus species. Can. J. Fish. Aquat. Sci., 46: 2140-2145.
CrossRefDirect Link - Tagawa, M. and T. Hirano, 1987. Presence of thyroxine in eggs and changes in its content during early development of chum salmon, Oncorhynchus keta. Gen. Comp. Endocrinol., 68: 129-135.
CrossRefDirect Link - Sinha, N., B. Lal and T.P. Singh, 1991. Pesticides induced changes in circulating thyroid hormones in the freshwater catfish Clarias batrachus. Comp. Biochem. Physiol. Part A: Comp. Physiol., 100: 107-110.
CrossRefDirect Link - Yadav, A.K. and T.P. Singh, 1986. Effect of pesticide on circulating thyroid hormone levels in the freshwater catfish, Heteropneustes fossilis (Bloch). Environ. Res., 39: 136-142.
CrossRefDirect Link - Brown, S.B., D.L. MacLatchy, T.J. Hara and J.G. Eales, 1990. Effects of low ambient pH and aluminum on plasma kinetics of cortisol, T3 and T4 in rainbow trout (Oncorhynchus mykiss). Can. J. Zool., 68: 1537-1543.
CrossRefDirect Link - Suchiang, P. and B.B.P. Gupta, 2011. Effects of partial and full feed restriction on the plasma levels of thyroid hormones and testicular activity in the male air-breathing catfish, Clarias gariepinus during different phases of the breeding cycle. Int. J. Biol., 3: 32-42.
CrossRefDirect Link - Oppenheimer, J.H., H.L. Schwartz, C.N. Mariash, W.B. Kinlaw, N.C.W. Wong and H.C. Freake, 1987. Advances in our understanding of thyroid hormone action at the cellular level. Endocr. Rev., 8: 288-308.
CrossRefPubMedDirect Link - Eales, J.G., 1988. The influence of nutritional state on thyroid function in various vertebrates. Am. Zool., 28: 351-362.
Direct Link - Van der Geyten, S., J.P. Sanders, V.M. Darras, E.R. Kuhn, J.L. Leonard and T.J. Visser, 1998. Cloning of tilapia type I and III deiodinases. Ann. N. Y. Acad. Sci., 839: 498-499.
CrossRefDirect Link - Reyns, G.E., K.A. Janssens, J. Buyse, E.R. Kuhn and V.M. Darras, 2002. Changes in thyroid hormone levels in chicken liver during fasting and refeeding. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol., 132: 239-245.
CrossRefDirect Link