
Mohamed Mohamed Soliman
Medical Laboratories, Turabah and College of Science, Taif University, Saudi Arabia
Hossam Fouad Attia
Medical Laboratories, Turabah and College of Science, Taif University, Saudi Arabia
Samir Ahmed El-Shazly
Department of Biochemistry and Histology, Benha University, Moshtohor, P.O. 13736 and Kafr El-Sheikh University, Egypt
Osama Mesilhy Saleh
Department of Biochemistry and Histology, Benha University, Moshtohor, P.O. 13736 and Kafr El-Sheikh University, Egypt
American Journal of Biochemistry and Molecular Biology
Year: 2012 | Volume: 2 | Issue: 3 | Page No.: 133-145







ABSTRACT
Cinnamon Extracts (CE) are widely used in Middle East and Asian countries as herbal medication for diabetes. It contains biologically active substances which have in vivo hypoglycemic activity and probably improve insulin sensitivity. Up till now the molecular mechanism of CE on genes expression is not yet determined. In this study, we examined the effect of CE (200 mg kg-1 body weight) in obesity and diabetic Wistar rats by testing its effect on genes related to lipids and glucose metabolism. Firstly, to test anti-obesity actions of CE, rats were fed High Fat Diet (HFD) for 3 months then treated with CE for 2 months. Secondly, experimentally diabetic rats were treated with CE for 2 months to examine its antidiabetic action. The results showed that CE treatment did not show any significant changes in plasma levels of obesity related parameters including lipid profiles, leptin and lipase enzyme. However, liver histology showed decrease in lipid and glycogen accumulation in CE treated rats. Moreover, CE induced significant (p<0.05) decrease in haptoglobin levels. In diabetic experiments, CE treatment normalized significantly (p<0.05) the increase in lipid profiles and glucose levels occurred in diabetic rats. In parallel, CE lowered glucose levels through the increase in insulin secretion from still working β cells. The antidiabetic action of CE was confirmed as CE treated diabetic rats showed an increase in mRNA expression of leptin, PPAR-gamma and adiponectin. In conclusion, CE is not anti-obesity herb but has antidiabetic and insulin-mimetic actions through the regulation of genes related to lipid and glucose metabolism.
PDF Abstract XML References Citation
Received: October 24, 2011;
Accepted: December 15, 2011;
Published: February 21, 2012
How to cite this article
Mohamed Mohamed Soliman, Hossam Fouad Attia, Samir Ahmed El-Shazly and Osama Mesilhy Saleh, 2012. Biomedical Effects of Cinnamon Extract on Obesity and Diabetes Relevance in Wistar Rats. American Journal of Biochemistry and Molecular Biology, 2: 133-145.
DOI: 10.3923/ajbmb.2012.133.145
URL: https://scialert.net/abstract/?doi=ajbmb.2012.133.145
DOI: 10.3923/ajbmb.2012.133.145
URL: https://scialert.net/abstract/?doi=ajbmb.2012.133.145
INTRODUCTION
Obesity and diabetes are chronic metabolic diseases associated with an increased risk of coronary heart disease, stroke, hypertension, renal failure, type 2 diabetes, dyslipidemia and all cause mortality (Hall, 2003; Havel, 2004; Trayhurn and Beattie, 2001). Clinically diabetic patients characterized by marked increase in blood glucose levels followed by mild hyperlipidemia (Reddy et al., 2009). Traditional herbal medicine has been widely used for diabetes treatment and is recognized as an interesting alternative to conventional medicine (Rao et al., 1997) especially in the third world countries and therefore, represent new avenues in the search for alternative hypoglycemic drugs (Day, 1998). However, most of them have been shown to exert little or no effect on glycemic control in experimental studies, although some herbs possess hypoglycemic properties. Treatment of diabetes depends on genes related to glucose metabolism. For example, glucose transport is the rate-limiting step in carbohydrate metabolism (Maughan, 2009) which is facilitated by glucose transporters (GLUT) across the cell membrane (Anand et al., 2010). So, compounds facilitating GLUT4 translocation and improve insulin sensitivity can be beneficial for the treatment of diabetes (Kipmen-Korgun et al., 2009; Shepherd and Kahn, 1999). Usage of natural products as cinnamon and other dietary modulators with anti-diabetic activity are the first choice of diabetic patients. This tendency is because insulin, to date, cannot be used orally and its repeated injections had many undesirable adverse effects. In addition, most of hypoglycemic agents or drugs are not effective to decrease blood glucose levels in chronic diabetic patients (Cheng and Fantus, 2005).
Cinnamon is the bark of the Cinnamomi cassiae (Lauraceae), it contains cinnamic aldehyde, cinnamic acid, tannin and Methyl-Hydroxychalcone Polymer (MHCP) as main components. Cinnamon Extract (CE) contains biologically active substances with insulin-mimetic properties (Kim et al., 2006). Qin et al. (2003) have reported that cinnamon extract decreases blood glucose and lipid profiles in rats and increases insulin sensitivity and glucose uptake in adipocytes (Khan et al., 2003a; Jarvill-Taylor et al., 2001; Safdar et al., 2004; Yaghmoor and Khoja, 2010). In vitro and in vivo studies have shown that cinnamon enhances glucose uptake by activating insulin receptor kinase activity, autophosphorylation of insulin receptor and glycogen synthase activity (Kannappan et al., 2006). Moreover, it posses the ability to reduce lipid levels in fructose-fed rats and affects immune responses by regulating anti-, proinflammatory and glucose transporter gene expressions in mouse macrophages (Cao et al., 2008). It has been shown that, cinnamon extract has antioxidant activity (Muchuweti et al., 2007), decreases oxidative stress activity (Ranjbar et al., 2007), with antimicrobial and anti-parasitic effects (Hussien et al., 2011; Vinitha and Ballal, 2008) and can be used for treatment of osteoarthiritis as anti-analgesic factor (Zahmatkash and Vafaeenasab, 2011). Also, cinnamon has hepatoprotective effect against carbon tetrachloride induced oxidative stress and liver injury in rats (Al-Attar, 2007; Moselhy and Ali, 2009). Clinical and animal studies have confirmed a strong relationship between obesity and hypertension with diabetes (Mohamed-Ali et al., 1998). Therefore, we can speculate that there is an interaction between cinnamon, diabetes and obesity through its effect on body weight, insulin, leptin and genes related to lipid and glucose metabolism and may be useful in their treatment. The present results reported that CE is not anti-obesity herbal medication but posses anti-diabetic properties and improve insulin sensitivity through regulation of various gene expression.
MATERIALS AND METHODS
Materials: Streptozotocin (STZ) was from sigma Aldrich, USA. The Wistar albino rats were from Egyptian Co for experimental animals import, Helwan, Egypt. Vehicles and related materials were from ADWIA pharmaceutical company, Egypt, Heparinzed vacuteiner tubes, TriZol reagents, Poly dT, chloroform, ethanol and cytokines primers were from Wako pure chemicals, Osaka, Japan. Biochemical kits for lipids, insulin, leptin and haptoglobin were from Clini Lab, Cairo, Egypt.
Cinnamon extracts preparation: Cinnamon extract was extracted based on method of Sheng et al. (2008). Briefly, cinnamon powder (100 g) was dissolved in 1000 mL double distilled water then subjected for revolving evaporator in vacuum state using vacuum pump till the volume of water reduced to 50%. The supernatant was filtered through Whatman paper No. 1 to obtain cinnamon water extract. The final concentration was measured by Lowry method for protein concentration.
Study design
Induction of obesity in wistar rats: Twenty rats 3 week old, weighing 75-85 g were selected randomly and given High Fat Diet (HFD). The HFD rats (HFD, n = 20) were fed a high-fat diet (15.5% protein, 38.8% fat and 45.7% carbohydrates, by calories) for 3 months. Obesity was confirmed by the increase in lipid parameters and body weight (data not shown). Next, rats were subdivided into 2 subgroups. Control (HFD group, n = 20) was given water as a vehicle and the second group was given CE and HFD (CE+HFD, n = 20). Cinnamon Extract (CE) was given in a dose of 200 mg kg-1 body weight orally and daily for 2 months as stated by Kim et al. (2006), Kim and Choung (2010).
Induction of diabetes in wistar rats: Adult Wistar rats weighting 200-250 g (90 days old) were used for induction of diabetes. The animals were injected STZ at the dose of 60 mg kg-1 of the body weight intraperitoneally. STZ induced diabetes within 3 days of injection as confirmed by the increase in blood glucose samples collected 3 days after STZ injection. The blood glucose levels over 200 mg dL-1 considered diabetic rats. Diabetic and non-diabetic control groups were kept in metabolic cages individually and separately. Next, rats were divided as follows:
| • | Control receiving water as vehicle |
| • | Control diabetic received STZ single dose then water as vehicle |
| • | Diabetic plus Cinnamon Extract (CE) in a dose of 200 mg kg-1 body weight |
Rats in all experiments were killed by decapicitation after 2 months of CE treatments. Blood was collected to get plasma and tissues were kept in TriZol reagent at -70°C until RNA extraction and mRNA expression.
Plasma chemistry analysis: Serum Triglycerides (TG), Total Cholesterol (TC), VLDL and HDL were measured using commercial kits that based on spectrophotometric analysis. Leptin, haptoglobin and lipase were measured using ELISA kits. Insulin and glucose were measured by commercial kits supported by Wako pure chemicals, Japan.
Liver histology: The liver tissues were fixed in 10% formalin and embeded in paraffin wax. Frozen liver sections from fixed tissues were cut in 10 μm thick and mounted on the slide glasses. To detect lipids, the sections were immersed in propylene glycol for 5 min and stained them with Oil red O (Sigma) for 7 min. Samples then were rinsed with 85% propylene glycol and water, counterstained with Mayer’s haematoxylin and copied by microscope. For glycogen demonstration, liver samples were stained with Best's carmine stain and PAS technique according to Bancroft et al. (1994).
RT-PCR analysis and gene expression: Livers and adipose tissue were collected from rats, flash frozen in liquid nitrogen and subsequently stored at -70°C. Frozen samples (approximately 100 mg of tissue per sample) were immediately added to 1 mL of TriZol reagent (Invitrogen, Carlsbad, CA) and homogenized using a Polytron 300 D homogenizer (Brinkman Instruments, Westbury, NY).
| Table 1: | PCR conditions for examined genes |
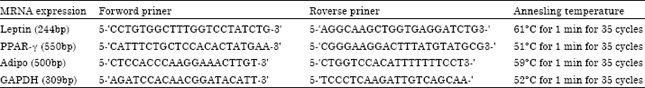 | |
PCR cycle of respective genes are shown, while temperature and time of denaturation and elongation steps of each PCR cycle are 94°C, 30sec and 72°C, 60sec, respectively | |
One milliliter of the tissue homogenate was transferred to a microfuge tube and total RNA was extracted via chloroform extraction followed by nucleic acid precipitation with isopropanol. The pellet was washed with 75% ethanol and resuspended in molecular biology grade water. Nucleic acid concentration was determined by o.d. 260 nm (Smart-Spec; Bio-Rad Laboratories, Hercules, CA), and RNA integrity was evaluated using an Agilent bioanalyzer (model 2100; Agilent Technologies, Foster City, CA).
RNA (1 μg) was treated at 72°C for 5 min and reverse transcribed using 100 units of Moloney murine leukemia virus reverse transcriptase (Gibco), 50 pmol of poly (dT) primer and 20 nmol of dNTPs in a total volume of 10 μL at 37°C for 1 h. After heating at 94°C for 5 min, PCR amplification was performed with 2.5 units Taq polymerase (Perkin-Elmer, Foster City, CA, USA), 3 mM MgCl2 and 50 pmol of forward and reverse primers specific for respective genes in a total volume of 50 μL. The PCR conditions for different tested genes as adiponectin, PPAR-γ, leptin and GLUT4 carried out as in Table 1. Electrophoresis in 1.5% agarose gel stained with ethidium bromide and visualization under UV lamp will be carried out. Intensities of PCR bands will be analyzed densitometrically using NIH Image program (http://rsb.info.nih.gov/nih-image/).
Statistical analysis: Results are expressed as means±S.E. of independent experiments. Statistical analysis was done using ANOVA and Fischer’s post hoc test, with p<0.05 being considered as statistically significant.
RESULTS
Effect of CE on changes in body weight, food intake and lipid profiles in diet induced obesity in wistar rats: Treatment of HFD fed rats with cinnamon extract induced minor changes in body weight (10% decreases) relative to obese (HFD alone) control rats without any significant changes in percent of food intake as seen in Fig. 1(a-b). After 60 days of CE treatment, plasma lipid parameters were assayed. As seen in Fig. 1c-f, CE did not induce any significant alteration in levels of cholesterol, TG, VLDL and HDL in both normal obese and CE treated obese rats.
Effect of CE on hepatic lipids and glycogen changes in high fat diet fed wistar rats: Although the changes in body weight are not clear, liver histology showed significant decrease in lipids accumulation in liver cells (Fig. 2a-b). Staining of liver by H and E stain shows large fat globules that replaced the contour of hepatocytes in obese rats but in CE treated rats, regeneration of hepatocytes in most of hepatic cores occurred and a decrease in size of fat globules and the number of lipid droplets in the liver tissues was markedly reduced in the cinnamon treated rats. It replaced by fine vacuoles in the contour of the hepatocytes. Moreover, glycogens storage was seen in normal and CE treated rats.
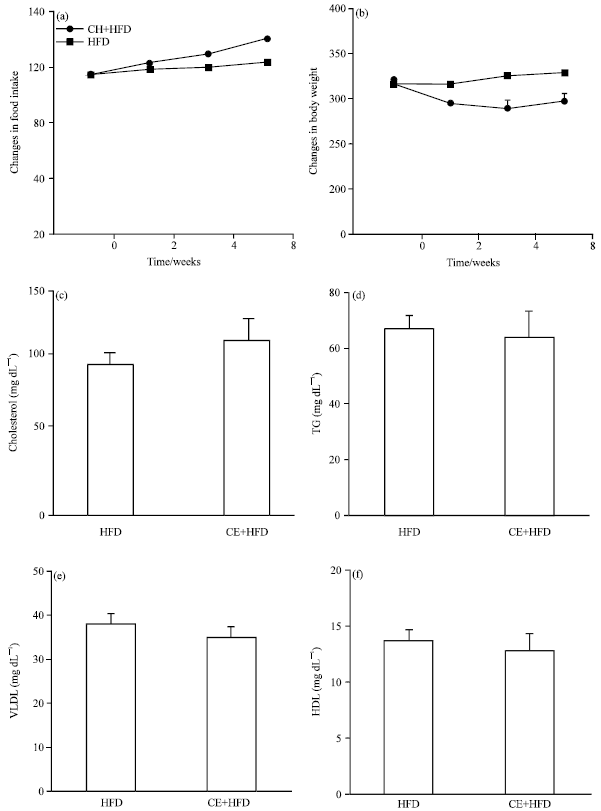 |
| Fig. 1: | Effect of cinnamon extract on changes in body weight (a), food intake (b) and lipid profiles (c-f) in obese Wistar rats. Rats were given high fat diet (HFD) for 3 months then treated either by cinnamon extract (CE+HFD) or vehicle in water for 2 months. During treatment, body weight and food intake were measured at indicated times in figures. Lipid parameters were measured after 2 months of treatment. Values are means±SEM of 3 independent experiments each one contains 5 rats |
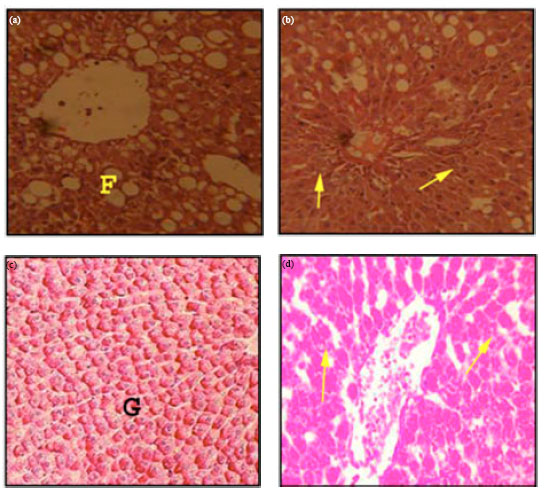 |
| Fig. 2(a-d): | Effect of cinnamon extract on liver histology. Lipid accumulation (a-b) and glycogen storage (c-d) were recorded in obese Wistar rats and cinnamon treated obese rats. Rats were treated either cinnamon extract (CE+HFD) or vehicle in water for 2 months. Liver histology was examined by H and E stain for lipids accumulations and Best's carmine and PAS stain for glycogen storage. F in (a) is fat globules, Arrows in (b) indicate decrease fat accumulation, G in (c) indicate numerous glycogen granules around the nuclei and Arrows in (d) indicate vacuoles are dissolved glycogen granules by PAS |
The liver of normal obese rats (Fig. 2c) stained with best’s carmine showed accumulation of glycogen granules in cytoplasm of hepatocytes around the basophilic nuclei. While liver of CE treated rats (Fig. 2d) stained by PAS showed dissolved glycogen granules in the cytoplasm of the hepatocytes. That results may suggests a possible role for cinnamon in obesity relevance. So, we examined the changes in leptin and other obesity related proteins.
Effect of CE on plasma levels of leptin, haptoglobin and lipase in high fat diet fed wistar rats: Next we examined the changes in plasma levels of some proteins related to obesity occurrence. CE tretment for 2 months did not induce any significant changes in plasma levels of leptin (Fig. 3a). Leptin as known increased with obesity to stimulate peripheral glucose utilization and lipolytic activity. In parallel, CE didn't change lipase acyivity but decrease haptoglobin secretion (Fig. 3b-c). These findings suggest that CE is not anti-obesity but acts as anti-inflammatory herbal medication.
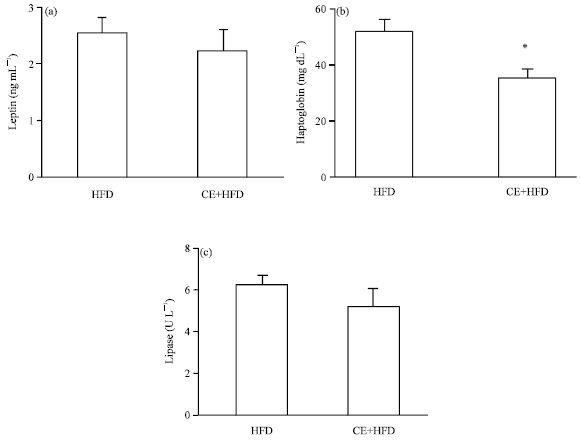 |
| Fig. 3(a-c): | Effect of cinnamon extract on leptin (a), haptoglobin (b) and lipase (c) in obese Wistar rats. HFD rats were treated either cinnamon extract (CE group) or vehicle in water and food for 2 months. Leptin, Haptoglobin and lipase were measured by ELISA kits. Values are means±SEM of 3 independent experiments each one contains 5 rats. *p<0.05 vs. control |
Effect of cinnamon extract on changes of lipids profiles, glucose and insulin in normal, diabetic control and CE-treated diabetic Wistar rats.
CE treatment induced alteration in plasma levels of lipid parameters. As seen in Fig. 4a-d, diabetic rats showed significant increase in cholesterol, TG, VLDL and decrease in HDL compared to normal non diabetic rats. CE treatment normalized the changes induced in diabetic rats. Moreover the CE decreased the significant increase in glucose concentration in diabetic rats (Fig. 4e). Plasma insulin levels was decrease in diabetic rats as results of pancreatic γ cells destruction and CE treatment stimulated the remaining cells to increase insulin secretion (Fig. 4f).
RT-PCR analysis and mRNA expression in epididymal fat tissues: Finally, we tested the effect of CE on modulation of leptin, PPAR-γ and adiponectin expression in rat epididymal adipose tissue using RT-PCR analysis. STZ diabetic rats have no alteration in leptin expression but treatment of diabetic rats by CE significantly increased leptin mRNA expression relative to control and diabetic control rats (Fig. 5a). Moreover, the expression of PPAR-γ, a type of nuclear regulatory protein involved in transcription of genes regulating glucose and fat metabolism was examined. As seen in Fig. 5b, STZ induced less significant increase in PPAR-γ expression (p<0.05) but treatment of STZ diabetic rats by CE induced 4 folds increase in PPAR-γ expression to increase peripheral glucose utilization and lipid metabolism.
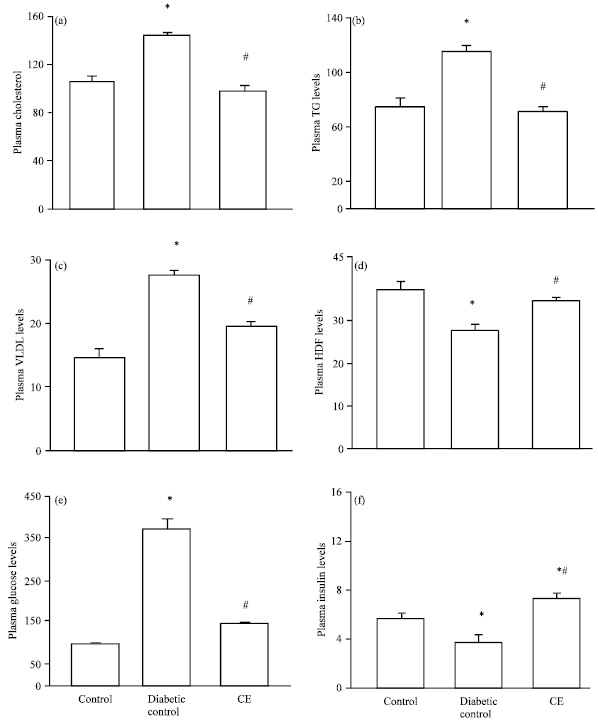 |
| Fig. 4(a-f): | Effect of cinnamon extract treatment on plasma changes of lipids profiles (a-d), glucose (e) and insulin (f) in normal, diabetic and CE-treated diabetic rats. Wistar rats were given cinnamon extract in water for 2 months. Plasma levels of cholesterol. TG, VLDL, HDL, glucose and insulin were measured for 3 different experiments. Values are means±SEM of 5 different rats for each experiment. *p<0.05 vs. control and #p<0.05 vs. diabetic rats |
At the end, we examined the expression of insulin sensitizing protein, adiponectin. Adiponectin expression was double fold increased (p<0.05) in CE treated rats compared to control and STZ diabetic rats as seen in Fig. 5c.
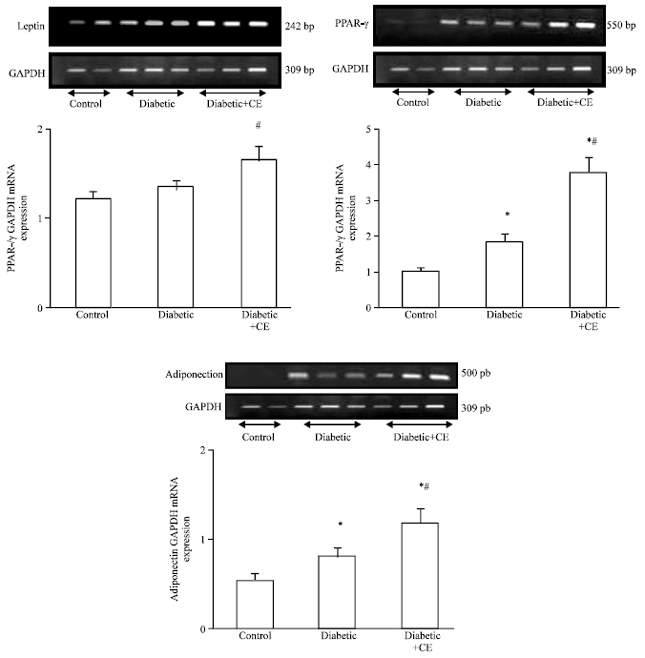 |
| Fig. 5: | RT-PCR analysis of leptin, PPAR-γ and adiponectin in epididymal fat tissue of rats. Rats were treated by either vehicle or CE for 2 months. RNA was extracted and reverse transcribed (1 μg) and RT-PCR analysis was carried out for leptin, PPAR-γ, adiponectin and GAPDH. The densitometric analysis of expressed bands (lower columns) was normalized with that of GAPDH and then relative to control. Values are means±SEM obtained from 3 different experiments. *p<0.05 vs. compared to control and. #p<0.05 vs. diabetic group alone |
DISCUSSION
In the present study, we have shown that treatment of STZ diabetic rats by water extract of cinnamon induced insulin mimetic effects and improved insulin sensitivity. However, CE extract did not induce any anti-obesity effects although a noticeable decrease in liver fat and glycogen deposition was recorded. The novel point is the decrease in haptoglobin secretion, a protein known to increase in obesity and is a marker of inflammation in animals and humans (Hall et al., 2003). Feeding of rats by high fat diet induced insulin resistance which is prevented by CE administration (Anand et al., 2010; Qin et al., 2003, 2004). Insulin resistance is strongly associated with metabolic dyslipidemia in diabetes and obesity. In obesity, our findings showed that CE treatments failed to modulate any changes in obesity related proteins and no changes in body weight, food intake and lipid profiles, same findings were reported by Kim et al. (2006). Although Qin et al. (2009), reported that the use of water-soluble CE improved the metabolic syndrome by regulating the circulating adipose-derived factors and multiple genes related to carbohydrate metabolism and lipogenesis. However, the only less significant finding in our study is that CE decreases hepatic fat deposition and glycogen deposition and suggest that more experiments are needed.
As known, diabetes is a chronic metabolic disorder that affects approximately 3% of population worldwide. Gaster and Hirsch (1998) reported that Sustained reductions in hyperglycemia will decrease the risk of developing microvascular diseases and reduce diabetes complications. Usage of oral hypoglycemic drugs to treat diabetes has several limitations, such as adverse effects and high rates of secondary failure (Kim et al., 2006). Those adverse effects forced the diabetic patients to use herbal medication that have a similar degree of efficiency without side effects and that was the purpose of this study.
CE treatment decreased glucose levels in STZ diabetic rats and that confirmed the insulin like effects of CE extract and that is seen by increase in insulin levels in plasma. The possible mechanism by which CE has its hypoglycemic action in diabetic Wistar rats may be by potentiating the effect of insulin in plasma or by increasing either the pancreatic secretion of insulin from the existing beta cells or its release from the bound form (Kim et al., 2006). CE might improve diabetes by normalizing the postprandial plasma glucose level as well as fasting blood glucose level (Kim et al., 2009; Khan and Safdar, 2003). In turn, in hyperinsulinemia, cinnamon increases insulin sensitivity for effective glucose disposal in rats although in humans cinnamon does not appear to improve fasting blood glucose levels and lipid parameters in patients with type 1 or type 2 diabetes (Baker et al., 2008; Khan et al., 2010). Here, CE improved and normalized the the changes in lipid parameters as TG, cholesterol and HDL and that is parallel with results of Khan et al. (2003b), Kim and Choung (2010) and Qin et al. (2003) and improved insulin resistance induced by feeding high fat diet (Thorens, 1996).
To understand the molecular mechanism of CE, we examined the mRNA expression of proteins involved in lipid and glucose metabolism in diabetic epididymal adipose tissue. Treatment of STZ diabetic rats with CE induced significant increase leptin, PPAR-γ and adiponectin expression. The correlation between glucose, insulin, leptin, PPAR-γ and adiponectin is the mechanism by which CE induced its antidiabetic effects. As the decrease in glucose levels in diabetic rats was correlated with the increase insulin secretion from working γ cells as suggested by our findings and that of Kim and Choung (2010). Also, the decrease in lipid parameters may be associated with the increase of leptin mRNA expression in diabetic rats, because leptin is known to be the potent lipolytic protein and increased peripheral glucose utilization (Ahima and Flier, 2000). Moreover, PPARγ mRNA expression was up-regulated in adipose tissue by the administration of the cinnamon extract. As known, PPARγ is highly expressed in adipose tissue and plays an important role in insulin sensitivity and the secretion of adipocytokines such as adiponectin. PPARγ activation through the binding of the synthetic thiazolidinediones, PPAR-γ agonists, results in a marked improvement in insulin and glucose in type 2 diabetic patients which results in improvement of the whole body insulin sensitivity (Frias et al., 2000; Miyazaki et al., 2001; Raskin et al., 2000). It has been shown that PPARγ increases the synthesis and production of adiponectin in animals and humans (Yamauchi et al., 2001; Fruebis et al., 2001; Maeda et al., 2001). The increase in expression of adiponectin constitutes the mechanism by which PPAR-γ acts in adipose tissue to increase whole-body insulin sensitivity (Yamauchi et al., 2001). So, the increase in insulin secretion and decrease in glucose levels and lipid parameters may be mediated by the increase in PPAR-γ (Yamauchi et al., 2001) and possibly by adiponectin expression.
In conclusion, this study demonstrated that CE is more anti-diabetic herbal medication through the improvement of insulin sensitivity, decrease in blood glucose levels and increases the expression of proteins related lipid metabolism. Moreover, cinnamon extract is not pure anti-obesity herbal therapy.
ACKNOWLEDGMENT
This study was supported in part by a Grant-in-Aid of the dean of Scientific Research (No.1051-1-1432), Faculty of Applied Medical Sciences, Taif University, Saudi Arabia and Faculty of Veterinary Medicine, Benha University, Egypt.
REFERENCES
- Al-Attar, A.M., 2007. Chemopreventive effect of cinnamon extract on carbon tetrachloride-induced physiological changes in the frog, Rana ridibunda. J. Applied Sci., 7: 79-90.
CrossRefDirect Link - Anand, P., K.Y. Murali, V. Tandon, P.S. Murthy and R. Chandra, 2010. Insulinotropic effect of cinnamaldehyde on transcriptional regulation of pyruvate kinase, phosphoenolpyruvate carboxykinase and GLUT4 translocation in experimental diabetic rats. Chemico-Biol. Interact., 186: 72-81.
CrossRefDirect Link - Baker, W.L., G. Gutierrez-Williams, C.M. White, J. Kluger and C.I. Coleman, 2008. Effect of cinnamon on glucose control and lipid parameters. Diabetes Care, 31: 41-43.
CrossRefPubMedDirect Link - Cao, H., J.F. Urban, Jr. and R.A. Anderson, 2008. Cinnamon polyphenol extract affects immune responses by regulating anti- and proinflammatory and glucose transporter gene expression in mouse macrophages. J. Nutr., 138: 833-840.
CrossRefDirect Link - Cheng, A.Y.Y. and I.G. Fantus, 2005. Oral antihyperglycemic therapy for type 2 diabetes mellitus. Can. Med. Assoc. J., 172: 213-226.
CrossRefPubMedDirect Link - Day, C., 1998. Traditional plant treatments for diabetes mellitus: Pharmaceutical foods. Br. J. Nutr., 80: 5-6.
CrossRefPubMedDirect Link - Frias, J.P., J.G. Yu, Y.T. Kruszynska and J.M. Olefsky, 2000. Metabolic effects of troglitazone therapy in type 2 diabetic, obese and lean normal subjects. Diabetes Care, 23: 64-69.
CrossRefPubMedDirect Link - Fruebis, J., T.S. Tsao, S. Javorschi, D. Ebbets-Reed and M.R. Erickson et al., 2001. Proteolytic cleavage product of 30-kDa adipocyte complement-related protein increases fatty acid oxidation in muscle and causes weight loss in mice. Proc. Natl. Acad. Sci. USA., 98: 2005-2010.
CrossRefPubMedDirect Link - Gaster, B. and I.B. Hirsch, 1998. The effects of improved glycemic control on complications in type 2 diabetes. Arch. Int. Med., 158: 134-140.
CrossRefDirect Link - Hall, J.E., 2003. The kidney, hypertension and obesity. Hypertension, 41: 625-633.
CrossRefPubMedDirect Link - Havel, J.P., 2004. Updates on adipocytes hormones: Regulation of energy balance and carbohydrate∕ lipid metabolism. Diabetes, 53: S143-S151.
PubMed - Hussien, J., C. Teshale and J. Mohammed, 2011. Assessment of the antimicrobial effects of some ethiopian aromatic spice and herb hydrosols. Int. J. Pharmacol., 7: 635-640.
CrossRef - Jarvill-Taylor, K.J., R.A. Anderson and D.J. Graves, 2001. A hydroxychalcone derived from cinnamon functions as a mimetic for insulin in 3T3-L1 adipocytes. J. Am. Coll. Nutr., 20: 327-336.
CrossRefDirect Link - Rao, B.K., R. Giri, M.M. Kesavuln and C.H. Apparao, 1997. Herbal medicine in the management of diabetes mellitus. Manphar Vaidhya Patrikal, 1: 33-35.
Direct Link - Kannappan, S., T. Jayaraman, P. Rajasekar, M.K. Ravichandran and C.V. Anuradha, 2006. Cinnamon bark extract improves glucose metabolism and lipid profile in the fructose-fed rat. Singapore Med. J., 47: 858-863.
PubMedDirect Link - Khan, A., M. Safdar, M.M.A. Khan, K.N. Khattak and R.A. Anderson, 2003. Cinnamon improves glucose and lipids of people with Type 2 diabetes. Diabetes Care, 26: 3215-3218.
CrossRefPubMedDirect Link - Khan, R., Z. Khan and S.H. Shah, 2010. Cinnamon may reduce glucose, lipid and cholesterol level in type 2 diabetic individuals. Pak. J. Nutr., 9: 430-433.
CrossRefDirect Link - Khan, A., M. Safdar and M.M.A. Khan, 2003. Effect of various doses of cinnamon on lipid profile in diabetic individuals. Pak. J. Nutr., 2: 312-319.
CrossRefDirect Link - Kim, S.H., S.H. Hynn and S.Y. Choung, 2006. Antidiabetic effect of cinnamon extract on blood glucose in db/db mice. J. Ehthnopharmacol., 104: 119-123.
CrossRefDirect Link - Kim, J.O., G.D. Lee, J.H. Kwon and K.S. Kim, 2009. Anti-diabetic effects of new herbal formula in neonatally streptozotocin-induced diabetic rats. Biol. Pharm. Bull., 32: 421-426.
CrossRefPubMedDirect Link - Kipmen-Korgun, D., S. Bilmen-Sarikcioglu, H. Altunbas, R. Demir and E.T. Korgun, 2009. Type-2 diabetes down-regulates glucose transporter proteins and genes of the human blood leukocytes. Scand. J. Clin. Lab. Invest., 69: 350-358.
CrossRefPubMedDirect Link - Maeda, N., M. Takahashi, T. Funahashi, S. Kihara and H. Nishizawa et al., 2001. PPARγ ligands increase expression and plasma concentrations of adiponectin, an adipose-derived protein. Diabetes, 50: 2094-2099.
CrossRefPubMedDirect Link - Miyazaki, Y., A. Mahankali, M. Matsuda, L. Glass and S. Mahankali et al., 2001. Improved glycemic control and enhanced insulin sensitivity in type 2 diabetic subjects treated with pioglitazone. Diabetes Care, 24: 710-719.
CrossRefPubMedDirect Link - Mohamed-Ali, V., J.H. Pinkney and S.W. Coppack, 1998. Adipose tissue as an endocrine and paracrine organ. Int. J. Obes. Relat. Metab. Disord., 22: 1145-1158.
CrossRefDirect Link - Moselhy, S.S. and H.K. Ali, 2009. Hepatoprotective effect of Cinnamon extracts against carbon tetrachloride induced oxidative stress and liver injury in rats. Biol. Res., 42: 93-98.
CrossRefPubMedDirect Link - Muchuweti, M., E. Kativu, C.H. Mupure, C. Chidewe, A.R. Ndhlala and M.A.N. Benhura, 2007. Phenolic composition and antioxidant properties of some spices. Am. J. Food Technol., 2: 414-420.
CrossRefDirect Link - Qin, B., M. Nagasaki, M. Ren, G. Bajotto, Y. Oshida and Y. Sato, 2003. Cinnamon extract (traditional herb) potentiates in vivo insulin-regulated glucose utilization via enhancing insulin signaling in rats. Diabetes Res. Clin. Pract., 62: 139-148.
CrossRefPubMedDirect Link - Qin, B., M.M. Polansky and R.A. Anderson, 2010. Cinnamon extract regulates plasma levels of adipose-derived factors and expression of multiple genes related to carbohydrate metabolism and lipogenesis in adipose tissue of fructose-fed rats. Horm. Metab. Res., 42: 187-193.
CrossRefPubMedDirect Link - Qin, B., M. Nagasaki, M. Ren, G. Bajotto, Y. Oshida and Y. Sato, 2004. Cinnamon extract prevents the insulin resistance induced by a high-fructose diet. Horm. Metab. Res., 36: 119-125.
CrossRef - Ranjbar, A., S. Ghaseminejhad, H. Takalu, A. Baiaty, F. Rahimi and M. Abdollahi, 2007. Anti oxidative stress potential of cinnamon (Cinnamomum zeylanicum) in operating room personnel; a before/after cross sectional clinical trial. Int. J. Pharmacol., 3: 482-486.
CrossRefDirect Link - Raskin, P., E.B. Rappaport, S.T. Cole, Y. Yan, R. Patwardhan and M.I. Freed, 2000. Rosiglitazone short-term monotherapy lowers fasting and post-prandial glucose in patients with type II diabetes. Diabetologia, 43: 278-284.
CrossRefPubMedDirect Link - Reddy, K.P., A.B. Singh, A. Puri, A.K. Srivastava and T. Narender, 2009. Synthesis of novel triterpenoid (lupeol) derivatives and their in vivo antihyperglycemic and antidyslipidemic activity. Bioorgan. Med. Chem. Lett., 19: 4463-4466.
CrossRefPubMedDirect Link - Khan, A. and M. Safdar, 2003. Role of diet, nutrients, spices and natural products in diabetes mellitus. Pak. J. Nutr., 2: 1-12.
CrossRef - Safdar, M., A. Khan, M.M.A.K. Khattak and M. Siddique, 2004. Effect of various doses of cinnamon on blood glucose in diabetic individuals. Pak. J. Nutr., 3: 268-272.
CrossRefDirect Link - Sheng, X., Y. Zhang, Z. Gong, C. Huang and Y.Q. Zang, 2008. Improved insulin resistance and lipid metabolism by cinnamon extract through activation of peroxisome proliferator-activated receptors. PPAR Res., Vol. 2008.
CrossRef - Shepherd, P.R. and B.B. Kahn, 1999. Glucose transporters and insulin action-implications for insulin resistance and diabetes mellitus. N. Engl. J. Med., 341: 248-257.
CrossRefPubMedDirect Link - Trayhurn, P. and J.H. Beattie, 2001. Physiological role of adipose tissue: White adipose tissue as an endocrine and secretory organ. Proc. Nutr. Soc., 60: 329-339.
CrossRefPubMedDirect Link - Thorens, B., 1996. Glucose transporters in the regulation of intestinal, renal and liver glucose fluxes. Am. J. Physiol., 270: G541-G553.
CrossRefDirect Link - Vinitha, M. and M. Ballal, 2008. In vitro anticandidal activity of Cinnamomum verum. J. Med. Sci., 8: 425-428.
CrossRefDirect Link - Yaghmoor, S.S. and S.M. Khoja, 2010. Effect of cinnamon on plasma glucose concentration and the regulation of 6-phosphofructo-1-kinase activity from the liver and small intestine of streptozotocin induced diabetic rats. J. Biol. Sci., 10: 761-766.
CrossRefDirect Link - Yamauchi, T., J. Kamon, H. Waki, K. Murakami and K. Motojima et al., 2001. The mechanisms by which both heterozygous peroxisome proliferator-activated receptor γ (PPARγ) deficiency and PPARγ agonist improve insulin resistance. J. Biol. Chem., 276: 41245-41254.
CrossRef - Zahmatkash, M. and M.R. Vafaeenasab, 2011. Comparing analgesic effects of a topical herbal mixed medicine with salicylate in patients with knee osteoarthritis. Pak. J. Biol. Sci., 14: 715-719.
CrossRefDirect Link