
H.B. Han
College of Animal Science and Technology, China Agricultural University, Beijing 100193, People Republic of China
X. Li
College of Animal Science and Technology, China Agricultural University, Beijing 100193, People Republic of China
K. Yu
College of Animal Science and Technology, China Agricultural University, Beijing 100193, People Republic of China
W.Z. Ma
College of Animal Science and Technology, China Agricultural University, Beijing 100193, People Republic of China
Z.C. Cao
College of Animal Science and Technology, China Agricultural University, Beijing 100193, People Republic of China
G.F. An
Zhong Ke Yi Nuo Biological Technology Co., Ltd, Beijing, P.R. China
Z.X. Lian
State Key Laboratory of Agrobiotechnolog, China Agricultural University Beijing 100193, People Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 7 | Page No.: 667-677







ABSTRACT
The purpose of this study was carried out the effects of egg white lysozyme treatment on collagen secreting in mice skin. As one of the most indispensable constituting components of skin, collagen is mainly proved to be responsible for its tensile strength. By far, results have shown that lysozyme, an antibacterial enzyme, plays some positive role in skin burn and yet its function has not been explained in detail in regards to skin condition. In this study, the role of egg white lysozyme in collagen biosynthesis was examined by both dermal fibroblasts in vivo and in vitro. RT-PCR was used to trace the expression levels of collagen in cultured cells and HE and VG staining assessed the collagen content in dermis in vivo. According to present study, egg white lysozyme is discovered to promote the secretion of collagen in fibroblasts treated with a dosage of 10 mg mL-1 around 12 to 48 h. Though, it was found that both he expression levels of col1a1 and col3a1 were increased in present experiment, yet the ratio between the secreted type-I collagen and type-cocollagen remained the same in fibroblasts treated with egg white lysozyme. In addition, higher skin moisture content was observed in the mice group treated with egg white lysozyme in vivo according to the report that egg white lysozyme might enhance the hyaluronan synthase1 (Has1) expression. Moreover, in the histological tests the collagen content also increased after egg white lysozyme treatment in both young and old groups with collagen fibril arranged more orderly and compactly than the control group. Egg white lysozyme promoted collagen biosynthesis and extracellular matrix proteins by fibroblasts in the dermis.
PDF Abstract XML References Citation
Received: December 22, 2010;
Accepted: May 16, 2011;
Published: June 15, 2011
How to cite this article
H.B. Han, X. Li, K. Yu, W.Z. Ma, Z.C. Cao, G.F. An and Z.X. Lian, 2011. Egg White Lysozyme Promoted Collagen Secreting of Dermal Fibroblasts in Mice. Asian Journal of Animal and Veterinary Advances, 6: 667-677.
DOI: 10.3923/ajava.2011.667.677
URL: https://scialert.net/abstract/?doi=ajava.2011.667.677
DOI: 10.3923/ajava.2011.667.677
URL: https://scialert.net/abstract/?doi=ajava.2011.667.677
INTRODUCTION
As one of the most important tissues occupying the largest surface of animal, skin functions through hydro and thermoregulation which prevents the heat and water loss from body, besides, it also wards off any ingression of potentially noxious chemicals or microorganisms. Usually, skin is cataloged into three main layers, from inside to outside which are the subcutaneous fat layer, the dermis and the epidermis (Meglinsky and Matcher, 2001; Olukemi et al., 2005). The dermis is the major component of the skin which is made up of collagen, elastin and glycosaminoglycans, collectively termed the Extracellular Matrix (ECM) (Opitz et al., 2004; Abhijit and Manjushree, 2010). Skin collagen, decreasing with age, has direct relationship with dermal thickness (Varani et al., 2000). The decreasing of collagen content in skin was ascribed to the loss of about 6% of dermal mass per decade (Gogly et al., 1997). Skin fibroblasts are major producer of collagen in dermal (Park et al., 2009). Type-I collagen and type-III collagen are the main formative collagen of skin. Type I collagen is composed of two α1 chains (col1a1) and an α2 chain and type-III collagen is composed of three α1 chains (col3a1) (Wang et al., 2007a). Hyaluronan HA) which is used ro retain skin moisture is synthesized by the HA Synthase (HAS). In dermal fibroblasts, HA synthesis is mainly regulated by Has1 (Akazawa et al., 2010). So it is crucial for dermal fibroblasts to increase the secretion of collagen and HA in order to repair the aged skin as well as to retard this very aging process.
Since lysozyme was discovered by Fleming (Pellegrini et al., 1997), it has been subject to extensive physico-chemical and microbiological investigations (Ranjbar et al., 2006; Fokunang et al., 2005). Lysozymes is ubiquitous enzymes found in biological secretion from many kinds of animals, such as hen egg-white, goose egg-white, human milk, human urine, baboo rat and cow stomach (Pooart et al., 2004). It functions as an antibacterial agent hydrolysing the specific kinds of polysaccharides comprising the cell wall of some Gram-positive bacteria (Ko et al., 2008; Ibrahim et al., 2001). As a protein with a molecular weight around 18000 Da, lysozyme is stable in heated acid solution but heat-labile in alkali (Bera et al., 2007). Lysozyme cleaves at α (1-4) glycosidic linkage connecting the C1 carbon of N-acetyl Muramic Acid (NAM) to the C4 carbon of N-acetyl Glucosamine (NAG) which represent the unit polysaccharide structure of many bacterial cell walls (Vocadlo et al., 2001). Therefore, lysozyme is often seen in antibacterial medicine especially for skin diseases. Murashova has proved that lysozyme had the positive effect on severe complications of skin burns (Alsarra, 2009). It has been reported that lysozyme values has unexplained correlation with skin condition (Bojar and Holland, 2002) . In this study, the function and influences of egg-white lysozyme on skin moisturization and secretion of collagen in dermal fibroblast both in vivo and vitro were thoroughly investigated. Present results suggested that egg white lysozyme not only promotes the secretion of type-I collagen and type-co collagen in dermal fibroblast, it also induces the synthesis of extracellular matrix proteins in the dermis of both male and female mice.
MATERIALS AND METHODS
Isolation and culture of mouse dermal fibroblast: Kunming white fetus mice were obtained from the Animal Center of Academy of Military Medical Sciences. The skin sample was immediately cut into 3-5 mm pieces when the mouse was killed. The epidermis and dermis were separated by floating the skin on 400 U mL-1 hyaluronidase and 600 U mL-1 collagenase I dispasein (Sigma, USA) phosphate-buffered saline at 4°C. overnight. Then the dermis were incubated in collagenase 1 dispase in phosphate-buffered saline at 30°C for 1.5 h. Then the cell suspension were allowed to pass through a 200-eye sieve and centrifuged, washed 3 times with a basic culture medium. Then the fibroblasts were isolated and grown in Dulbecco’s minimum essential medium (DMEM, Gibco, Paisley, PA49RF, Scotland, UK) supplemented with 10% (v/v) fetal bovine serum (FBS, Hyclone, USA). Cells were used for experiments when they were 2 passages and seeded 1x106 cells per 100 mm dish.
Collagen content assay: Hydroxyproline (Hyp) (Sigma, USA) is a post-translational product of proline hydroxylation which is thought to be confined almost exclusively to the connective tissue collagen, where it is present in the Y position of the Gly-X-Y repeating tripeptide (AL-Omireeni et al., 2009; Nemethy and Scheraga, 1986). The dermis fibroblasts were cultured according to the method of Dasu et al. (2003). After 24 h of incubation, cells reached 70-80% of confluence and were treated with egg white lysozyme (Sigma, USA). The egg white lysozyme was dissolved in DMEM at a concentration of 50 mg mL-1, then stored at -20°C for use throughout the study. Then the cells were treated with final gradient concentrations (2.5, 5,10 mg mL-1) in serum-free medium at 37°C in incubator. The supernatants were collected at 12, 24 h and immediately carried out Hyp assay which presented the amount of collagen as the method described by Kivirikko and Myllyla (1982).
Isolation of total RNA and RT-PCR: The cells were harvested after egg white lysozyme treatment with final gradient concentrations (5,10 mg mL-1). Total RNA was extracted using the Trizol (Invitrogen, USA) method according to the manufacturer’s protocol. Total RNA was reverse-transcribed and amplified by PCR with the following primers: β-actin (forward, 5’-GCCCAGAGCAAGAGAGGTATC-3’; reverse,5’-ACCAGAGGCATACAGGGACA-3’;267 bp) ;col1a1(forward,5’-TTGCTTCCCAGATGTCCTATG-3’;reverse,5’-CACCATCCAAACCACTGAAG-3’;348 bp);col3a1(forward,5’-TGCTGGAAAGAATGGGGAGA-3’; reverse, 5’-CACCAGAATCACCCTTGCCTC-3’;215 bp); Has1 (forward,5’-GTCCAAATCTTACTTCCGAG-3’; reverse,5’-TAGTTAGCAGCCAG TTTCTTC-3’;36 3 bp); Hyal1(forward,5’-AACTTTAGTGGAGGCAGCAG-3’; reverse, 5’-TACTGGGGTAAAGGGCATAG-3’;363 bp). After amplification, products were resolved on a 2.0% agarose gel electrophoresis, visualized by EB and photographed, then quantified by the Quantity One 4.5.2 software (Bio-RAD, USA).
Mouse aging model: D-galactose (Sigma, USA) is a sugar which can induce the changes which resembled accelerated aging. Both physiologically and pathologically, the D-galactose-treated animals resemble their aged control counterparts of 16- to 24-months old (Gong and Xu, 1991; Li et al., 1995). We randomly divided 28 kunming white mice (female) into two groups: control group (young group, 16 mice), treatment group (old group, 12 mice). Then the mice of control group (young group) and the treatment group (old group) were given, respectively the following daily intraperitoneal injections for 6 weeks: 0.3 mL physiological saline solution as vehicle control and D-galactose at 80 mg kg-1 weight.
Treatment of egg white lysozyme: The mice of young group were then randomly divided into two groups, so it is with old group. One day before administration, the mice back was depilated about 1.5 cm2 area of right region of spine. Two groups were smeared with blank matrix, which were, respectively chosen from young group (8 mice) and old group (6 mice) and the other two groups were smeared with matrix mixing 200 mg mL-1 egg white lysozyme for 4 weeks. Mice were sacrificed at the end of treatment and skin samples were immediately collected for later experiment.
Skin moisture: Skin moisture was measured by using a Multi Skin Test Center MC750® (Courage+Khazaka electronic GmbH, Koln, Germany) as the method described by the instruction manual. The measurement was taken at the end of treatment of egg white lysozyme. It performed three times at the depilated area of right region of mouse spin, and mean values were obtained.
Hematoxylin-eosin (H&E) and van gieson staining of mice skin sections: The skin samples obtained were fixed in formalin for 24 h before processing and embedding in paraffin wax. Twenty of them, five per group, were reserved. For one skin sample, two consecutive 4 μm thick sections were cut and one of them were stained with haematoxylin and eosin by National Animal TSE Lab, China, the other of them for collagen analysis with Weigert Van Gieson staining, as follows:
Weigher Iron Hematoxylin for 15 min followed by dehydration in Ethyl Alcohol and rinse with distilled water for 10 min. Then stained with Van Gieson for 1-5 min, rapidly placed in 70% Ethyl Alcohol for differentiation and dehydrate in Absolute Alcohol. At last, cleared and mounted in Neutral Balsam. Collagen will be stained pink or deep red; Muscle, Cytoplasm, RBC and Fibrin yellow; Nuclei brownish black.
Statistical analysis: The results were compared and analyzed by using the Statistical Package for Social Science (SPSS version 10.0). A p-value<0.05 was considered to be significant. Results are expressed as Means±Standard Deviation (SD).
RESULTS
Egg white lysozyme promoted collagen synthesis: The hydroxyproline content of collagen preparations from different mammalian and avian sources has been found to be 13.4±0.24% (Neuman and Logan, 1950a). Hydroxyproline may be converted to its equivalent of collagen through multiplication by the factor 7.46 (Neuman and Logan, 1950b). The collagen content may be expressed by the relation: Micrograms hydroxyproline in 1 mL hydrolysate x7.46.
To study the effects of egg white lysoyzme on collagen secretion in dermal fibroblasts, collagen content was estimated indirectly through hydroxyproline content assay. Figure 1a is the standard curve plotted against standard hydroxyproline concentrations varying from 0 to 25 μg mL-1 and the concentrations of hydroxyproline in sample supernatants were thus determined by this standard curve. Figure 1b shows the collagen production in dermal fibroblasts after 12 and 24 h of exposure to egg white lysozyme with dosage varying from 2.5 to 10 mg mL-1.
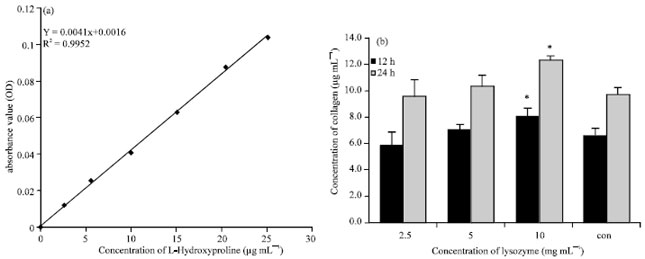 | |
| Fig. 1(a-b): | The effect of egg white lysoyzme on collagen secreted by dermal fibroblasts. (a) Hydroxyproline standard curve shows linear regression of hydroxyproline concentration versus absorbance value. It was generated by plotting the absorbance of the standard solution as a function of its hydroxyproline concentration, as 0, 2.5, 5, 10, 15, 20 and 25 μg mL-1. (b), egg white lysoyzme-induced collagen secreted by dermal fibroblasts. The dermal fibroblasts were treated with different concentrations of egg white lysozyme (2.5, 5, 10, 0 mg mL-1) for 12 and 24 h. The concentration hydroxyproline of in the cell supernatants was determined from the standard curve. The collagen concentration was calculated using the formula: Micrograms hydroxyproline in 1 mL hydrolysate x7.46. Values represent Mean±SD (n = 3), *p<0.05 vs. control cells |
It was then discovered that the biosynthesis of collagen increased significantly in cultured dermal fibroblasts with egg white lysozyme at the dosage of 10 mg mL-1 (p<0.05) (Fig. 1b). Collagen synthesis in fibroblasts treated respectively with dosages of 2.5 and 5 mg mL-1, however, remained almost the same as that in the control cells (Fig. 1b), which clearly shows that egg white lysozyme may fail to increase the collagen secretion in dermal fibroblasts in vitro unless the dosage increase to its threshold around 10 mg mL-1.
Egg white lysozyme Induced expression of type I/type III procollagen: Collagen bundles were 93.35% in the superficial dermis and 89.2% in the reticular dermis (Branchet et al., 1991). Being two important components of extracellular fibers, type III collagen and I are mainly responsible for the tensile strength of the skin. Based on the results above, the expression levels of type I and III procollagen in dermal fibroblasts were also observed after 18 and 40 h egg white lysozyme treatment. By applying semi-quantitative RT-PCR method, it was shown that the expression level of col1a1 increased significantly after egg white lysozyme treatment (p<0.05) (Fig. 2a). Likewise, compared with the blank control, a remarkable increase of expression level of col3a1 was also observed post egg white lysozyme treatment (p<0.05) (Fig. 2b).
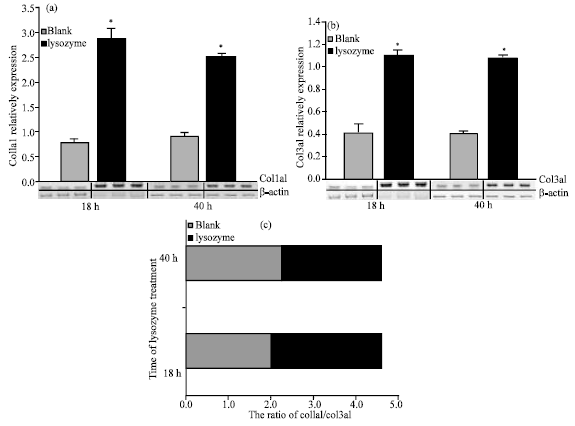 | |
| Fig. 2(a-c): | Egg white lysozyme promote expression levels of col1a1 and col3a1. (a) and (b), after 10 mg mL-1 egg white lysozyme treatment, the expression levels of both col1a1 and col3a1 by dermal fibroblasts were examined by RT-PCR at 18 and 40 h and quantified using the quantity one 4.5.2 soft and normalized with the expression of β-actin. The level of mRNA of untreated control cells was assumed to represent basal expression and was arbitrarily set at 1 relatively expression. The data shown here is Mean±SD from three independent experiments. *p<0.05 vs. control cells. (c), the ratio of col1a1/col3a1 at 18 and 40 h post treatment were basically identical with that of blank treatment |
Furthermore, the expression levels of both col1a1 or col3a1 barely changed at the two time points (18 and 40 h) (p>0.05). Taken all results together, we thus confer that promotion of egg white lysozyme on the expression of col1a1 and col3a1 in dermal fibroblasts is rather irrelevant to the period of treatment.
It was reported that the ratio of collagen type I/type III in the dermis was reduced with age (Kohn and Schnider, 1988; De-Paepe et al., 1987). To further elucidate the role egg white lysozyme played in collagen secretion, the ratio between type I and III procollagen expression levels was calculated using the expression level: ration = expression level (col1a1) / expression level (col3a1). The ratio of col1a1/col3a1 were 2.0 and 2.2 for control group (blank treatment) at 18, 40 h, respectively and 2.6 and 2.4 for lysozyme treatment group. As shown in Fig. 2c, the ratio of col1a1 /col3a1 at 18 h post egg white lysozyme treatment was basically identical with that of blank treatment, which further implies that egg white lysozyme might induce the expression of col1a1 and col3a1, but it never ends in disorder of typeI/type III collagen secretion under the circumstances that the ratio between them remains the same.
Lysozyme increased skin moisturization: Skin moisture content is one of the most parameters used in judging skin condition. Previous researches have proved that water content in the mice skin decreased markedly with aging (Jung et al., 1997). So in order to investigate whether lysozyme will increase the skin moisture, egg white lysozyme was applied to depilated back of mice for a month in our study. Among young mice, significant difference of skin moisture content was found between the treatment group (40.75±2.94) and the blank group (37.88±2.08) (p<0.05) (Fig. 3a). For the aged mice, skin moisture content between the treatment group (38.72±2.32) was remarkably higher than that of blank group (36.00±1.56) (p<0.05) (Fig. 3a). Thus, present results firmly indicated that egg white lysozyme has played a positive role on water-holding capacity of mouse skin.
Except the epidermal function on skin moisture, the dermis contributes to the water-holding capacity through its content of hydrophilic glycosaminoglycans, mainly hyaluronic acid, synthesized in the plasma membrane of fibroblasts and other cells (Hall and Phillps, 2005). With advancing age, the skin cells lose their ability to produce hyaluronic acid (Hotta , 2006) To further clarify the role of egg white lysozyme in increasing skin moisture, the expression levels of hyaluronan synthase1(Has1) and hyaluronidase1(Hyal1) were examined after egg white lysozyme treatment. Semi-quantitative RT-PCR revealed that the expression level of Hyaluronan synthase 1, one of three kinds of hyaluronic acid synthetase genes, increased notably at 18 and 40 h post egg white lysozyme treatment (p<0.05) (Fig. 3b). Hyaluronidase 1 is one of the β-endoglucosidases family members that degrade HA into small fragments (Tanimoto et al., 2007). Compared with control group, no significant difference was found between the expression levels of hyaluronidase 1 at 18 and 40 h post egg white lysozyme treatment (Fig. 3c). Taken together, all these results confirmed that the egg white lysozyme increased the mouse skin moisture content via promoting the biosynthesis of hyaluronic acid in dermal fibroblast.
Egg white lysozyme produced histological changes to mouse skin: Thinning of the dermis, which is closely related to collagen loss, often accompanies aging (Hall and Phillps, 2005). To evaluate the changes and the arrangement of collagen in dermis of mice treated with egg white lysozyme, dermal histological sections were analyzed with Hematoxylin-eosin and Van Gieson staining. Figure 4 (HE) shows the disorder and rarefaction of the collagen fibres (red staining) in aged mice compared with young mice.
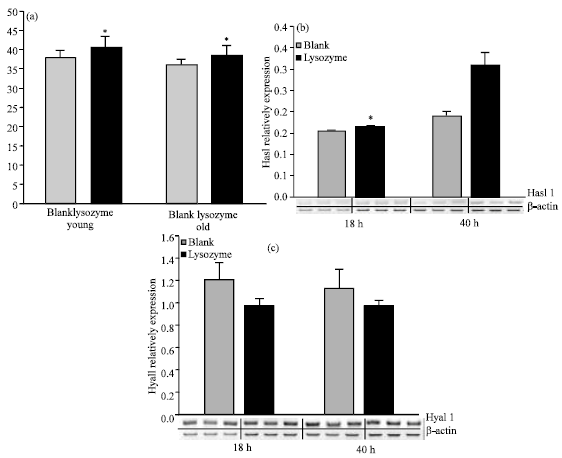 | |
| Fig. 3: | Effect of egg white lysozyme on skin moisture. (a) mouse skin moisture was measured by using a Multi Skin Test Center MC750 with egg white lysozyme treatment for a month. Egg white lysozyme significantly increased the skin moisture in both young and old mice,*p<0.05. b and c, after 10 mg mL-1 egg white lysozyme treatment, the expression levels of both Has1 and Hyal1 by dermal fibroblasts were examined by RT-PCR at 18 and 40 h and quantified using the quantity one 4.5.2 soft and normalized with the expression of β-actin. (b) There was remarkable difference of expression level of Has1 between egg white lysozyme treatment and blank treatment at both 18 and 40 h, *p<0.05. (c) No difference of expression level of Hyal1 was displayed between egg white lysozyme treatment and blank treatment, p>0.05 |
These photos clearly show that age-related alteration manifested by dermis thinning is often accompanied with collagen losing Fig. 4 (V.G), so it is quite feasible and worth while to conduct some researches on the change of skin mositurization in mice aging models induced by D-galactose after egg white lysozyme treatment. Figure 4 (V.G) shows that collagen content increased post egg white lysozyme treatment in either young or aged group., and the collagen fibril became more orderly and compactly arranged in aged mice with egg white lysozyme treatment compared with the control group Fig. 4 (HE).
Although the effect of egg white lysozyme on dermal fibroblasts has previously been demonstrated, while our results now have provided direct evidences that egg white lysozyme plays a significantly positive role on skin condition by inducing the secretion of collagen and hyaluronic acid in dermal fibroblasts.
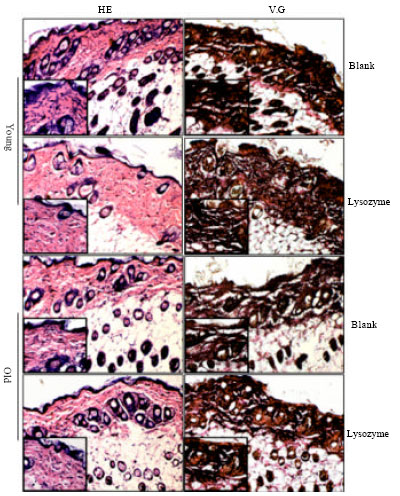 | |
| Fig. 4: | Effect of egg white lysozyme on mouse skin. The skin samples obtained after smearing egg white lysozyme on mouse back skin for a month. Then the histological sections stained with Hematoxylin-eosin (HE) and Van Gieson (V.G). By V.G staining, collagen will be stained pink or deep red. Mice were divided into two groups. One group was intraperitoneal injections 0.3 mL physiological saline solution (young group) for 6 weeks and the other was injections D-galactose at 80 mg kg-1 weight (old group). Two groups were smeared with blank matrix, which were, respectively chosen from young group and old group and the other two groups were smeared with matrix mixing 200 mg mL-1 egg white lysozyme for 4 weeks (x100). On the left lower quadrant was magnifying of the same section (x200) |
DISCUSSION
There were some studies of collagen secreting of fibroblasts in different skin condition. Estradiol treatment stimulated collagen production in sun-protected skin (Rittie et al., 2008). Cross-linked hyaluronic acid increased the collagen synthesis in photo damaged skin (Wang et al., 2007b). It has so far been known little about the relationship between lysozyme and skin collagen secreting.
Types I and III collagen are most significant components of skin structure. Accompany with aging, the collagen content steadily decreases (Broniarczyk-Dyla and Joss-Wichman, 2001). Present results demonstrated that egg white lysozyme efficiently induced collagen biosynthesis in dermal fibroblasts and promoted collagen content in mouse skin. So we presumed that egg white lysozyme, in some extent, could delay skin aging.
We detected hydroxyproline content increased in dermal fibroblasts supernatants with egg white lysozyme treatment. This result showed two possibilities: (a) egg white lysozyme promoted fibroblasts proliferation without changing their collagen secreting, (b) egg white lysozyme promoted collagen secretory volume per cell. Furthermore, cell proliferation assay revealed no significant difference between egg white lysozyme treatment and blank treatment (data not shown). Combined with result of increasing expression level of col1a1 and col3a1, it was prove that egg white lysozyme had definite effects on promoting collagen biosynthesis.
Ratio of types I and III collagen has different physical properties in different tissue (Wei et al., 1999). Most pliable tissues (skin, intestine, blood vessels, lung) have a I: III ratio of 2-3 to 1 (Hance and Crystal, 1977). Any fluctuate of the ratio will cause some changes of skin, such as it increased in hypertrophic scars (Friedman et al., 1993) and it decreased in inguinal hernia (Klinge et al., 2000). Present study shows that egg white lysozyme induced expression level of typeI, III and its ratio within normal accelerational range. It disclosed that egg white lysozyme can be used as collagen promoter without negative effect because it does not alter the expression property of typeI/type III .
Above results can only demonstrate the effect of egg white lysozyme on fibroblasts in vitro. Therefore, it is of critical importance to clarify whether egg white lysozyme has the same effect on mice dermis in vivo. In order to study the effect on mice at different age, aging mice model was induced by D-galactose. By using HE and VG staining technique, we found increased collagen content and more orderly collagen arrangement with egg white lysozyme treatment in the dermis of either young and old mice. Otherwise, data through skin moisture assay, in both young and old mice, revealed that smearing egg white lysozyme on skin for a while can promote the skin moisture which may directly or indirectly influence by expression alterations of hyaluronic acid in the dermis.
In conclusion, egg white lysozyme can raise the moisture and collagen content of mouse dermis though induced the expression level of collagen and hyaluronic acid. The skin condition was improved in some extent at different age with lysozyme treatment. Finding of this study was the first time to reveal the anti-aging or decreasing wrinkle effects of lysozyme.
ACKNOWLEDGMENTS
We thank Mr. Lu QZ for careful reading the manuscript and for his valuable suggestions. This work was funded by the National Key Basic Research ‘973’ Program of China (2006 B1021 00).
REFERENCES
- Al-Omireeni, E.A., N.J. Siddiqi and A.S. Alhomida, 2009. Effect of different doses of sodium fluoride on varous hydroxyproline fractions in rat kidneys. Kidney Res. J.
Direct Link - Bera, A., R. Biswas, S. Herbert, E. Kulauzovic, C. Weidenmaier and A.G. Friedrich, 2007. Peschel, influence of wall teichoic acid on lysozyme resistance in Staphylococcus aureus. J. Bacteriol., 189: 280-283.
CrossRef - Bojar, R.A. and K.T. Holland, 2002. Review: The human cutaneous microflora and factors controlling colonisation. World J. Microbiol. Biotechnol., 18: 889-903.
CrossRef - Branchet, M.C., S. Boisnic, C. Frances, C. Lesty and L. Robert, 1991. Morphometric analysis of dermal collagen fibers in normal human skin as a function of age. Arch. Gerontol. Geriat., 13: 1-14.
PubMed - Broniarczyk-Dyla, G. and E. Joss-Wichman, 2001. Ageing of the skin during menopause. J. Eur. Acad. Dermatol. Venereol., 15: 494-495.
CrossRef - De-Paepe, A., A. Nicholls, P. Narcisi, F. De-Keyser and J. Quatacker et al., 1987. Ehlers-Danlos syndrome type I: A clinical and ultrastructural study of a family with reduced amounts of collagen type III. Brit. J. Dermatol., 117: 89-97.
CrossRef - Fokunang, C.N., K.A. Watson, A. Purvis and E.A. Tembe-Fokunang, 2005. Crystallization and structural studies of lysozyme from hen egg white, using vapour diffusion techniques. Biotechnology, 4: 341-346.
CrossRefDirect Link - Friedman, D.W., C.D. Boyd, J.W. Mackenzie, P. Norton, R.M. Olson and S.B. Deak, 1993. Regulation of collagen gene expression in keloids and hypertrophic scars. J. Surg. Res., 55: 214-222.
CrossRef - Gogly, B., G. Godeau, S. Gilbert, J.M. Legrand, C. Kut, B. Pellat and M. Goldberg, 1997. Morphometric analysis of collagen and elastic fibers in normal skin and gingiva in relation to age. Clin. Oral Investigations, 1: 147-152.
CrossRef - Hall, G. and T.J. Phillps, 2005. Estrogen and skin: The effects of estrogen, menopause and hormone replacement therapy on the skin. J. Am. Acad. Dermatol., 53: 555-568.
CrossRef - Hance, A.J. and R.G. Crystal, 1977. Rigid control of synthesis of collagen Types I and III by cells in culture. Nature, 268: 152-154.
CrossRef - Hotta, T., 2006. Understanding the anatomy when using dermal fillers enhances patient safety. Plast. Surg. Nurs., 26: 149-151.
PubMedDirect Link - Jung, J.W., S.H. Chab, S.C. Lee, I.K. Chun and Y.P. Kim, 1997. Age-related changes of water content in the rat skin. J. Dermatol. Sci., 14: 12-19.
CrossRef - Klinge, U., Z.Y. Si, H. Zheng, V. Schumpelick, R.S. Bhardwaj and B. Klosterhalfen, 2000. Abnormal collagen i to iii distribution in the skin of patients with incisional hernia. Eur. Surg. Res., 32: 43-48.
CrossRef - Ko, K.Y., A.F. Mendonca and D.U. Ahn, 2008. Effect of ethylenediaminetetraacetate and lysozyme on the antimicrobial activity of ovotransferrin against Listeria monocytogenes. Poult. Sci., 87: 1649-1658.
CrossRef - Li, W.B., F. Wei, M. Fan, J.L. Zhang and B.L. Zhang et al., 1995. Mimetic brain aging effect induced by D-galactose in mice. Chinese J. Pharm. Toxicol., 9: 93-95.
Direct Link - Meglinsky, I.V. and S.J. Matcher, 2001. Modelling the sampling volume for skin blood oxygenation measurements. Med. Biol. Eng. Comput., 39: 44-45.
CrossRef - Dasu, M.R., R.E. Barrow, M. Spies and D.N. Herndon, 2003. Matrix metalloproteinase expression in cytokine stimulated human dermal fibroblasts. Burns, 29: 527-531.
PubMed - Nemethy, G. and H.A. Scheraga, 1986. Stabilization of collagen fibrils by hydroxyproline. Biochemistry, 25: 3184-3188.
CrossRefPubMedDirect Link - Neuman, R.E. and M.A. Logan, 1950. The determination of hydroxyproline. J. Biol. Chem., 184: 299-306.
PubMedDirect Link - Neuman, R.E. and M.A. Logan, 1950. The determination of collagen and elastin in tissues. J. Biol. Chem., 186: 549-556.
Direct Link - Olukemi, O.A., J.M. Oluseyi, I.O. Olukemi and S.M. Olutoyin, 2005. The use of selected Nigerian natural products in management of environmentally induced free radical skin damage. Pak. J. Biol. Sci., 8: 1074-1077.
CrossRefDirect Link - Opitz, F., K. Schenke-Layland, T.U. Cohnerta, B. Starcherb, K.J. Halbhuberc, D.P. Martind and U.A. Stocka, 2004. Tissue engineering of aortic tissue: Dire consequence of suboptimal elastic fiber synthesis in vivo. Cardiovascular Res., 63: 719-730.
CrossRef - Park, H.J., D.H. Cho, H.J. Kim, J.Y. Lee and B.K. Cho et al., 2009. Collagen synthesis is suppressed in dermal fibroblasts by the human antimicrobial peptide LL-37. J. Investigative Dermatol., 129: 843-850.
CrossRef - Pooart, J., T. Torikata and T. Araki, 2004. The primary structure of a novel goose-type lysozyme from rhea egg white. Biosci. Biotechnol. Biochem., 68: 159-169.
CrossRef - Pellegrini, A., U. Thomas, N. Bramaz, S. Klauser, P. Hunziker and R. von Fellenberg, 1997. Identification and isolation of a bactericidal domain in chicken egg white lysozyme. J. Applied Microbiol., 82: 372-378.
PubMedDirect Link - Rittie, L., S. Kang, J.J. Voorhees and G.J. Fisher, 2008. Induction of collagen by estradiol: Difference between sun-protected and photodamaged human skin in vivo. Arch. Dermatol., 144: 1129-1140.
PubMed - Varani, J., R.L. Warner, M. Gharaee-Kermani, S.H. Phan and S. Kang et al., 2000. Vitamin A antagonizes decreased cell growth and elevated coallagen-degrading matrix metalloproteinases and stimulates collagen accumulation in naturally aged human skin. Soc. Investigative Dematol., 114: 480-486.
CrossRef - Tanimoto, K., Y. Nakatani, N. Tanaka, M. Ueki and T. Yanagida et al., 2007. Inhibition of the proliferation of human periodontal ligament fibroblasts by hyaluronidase. Arch. Oral Biol., 53: 330-336.
CrossRef - Vocadlo, D.J., G.J. Davies, R. Laine and S.G. Withers, 2001. Catalysis by hen egg-white lysozyme proceeds via a covalent intermediate. Nature, 412: 835-838.
CrossRef - Wang, Q., Z. Peng, S. Xiao, S. Geng, J. Yuan and Z. Li, 2007. RNAi-mediated inhibition of COL1A1 and COL3A1 in human skin fibroblasts. Exp. Dermatol., 16: 611-617.
CrossRef - Wang, F., L.A. Garza, S. Kang, J. Varani, J.S. Orringer, G.J. Fisher and J.J. Voorhees, 2007. In Vivo stimulation of de novo collagen production caused by cross-linked hyaluronic acid dermal filler injections in photodamaged human skin. Arch. Dermatol., 143: 155-163.
PubMed - Wei, S., L.T. Chow, I.O. Shum, L. Qin and J.E. Sanderson, 1999. Left and right ventricular collagen type I/III ratios and remodeling post-myocardial infarction. J. Card Fail., 5: 117-126.
Direct Link - Ranjbar, B., S. Afshar, A. Kakanejadifard, K. Khajeh, H. Naderi-Manesh, L. Hassani and N. Alizadeh, 2006. Effects of glyoxime and dichloroglyoxime on lysozyme: Kinetic and structural studies. Asian J. Biochem., 1: 153-161.
CrossRefDirect Link - Alsarra, I.A., 2009. Chitosan topical gel formulation in the management of burn wounds. Int. J. Biol. Macromol., 45: 16-21.
CrossRef - Kivirikko, K.I. and R. Myllyla, 1982. Post-translational enzymes in the biosynthesis of collagen: Intracellular enzymes. Method Enzymol., 82: 245-304.
CrossRef - Sahasrabudhe, A. and M. Deodhar, 2010. Anti-hyaluronidase, anti-elastase activity of Garcinia indica. Int. J. Bot., 6: 299-303.
CrossRefDirect Link