
R. Ramesh
Noorul Islam College of Arts and Science, Kumaracoil-629180, India Centre of Advanced Study in Marine Biology (Annamalai University), Parangipettai-608 502, India
S. Ravichandran
Noorul Islam College of Arts and Science, Kumaracoil-629180, India Centre of Advanced Study in Marine Biology (Annamalai University), Parangipettai-608 502, India
Trends in Applied Sciences Research
Year: 2008 | Volume: 3 | Issue: 2 | Page No.: 189-195







ABSTRACT
Scanning Electron Microscopic studies on the radula of Turbo brunneus was made to study the anatomy of feeding apparatus. Feeding experiment was carried out to find out algal preference and rate of consumption. Study on the radula of Turbo brunneus possesses a large central rachidian tooth, five lateral teeth and many marginal teeth. The inner marginal situated towards the lateral teeth are formed of a strong extended primary cusp and a short secondary cusp along outer margin. Observation of T. brunneus in its natural habitat shows that they are mainly found in thickly grown algal fronds of Rhodophytes such as Hypnea sp. Ceramium miniatum and Gracilaria sp. Analysis on the gut content shows that the species of algae found in the gut vary during different months. The Rhodophytes dominate during most of the months. Crustose coralline algae always form a considerable part in the gut content during all seasons. Turbo brunneus consumed most of the algae fed to them, Ceramium miniatum and Hypnea muciformis are found to be higher with 3.67% and 3.46% b.wt. day-1, respectively.
PDF Abstract XML References Citation
How to cite this article
R. Ramesh and S. Ravichandran, 2008. Feeding Biology with Reference to Algal Preferance and Scanning Electron Microscopy Studies on the Radula of Turbo brunneus. Trends in Applied Sciences Research, 3: 189-195.
URL: https://scialert.net/abstract/?doi=tasr.2008.189.195
URL: https://scialert.net/abstract/?doi=tasr.2008.189.195
INTRODUCTION
The gastropoda are remarkable for their feeding mechanisms and for the wide variety of food they subsist. The feeding habits and food preference do not form a distinctive part of a particular group. Thus, the prosobranchia in particular, displays almost every conceivable food preference. Among prosobranchs, the Archaeogastropoda show rather primitive feeding habits such as browsing and grazing of algae.
Raffaelli (1976) observed that the intertidal browsing gastropods appear unselective in their diets when feeding within a particular zone on the shore. The evolutionary success is attributable in large measure to the structural and functional plasticity of feeding apparatus. Indeed, Purchon (1977) stated that the adaptive radiation of the gastropoda is principally based on adaptations of the buccal mass and radula to meet the problems faced in feeding in various ways on a wide variety of food substances. Various authors relate radular morphology of a similar group of molluscs to similar feeding habits.
There are only few studies related to the feeding biology of Turbinids from different parts of the world. Turbo brunneus despite its economic value and edibility has not been considered for aquaculture. If T. brunneus is considered and developed for aquaculture, knowledge about the algal preference, feeding apparatus, consumption rates are essential. Such data would provide a basis for detailed feeding and growth studies.
MATERIALS AND METHODS
SEM Study on the Radula
For the Scanning Electron Microscopic studies on the radula of T. brunneus, small cut was made on the dorsal surface of the head and the radula was exposed. Then dissection was done till the end of the snout and the radula along with the tongue like odontophore was detached from the muscle fibers attached to it. The separated radula was washed in mild alkaline solution and a sort brush is used to remove adhering tissues. The radula was then washed and dehydrated by using a serious of alcohol grades (30, 50, 70, 90 and 95%). The dehydrated radula was brought to the next step for coating thereby making them suitable for SEM observation.
Principally coating of the radula was done by placing the specimen in a vacuum evaporator, pumping the chamber down to a vacuum level equivalent to that in the SEM specimen chamber and heating the conductive metal (gold palladium) to the point that it vapourises. A small part of the cloud of metallic vapour will hit and stick to the surface of the object, so as to provide a thin metallic coating over it. Continuous tilting and rotation of the specimen is facilitated so that the metallic vapour covers all the sides of the specimen.
The radula was mounted on the SEM stub, thereby it could be tilted in any angle for easy viewing. The radula when drying in vacuum became folded and shrunk, so Duco cement was introduced for mounting the radula on the SEM slub.
Field Observation of T. brunneus
Field observation was made during early hours at low tide or before it. The feeding association of T. brunneus with different kinds of algae were observed and noted down. Different groups of algae found in the area were identified and classified in respective divisions.
Gut Content Analysis
Twenty snails of different sizes were collected monthly (May 2002 to April 2003). The snails, which were found actively feeding, were collected, killed immediately and preserved in formalin to prevent further digestion of the ingested algal matter. They were taken to the laboratory, dissected and complete contents of the digestive tract were spread evenly in a Petri dish. A little quantity of distilled water was added to dilute the contents. Another Petri dish was taken and a graph sheet was placed under it to divide whole area into many divisions. The diluted gut contents were then pipetted out in small drops over the Petri dish with divisions formed by graph sheet. Then each drop was analysed for algal material. Algal filaments which were intact and identifiable were counted and their percentage from the total species of algae and detritus was calculated. Algal forms that were fully digested could not be recognized which was included under detritus. Detritus also included small stone bits, spicules of sponges, appendages of small crustaceans and other unidentifiable vegetable or non-vegetable matter. The different forms of algae identified were place under respective division.
Consumption Rate of Different Algal Types by T. brunneus
New animals were collected for each experiment to avoid pseudo-replication and to ensure that animals were not affected by long-term laboratory exposure. For each experiment, snails in the size range of 35 to 50 mm. Ten in numbers were placed in 10 aerated plastic troughs of 5 L capacity and were allowed to acclimatize for six days prior to analyses. During this period animals were starved to empty their gut contents and also to accept the food readily. The sea water was replaced daily with freshly filtered one.
Each day the animals were provided with an excess of fresh algae upon which they could feed and libitum. The amount offered was blotted damp-dry and weighed to the nearest 0.1 mg. The following day, uneaten food was removed, blotted damp dry and reweighed. This procedure was followed for five consecutive days for each alga. The mean amount consumed was calculated for the five days for each snail and used to determine the daily food consumption rates and specific consumption rates. Daily food consumption rate was calculated using the formula (Foster and Hodgson, 1998).
Cg = F – R
where:
| Cg | = | Consumption (g) |
| F | = | Initial food weight |
| R | = | Weight of food remaining after feeding. |
Daily food consumption (Cg) was then standardized for body weight and is expressed as a daily specific consumption rate using the formula.
where:
| C% b.wt | = | Animal food consumption (percentage of body weight) |
| Cg | = | Daily food consumption |
| w | = | Animal weight (g). |
RESULTS
SEM Study on the Radula
The radula of Turbo brunneus possesses a large central rachidian tooth, five lateral teeth and many marginal teeth (Fig. 1a). The rachidian tooth situated in the middle of the row is large, broad and is highly cusped (Fig. 1b). It is very much wider than the adjacent lateral teeth (Fig. 1c). The rachidian is followed on each side by five strongly cusped, progressively narrower laterals with he expanded bases overlapping one another. The laterals from each side of the row are in turn flanked by numerous marginal in outer border. The inner marginal situated towards the lateral teeth are formed of a strong extended primary cusp and a short secondary cusp along outer margin. While the marginal that are projecting on the outside (i.e.,) next to the outer edge of inners marginal have a single narrow serrate cusp (Fig. 1d, e).
Field Observation of Turbo brunneus
Different species of algae found in the study area are classified as:
Chlorophyta
| • | Enteromorpha prolifera |
| • | Ulva lactuca |
| • | U. reticulate |
| • | Chaetomorpha torta |
| • | Chaetomorpha linnoides |
| • | Caulerpa racemosa |
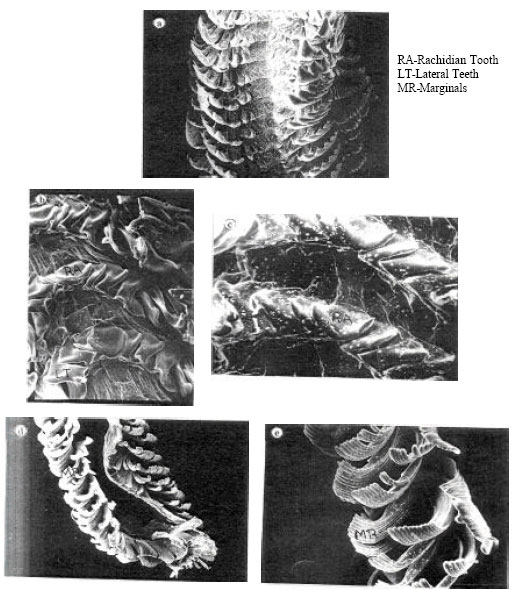 | |
| Fig. 1: | (a) SEM photograph showing the mid-region of the radula x 30, (b) SEM photograph showing the rachidian and lateral x 140 teeth, (c) A close view of the rachidian tooth x 160, (f) The anterior of the radula with its marginals x 25 and (e) A close view of the outer marginals x 130 |
Phaeophyta
| • | Padina gymnospora |
| • | P. tetrastramatica |
| • | Sargassum tenerrium |
| • | S. wightii |
Rhodophyta
| • | Acanthophora spicifera |
| • | Hypnea musciformis |
| • | H. valentiae |
| • | Centroceras clavulatum |
| • | Spyridia filamentosa |
| • | Ceramium miniatum |
| • | Gracilaria edulis |
| • | G. crassa |
| • | Polysiphonia tuticorinensis |
| • | Champia parvula |
Observation of T. brunneus in its natural habitat shows that they are mainly found in thickly grown algal fronds of Rhodophytes such as Hypnea sp. Ceramium miniatum and Gracilaria sp. Next to it, they are found in Chlorophytes such as Ulva lactuca and Chaetomorpha torta. No association was found with Phaeophytes. Many snails are also found scrapping over the crustose coralline algae formed over dead coral piece and their occurrence is high when other filamentous algae are not found in good quantities.
Snails are found feeding actively during early hours, the slow undulating movement of the head along with tentacles indicating the feeding activity. In an exposed shore, the snails accommodate it in small crevices or inside thickly grown algal fronds and feeding are continuous with the available matter. The snail hides itself from the sunlight during day hours and scraps on the detritus material in the vicinity of them. When the feeding snail is forcibly separated from their attachment, small pieces of algal filaments or scraps of coral bits with crustose coating are found in between the folds of foot and also in the snout region. This slows that the snails are actively feeding on the algal forms and the foot plays a major role in holding the algal filaments while the snail feeds on it.
Gut Content Analysis
Analysis on the gut content shows that the species of algae found in the gut vary during different months. The Rhodophytes dominate during most of the months. The minimum qualities, but their mean percentage increases during November and December months. Crustose coralline algae always form a considerable part in the gut content during all seasons (Table 1).
Consumption Rates of Different Algae Types
Turbo brunneus consumed most of the algae fed to them, Ceramium miniatum and Hypnea muciformis are found to be higher with 3.67 and 3.46% b.wt. day-1, respectively. Gracilaia edulis is consumed more than G. crassa.
| Table 1: | Mean percentage of different algae and detritus found in the guts of T. brunneus during different months of observation (± SD) |
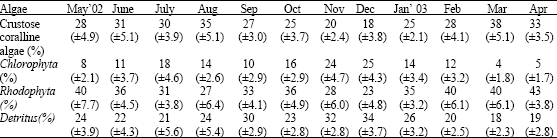 | |
| Table 2: | Daily consumption rates of adult T. brunneus fed with different algae |
 | |
The consumption rate is found to be least with Chaetomorpha torta 1.28% b.wt day-1. No consumption was observed with algae such as Polysiphonia tuticorinensis and Acanthophora spicifera (Table 2).
DISCUSSION
The radula of T. brunneus is rhiphidoglossan type and is similar to that of Tricolia pullus as explained by Robertson (1958) and Fretter and Graham (1962). Hickman (1976) suggested that the rhipidoglossan gastropod has a wider range of food resources. The rhipidoglossate radula of T. brunneus is used for feeding different kinds of algae such as crustose coralline (Lithothamnium sp.), filamentous (Hypnea sp. and Ceramium sp.) and algae with thick fronds (Ulva sp.). But, Steneck and Walting (1982) and Hawkins and Hartnoll (1983) stated that the radulae of Turbo sp. can only serve as brooms capable of sweeping microalgae and filamentous algae and also they are unable to remove large quantities of algal material because of their weak musculature. The present observation showed that T. brunneus is capable of feeding large quantities of crustose coralline and other algal forms as noted by Worthington and Weather (1989) and Foster and Hodgson (1998). Also, Steneck and Walting (1982) observed the rachidian tooth to be small and its size is same as that of laterals in a rhipidoglossate radula. But in T. brunneus the rachidian tooth is larger and broader than the laterals. But, Raffaelli (1985) observed that the algal categories taken by gastropods might be more dependent on the algal availability than radular morphology.
The gut content analysis of T. brunneus shows that crustose coralline algae in considerable quantity as observed in T. udulatum by Worthington and Fair Weather (1989). The coralline algae, although nutritionally poor, are consumed by Turbo sp. and other vetigastropods, (Clarke 1988; Worthington and Weather, 1989; Wood and Buxton, 1996; Foster and Hodgson, 1998).
Rao (1937) and Yamaguchi and Kikutani (1989) observed Hypnea sp. as commonly occurring algae in the gut of Turbo sp. In the present study also Hypnea sp. formed one of the major constituents of Rhodophytes found in the gut of T. brunneus. The other algal forms in the gut mainly depend on their availability and growth in different seasons. Worthington and Fair Weather (1989) and Chapman (2003) observe that Turbo is essentially an opportunistic feeder that eats a wide range of algae, depending on their availability.
The consumption rate of T. brunneus ranges between 1.28 to 3.67% body wt day-1, depending on the algae. The feeding rate was high (3.67 body wt day-1) with Ceramium miniatum and low (1.28 body wt day-1) Chaetomorpha torta. Foster and Hodgson (1998) observed it to vary between 1.06 and 9.50% of its body wt day-1. The values observed in T. brunneus are within the range reported for other grazing gastropods (Graham, 1973; Clarke, 1988; Dixon, 1992; Catterall et al., 2001).
Detritus formed by vegetable and non-vegetable matter was taken by T. brunneus during all seasons. Raffaelli (1985) explained that even when algae were ingested in large amounts, the additional use of detritus as a food source by the gastropods could not be ruled out. The present study indicates that the detritus content may have been eaten inadvertently along with algal food, or it may offer an additional resource to species traditionally viewed as herbivores.
REFERENCES
- Catterall, C.P., I.R. Poiner and C.J. O'Brien, 2001. Long-term population dynamics of a coral reef gastropod and responses to disturbance. Aust. Ecol., 26: 604-617.
Direct Link - Chapman, M.G., 2003. The use of sandstone blocks to test hypotheses about colonization of intertidal boulders. J. Mar. Biological Assoc. UK., 83: 415-423.
Direct Link - Foster, G.G. and A.N. Hodgson, 1998. Consumption and apparent dry matter digestibility of six intertidal macroalgae by Turbo sarmaticus (Mollusca: Vetigastropoda: Turbinidae). Aquaculture, 167: 211-227.
CrossRef - Graham, A., 1973. The anatomical basis of function in the buccal mass of prosobranch and Amphineuran molluscs. J. Zool. Lond., 169: 317-348.
CrossRefDirect Link - Hawkins, S.J. and R.G. Hartnoll, 1983. Grazing of intertidal algae by marine invertebrates. Oceanogr. Mar. Biol. Annu. Rev., 21: 195-282.
Direct Link - Steneck, R.S. and L. Watling, 1982. Feeding capabilities and limitation of Herbivorous molluscs: A functional group approach. Mar. Biol., 68: 299-319.
CrossRefDirect Link - Wood, A.D. and C.D. Buxton, 1996. Aspects of the biology of the abalone Haliotis midae (Linne, 1758) on the east coast of South Africa. 1. Feeding biology. S. Afr. J. Mar. Sci., 17: 61-68.
CrossRefDirect Link