
Nabil S. Farag
Plant Pathology Research Institute ARC, Giza, Egypt
Ahmad A. Gomah
Plant Pathology Research Institute ARC, Giza, Egypt
Naglaa M.A. Balabel
Potato Brown Rot Project, ARC, Dokki, Egypt
Plant Pathology Journal
Year: 2010 | Volume: 9 | Issue: 2 | Page No.: 73-78







ABSTRACT
The multiplex-PCR showed a high degree of reliability to detect the following pathogenic bacteria of potato i.e.,: Ralstonia solanacearum, Clavibacter michiganensis ssp. sepedonicus and Erwinia carotovora ssp. atroseptica. The possible carryover of tuber constituents in extracts, even at high concentration, did not affect the positivness of the reaction differentiating the three pathogens in combination. Antibiotics used in certain bacteriological selective media, varied in their effect on multiplex-PCR results, regardless of their mode (s) of action. Polymyxin B and Bacitracin (polypeptide antibiotics) at the used concentrations caused complete inhibition of the reaction. Similar results were obtained for Trimethoprim (dihydroxyfolate reductase). Other antibiotics as Penicillin G (β-lactam), Chloramphenicol (acetamide), Amphotericin (polyene antibiotic) and Nalidixic acid (quinolone) did not affect the resolving potential of the reaction. Other components of the selective media as TTC (tri-phenyltetrazolium chloride) and CV (rosaniline chloride) did not inhibit the reaction as well. More investigations are needed to study the least concentration of polypeptide and other groups of antibiotics that may influence the multiplex-PCR reaction, hence gives false negative results. The Multiplex-PCR inhibition with certain chemical(s) carryover may be important in other fields of PCR applications as in forensic investigations.
PDF Abstract XML References Citation
How to cite this article
Nabil S. Farag, Ahmad A. Gomah and Naglaa M.A. Balabel, 2010. False Negative Multiplex PCR Results with Certain Groups of Antibiotics. Plant Pathology Journal, 9: 73-78.
DOI: 10.3923/ppj.2010.73.78
URL: https://scialert.net/abstract/?doi=ppj.2010.73.78
DOI: 10.3923/ppj.2010.73.78
URL: https://scialert.net/abstract/?doi=ppj.2010.73.78
INTRODUCTION
Multiplex-PCR is a variant of PCR which enabling amplification of many targets, concerned, in one reaction by using more than one pair of primers. Multiplex assays can be tedious and time-consuming since it requires lengthy optimization procedures, though it is a cost saving technique in many diagnostic laboratories (Elnifro et al., 2000). The technique is subject, however, to certain difficulties related in principal to the availability of primers for various plant pathogens (Martin et al., 2000; Schaad and Fredrick, 2002); the formation of primer dimers (Jannine et al., 1997) and the carryover of certain inhibitory factors in the samples (Rossen et al., 1992; Wilson, 1997). The most distinguished inhibitory factors may be certain plant polysaccharides (Demeke and Adams, 1992) and humic acid in plant and soil (Tsai and Olson, 1992; Watson and Blackwell, 2000). Thus, the optimization of reaction conditions should aim to minimize such non-specific reactions and to avoid false results (De Lomas et al., 1992).
This research was undertaken to study the effect of potato tuber extracts and the antibiotic carryover from laboratory selective media of bacterial plant pathogens, on the sensitivity of multiplex -PCR results.
MATERIALS AND METHODS
Extraction Potato tissue: Composite samples, each containing 200 tubers, were processed as described by Pastrik (2000). After washing tubers in running tap water, to free from adherent soil, the stolon ends were removed and cores of the vascular tissues were taken in 50 mL phosphate buffer (pH 7.0). Rotary shaking was controlled (100 rpm) at 24°C for 2 h. Cooling centrifugation at 10°C and 10.000 rpm was run for 10 min, then the supernatant was carefully discarded. The resulting pellet was resuspended in 1 mL pellet buffer (pH 7.2).
DNA extraction: Cultures of the bacterial pathogens were harvested separately in the late log phase. For the isolation of the genomic DNA, a loopful of bacterial culture was suspended in 1 mL of 0.01 M phosphate buffer saline (PBS g L-1: 2.7 Na2 HPO4. 12H2O, 0.4NaH2PO4 2H2O, 8.0 NaCl, 1000 mL distilled water; pH 7.2). Cell lysis was made by heating to 96°C for 4 min, then cooled on ice for 10 min. Aliquots of 2 μL of the extract were added to the reaction mixtures (Seal et al., 1993).
Source of bacterial pathogens: Bacterial cultures of the pathogens in concern and their origins are shown in Table 1, the ring rot pathogen (BRR) was grown on yeast extract glucose mineral salt agar (YGMA) at 20°C (Anonymous, 1993). The brown rot pathogen (BR) on casamino peptone glucose agar at room temperature (Kelman, 1954). The blackleg pathogen (BL) was verified on Crystal Violet Pectate (CVP) medium at 20°C (Cupples and Kelman, 1974).
Primers: The sequence of primers used in this study is shown in Table 2, the primers were provided by OPRON, USA
Procedures: PCR amplification was run by PTC-200 Peltier (USA) thermal cycler for primers listed in Table 2, the reaction mixture (25.0 μL) consisted of 12.5 μL Qiagen Taq PCR master mix, 1 μL of atroseptica primers (0.1 dilution) in sterile UPW; 1 μL of each of solanacearum primers and 2 μL of sepedonicus primers along with 2.0 μL of DNA extract. Supplement of antibiotics and dyes were added separately as 2.5 μL solution, to the reaction mixture and being replaced by the same quantity of sterile UPW in the check treatment.
| Table 1: | Potato bacterial pathogens, source of cultures and disease produced |
 | |
| *PD: Plant protection service; Wageningen NL. **Potato brown rot project, Dokki, Egypt | |
| Table 2: | Sequence of primers used with Clavibacter michiganensis ssp. sepedonicus, Erwinia carotovora ssp. atroseptica and Ralstonia solanacearum, race 3 |
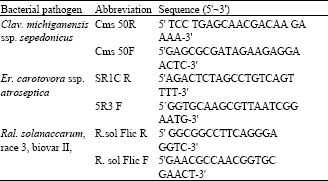 | |
The initial denaturation was made at 96°C/3 min, followed by 35 reaction cycles at 94°C/30 sec, then 52°C/30 sec and 72°C/30 sec. The mixture was then kept at 72°C/10 min. and stored at 4°C. Aliquots (13 μL) of the reaction mixture were resolved by electrophoresis in agarose gel (2%) and DNA fragments were visualized by ethidium bromide (0.5% μg mL-1)
Antibiotics: Laboratory grade of antibiotics with different mode(s) of action are shown in Table 3. Their supplements to the reaction mixture were provided as 2.5 μL, to replace the distilled water, of the antibiotic solution. The antibiotic concentration was calculated to correspond those given for BRR (Trimethoprim, Nalidixic acid, Amphotericin) and BR (Polymyxin B, Bacitracin, Chloramphenicol and Penicillin) selective media (Jansing and Rudolph, 1998; Elphinstone et al., 1996).
Tetrazolium and crystal violet: 2,3, 5 triphenyltetrazolium chloride in SMSA medium, selective for BR pathogen (Elphinstone et al., 1996) and crystal violet concentration in SMSA (Elphinstone et al., 1996) and Crystal Violet Pectate agar (CVP) selective for BL (Cupples and Kelman, 1974) were considered. The calculated concentrations are shown in Table 3.
Treatments numbering: List of treatments of the target organisms are shown in the following:
 |
| Antibiotics and dyes were added separately to each treatment, from 10-16, as shown in the figures |
| Table 3: | Antibiotic groups, mode(s) of action and concentration of dyes used |
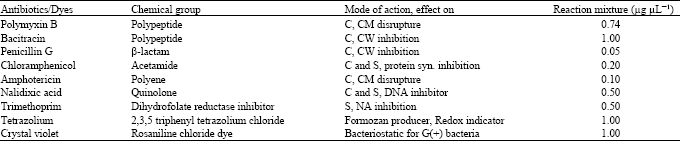 | |
| C: Cidal, S: Static, CM: Cell membrane, CW: Cell wall, PS: Protein synthesis NA: Nucleic acid | |
RESULTS
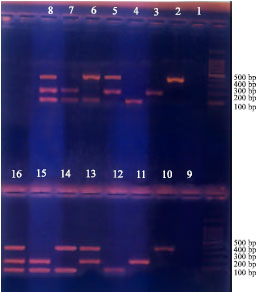
Multiplex-PCR: Figure 1 show the bands separated for the three pathogens, either single or in combination. R.solanacearum was developed at 400 bp (treatments 2, 5, 6 and 8). Clav. michiganensis ssp. sepedonicus was separated at 200 bp (Treatments 3, 5, 7 and 8). Er. carotovora ssp. atroseptica was shown up 100 bp (treatments 4, 6, 7 and 8).
Tuber extract: Cores from the stolon end of tubers were processed in phosphate buffer (pH. 7.0) as described by Pastrik (2000). Figure 2 shows no inhibition of multiplex-PCR results of Clav. michiganensis ssp. sepedonicus, Er.carotovora ssp. atroseptica and Ral. solanacearum. The development of bands at the base pairs mentioned above was noticed (control treatments from 2-8) and bands developed with potato extracts (treatments 10-16) are shown in the figure along with the negative controls (treatments 1 and 9).
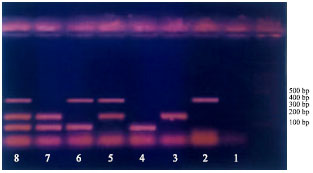 | |
| Fig. 1: | Treatments 2, 5, 6 and 8 showing bands of R. salanacearum at 400 bp. Treatments 3,5,7 and 8 showing bands of Clav sepedonicus at 200 bp. Treatments 4, 6, 7 and 8 showing atroseptica at 100 bp. Treatment 1 is negative controls |
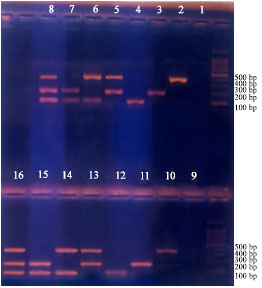 | |
| Fig. 2: | Potato extract did not affect band(s) development for different treatments (from 2-8 without extract) and treatments (10-16, with extract) |
Antibiotics: Polymyxin B (0.74 μg μL-1) and Bacitracin (1.00 μg μL-1), both are polypeptide antibiotics, caused complete inhibition of multiplex-PCR (Fig. 3, 4). Similar results for Trimethoprim (dihydroxyfolate reductase inhibitor) at the concentration used (0.5 μg μL-1) in selective media preparation (Fig. 5).
Other antibiotics as Penicillin G (β lactam) antibiotic at 0.05 μg μL-1, Choramphenicol (acetamide, at 0.2 μg μL-1), Amphotericin (polyene antibiotic at 0.1 μg μL-1) and Nalidixic acid (quinolone antibiotic at 0.5 μg μL-1) did not affect the resolution potential of the multiplex-PCR (Fig. 6).
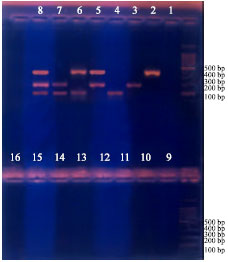 | |
| Fig. 3: | Treatments (2-8) without polymyxin B and treatments (10-16) with polymyxin B. Treatments 1 and 9, negative controls |
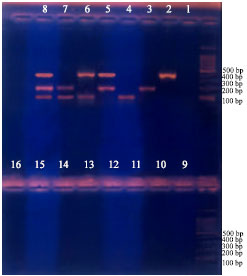 | |
| Fig. 4: | Complete inhibition with Bactracin for different treatments from 10-16. negative controls, 1 and 9 |
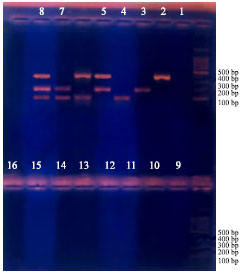 | |
| Fig. 5: | Complete inhibition with trimethoprim for different treatments (from 10-16). Negative controls, 1 and 9 |
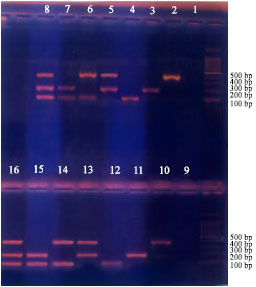 | |
| Fig. 6: | No inhibition with penicillin G (Treatments 10-16). Negative controls 1 and 9. Similar results for different treatments were reported for- Chloramphenicol-Amphotericin- Nalidixic |
Tetrazolium and crystal violet: The redox indicator (2,3,5 triphenyltetrazolium chloride at 1 μg μL-1) and the rosaniline dye crystal violet at 1 μg μL-1 did not inhibit the reaction (Fig. 7).
 | |
| Fig. 7: | No inhibition with 2,3,5 tri-phenyltetrazalium chloride for different treatments (from 10-16). Negative controls 1 and 9. Similar result was reported for different treatments with rosaniline dyes |
DISCUSSION
Multiplex -PCR is nowadays used in many diagnostic laboratories as it is cost saving, sensitive and reliable procedure (Elnifro et al., 2000). The technique, however, is subject to certain difficulties related to availability of primers for different plant pathogens (Martin et al., 2000; Schaad and Fredrick, 2002); the formation of dimers (Jannine et al., 1997) and the carryover of certain inhibitory factors in the samples (Rossen et al., 1992; Wilson, 1997).
The present work revealed that the high concentration of potato tuber extracts did not inhibit the multiplex reaction, to detect R. solanacearum, Clav. michiganensis ssp. sepedonicus and Er. carotovora spp. atroseptica the bacterial agents of BR, BRR and BL, respectively. In this regard Demeke and Adams (1992) studied the effect of plant polysaccharides and buffer additives on PCR results. They found that most polysaccharides tested were not inhibitory for PCR, with exception of two acidic polysaccharides, dextran sulfate and gum ghatti. They reported as well, that the addition of 0.5% Tween 20 reversed the inhibitory effect, similar to dextran sulphate, at lower concentration of Tween 20. This indicates that the composition of the reaction mixture and the carryover of certain compounds, to the pool of the reaction, may alter the reliability of PCR and consequently the multiplex -PCR. The inhibition of PCR by agar from bacteriological transport media was also reported by Gibb and Wong (1998).
Antibiotics and antimetabolites may be incorporated with laboratory media to increase selectivity for certain bacterial pathogens (Schaad et al., 2001). The antibiotics may exert either cidal or static effects on the viable cell, through one of four modes of action(s) that being targted to the inhibition of cell wall synthesis, interference with protein synthesis, disrupting the cell membrane and nucleic acid inhibition (Klement et al., 1990). Such mode(s) of action are functioning during the course of proliferation of different bacterial species, principally by interference with certain enzyme(s) activity. At this point a wonders may arise around the effect of such antibiotics on some enzymatic reactions in vitro, or extrinsic to the intact cell. Therefore, the possible chemical carryover, from the selective media, on the results of multiplex-PCR was considered in this study.
The polypeptide antibiotics, at rates used caused complete inhibition of multiplex-PCR results. Polymyxin B exerts a bactericidal effect through disrupting the cell membrane(s). Bacitracin has the same effect, but through inhibition of cell wall formation. Since the reaction mixture of multiplex-PCR is devoiding viable cells, so the recorded inhibition is independent of either the mode of action or function of these polypeptide antibiotics and may be attributed to the possible physical and chemical changes, during the process of thermocycling, that inhibited the polymerase reaction.
The dihydroxyfolate reductase, necessary for nucleic acid synthesis, is inhibited by the antibiotic Trimethoprim. This antibiotic exerts no cidal but static effect on the growing cells and caused complete inhibition of the multiplex-PCR reaction. Contrary to the aforementioned nucleic acid inhibitor, the quinolone antimetabolites as Nalidixic acid, which is a strong DNA inhibitor and exerts either cidal or static effects did not affect the multiplex-PCR. This indicates that the DNA inhibitors in vivo does not necessarily inhibits polymerization of DNA in vitro, in the multiplex -PCR reactions.
Moreover, the quinolones are a family of synthetic broad spectrum antibiotic. They prevent the unwinding of bacterial DNA through inhibiting the DNA gyrase (Hooper, 2001). This enzyme is essential during replication as it enables DNA supercoils to be relaxed and reformed.
Other groups of antibiotics as β-lactam (Penicillin G), acetamide (Chloramphenicol) and polyene (Amphotericin) regardless of their effect either cidal or static and /or their effects on specific vital process within the growing cell, showed no effect on results of muliplex-PCR in vitro. The negative effect may be attributed to the possible change in ionic strength, high temperature and /or the pH values on these antibiotic. Such attributes were considered by Loftin et al. (2008) in studies on degradation rates of a group of Tetracyclines and Methoprims. They reported that the hydrolysis rates of Tetracycline increased as pH and temperature increased following Arrhenius relationship. No degradation could be observed for Trimethoprim. Their results may partly explain the positive effect of Trimethoprim on PCR inhibition in this work. Further investigations are needed on the effect of other members of polypeptide antibiotics that may influence the multiplex-PCR as in the present study.
Tetrazolium chloride, which is a redox indicator in microbiological and cellular studies, is known to be heat and light unstable and may explain the negative effect on multiplex-PCR reaction. Rosaniline(s), as the basis of various dies and stains, including gentian violet, crystal violet and fuchsin, are widely used in selective media recipies and in staining (Blood et al., 2007). Crystal violet at the used concentration in the present study, did not inhibit multiplex-PCR reaction.
CONCLUSIONS
The present study showed that the tested polypeptide antibiotics (polymyxin B and Bacitracin) at the concentration(s) used in selective laboratory media resulted in a false negative reaction for detection of R.solanacearum, Clav.michiganensis ssp. sepedonicus and Er. carotovora sp. atroseptica. Trimethoprim (dihydroxyfolate reductase inhibitor), resulted in false negative results as well.
Penicillin G, Chloramphenicol, Amphotericin and Nalidixic acid did not interfer with the reaction. Redox indicator (TTC), rosaniline dye (CV) and tuber extract did not interfer with the reaction.
Further investigation are needed to study quantitatively the least concentration of antibiotics that interfer with PCR results.
The false negative multiplex-PCR reaction as a result of polypeptide antibiotic carryover may be important in other fields of studies as in forensic investigations.
REFERENCES
- Brownie, J., S. Shawcross, J. Theaker, D. Whitcombe, R. Ferrie, C. Newton and S. Little, 1997. The elimination of primer-dimer accumulation in PCR. Nucl. Acids Res., 25: 3235-3241.
Direct Link - Cupples, D. and A. Kelman, 1974. Evaluation of selective media for isolation of soft rot baceria from soil and plant tissue. Phytopathology, 64: 468-475.
Direct Link - De Lomas, J.G., F.J. Sunzeri and M.P Busch, 1992. False-negative results by polymerase chain reaction due to contamination by glove powder. Transfusion, 32: 83-85.
CrossRefPubMedDirect Link - Demeke, T. and R.P. Adams, 1992. The effects of plant polysaccharides and buffer additives on PCR. Biotechniques, 12: 332-334.
PubMed - Elnifro, E.M., A.M. Ashshi, R.J. Cooper and P.E. Klapper, 2000. Multiplex-PCR: Optimization and application in diagnostic virology. Clin. Microbiol. Rev., 13: 559-570.
CrossRefDirect Link - Elphinstone, J.G., J. Hennessy, J.K. Wilson and D.E. Stead, 1996. Sensitivity of different methods for the detection of Ralstonia solanacearum in potato tuber extracts. EPPO Bull., 26: 663-678.
CrossRefDirect Link - Gibb, A.P. and S. Wong, 1998. Inhibition of PCR by agar from bacteriological transport media. J. Clin. Microbiol., 36: 275-276.
Direct Link - Hooper, D.C., 2001. Emerging mechanisms of fluoroquinolone resistance. Emerg. Infect. Dis., 7: 337-341.
PubMedDirect Link - Jansing, H. and K. Rudolph, 1998. Physiological capabilities of Clavibacter michiganensis subsp. sepedonicus and development of a semi-selective medium. Zeitschrift Pflanzenkrankheiten Pflanzenschutz, 105: 590-601.
Direct Link - Kelman, A., 1954. The relationship of pathogenicity of Pseudomonas solanacearum to colony appearance in a tetrazolium medium. Phytopathology, 44: 693-695.
Direct Link - Loftin, K.A., C.D. Adams, M.T. Meyer and R. Surampalli, 2008. Effects of ionic strength, temperature and pH on degradation of selected antibiotics. J. Environ. Qual., 37: 378-386.
CrossRefPubMedDirect Link - Martin, R.R., D. James and C.A. Levesque, 2000. Impacts of molecular diagnostic technologies on plant disease management. Ann. Rev. Phytopathol., 38: 207-239.
CrossRefDirect Link - Pastrik, K.H., 2000. Detection of Clavibacter michiganensis Subsp. sepedonicus in potato tubers by multiplex PCR with coamplification of host DNA. Eur. J. Plant Pathol., 106: 155-165.
Direct Link - Rossen, L., P. Norskov, K. Holmstrom and O.F. Rasmussen, 1992. Inhibition of PCR by components of food samples, microbial diagnostic assays and DNA-extraction solutions. Int. J. Food Microbiol., 17: 37-45.
CrossRefPubMedDirect Link - Seal, S.E., L.A. Jackson, J.P.W. Young and M.J. Daniels, 1993. Differentiation of Pseudomonas solanacearum, Pseudomonas syzygii, Pseudomonas pickettii and the blood disease bacterium by partial 16S rRNA sequencing: Construction of oligonucleotide primers for sensitive detection by polymerase chain reaction. J. Gen. Microbiol., 139: 1587-1594.
CrossRefPubMedDirect Link - Tsai, Y.L. and B.H. Olson, 1992. Rapid method for separation of bacterial DNA from humic substances in sediments for polymerase chain reaction. Applied Environ. Microbiol., 58: 2292-2295.
Direct Link - Watson, R.J. and B. Blackwell, 2000. Purification and characterization of a common soil component which inhibits the polymerase chain reaction. Can. J. Microbiol., 46: 633-642.
PubMedDirect Link - Wilson, I.G., 1997. Inhibition and facilitation of nucleic acid amplification. Applied Environ. Microbiol., 63: 3741-3751.
PubMedDirect Link - EC, 1993. Council directive 93/85/EEC of 4 October 1993 on the control of potato ring rot. Official J. Eur. Commun., L259: 1-50.
Direct Link