
John E. Erpelding
USDA-Agricultural Research Service, Tropical Agriculture Research Station,
2200 Pedro Albizu Campos Ave., Suite 201, Mayaguez, Puerto Rico 00680-5470, USA
Ming Li Wang
USDA-Agricultural Research Service, Plant Genetic Resources Conservation Unit,
1109 Experiment Street, Griffin, Georgia 30223, USA
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 2 | Page No.: 127-133







ABSTRACT
A subset of 97 accessions was randomly selected from the sorghum collection maintained by the US National Plant Germplasm System and evaluated for anthracnose disease response in Isabela, Puerto Rico during the 2004 wet growing season. Two evaluations were conducted and climatic conditions varied between evaluations. The amount of rainfall was similar between evaluations; however, the lower temperatures during the second evaluation appeared more favorable for disease expression. Thirty accessions conferred a resistant disease response for the two evaluations. Sorghum germplasm from 28 countries were evaluated for anthracnose disease response and resistance accessions were observed for collections from 14 countries. Results indicate that germplasm from Ethiopia and Mali could be an important source of anthracnose resistance. The germplasm collections from Benin, Burundi, Liberia, Malawi, Nigeria, Sudan, South Africa and Uganda could also be useful for the identification of anthracnose resistance and could contribute to the genetic diversity for disease resistance to enhance sorghum improvement.
PDF Abstract XML References
How to cite this article
John E. Erpelding and Ming Li Wang, 2007. Response to Anthracnose Infection for a Random Selection of Sorghum Germplasm. Plant Pathology Journal, 6: 127-133.
DOI: 10.3923/ppj.2007.127.133
URL: https://scialert.net/abstract/?doi=ppj.2007.127.133
DOI: 10.3923/ppj.2007.127.133
URL: https://scialert.net/abstract/?doi=ppj.2007.127.133
INTRODUCTION
Sorghum anthracnose (Colletotrichum sublineolum P. Henn, Kabát and Bubák) occurs worldwide and is considered one of the most important diseases limiting sorghum production (Cardwell et al., 1989; Thakur and Mathur, 2000; Ngugi et al., 2002; Marley, 2004). The disease is frequently observed in tropical and subtropical environments where hot and humid climatic conditions contribute to the development and spread of the disease. Grain yield losses of more than 50% have been reported (Harris et al., 1964; Thakur and Mathur, 2000). Experimental evaluations have shown grain yield losses ranging from 30 to 70% for susceptible cultivars (Ali et al., 1987; Thomas et al., 1996). Yield losses are typically associated with a reduction in seed weight due to poor grain development (Ali et al., 1987). Leaf death prior to maturity and stalk breakage are frequently observed for highly susceptible cultivars resulting in significant reductions in grain yield.
The pathogen is capable of infecting all above ground tissues of the sorghum plant including the leaf, stalk, panicle and seed (Coleman and Stokes, 1954; Harris et al., 1964; Thakur and Mathur, 2000; Hess et al., 2002). Foliar infection is more frequently observed and can occur at every stage of plant development (Thakur and Mathur, 2000). Symptoms of anthracnose infection generally appear 40 days after seedling emergence on susceptible cultivars. Typical symptoms of foliar infection include circular, elliptical or elongated lesions with tan, red, purple or black margins depending on the response of the host plant. On susceptible cultivars, the lesions will elongate and coalesce covering most of the leaf surface resulting in leaf senescence. Black spots or fruiting bodies (acervuli) will develop in the center of the lesions as the fungus sporulates.
The disease can be controlled through the use of resistant cultivars, but the development of resistant cultivars is hindered by the occurrence of multiple pathotypes within the pathogen population (Ali and Warren, 1987; Cardwell et al., 1989; Pande et al., 1991; Valerio et al., 2005). This pathotype variation and the occurrence of new virulent pathotypes have contributed to a loss of host plant resistance (Rosewich et al., 1998). To manage the disease more effectively, additional sources of resistance will be essential. The evaluation of sorghum accessions from germplasm collections to identify additional sources of anthracnose resistance has been successfully accomplished for evaluation subsets from Sudan, Mali and Mozambique (Erpelding and Prom, 2004; Erpelding et al., 2005; Erpelding and Prom, 2006). But with more than 43,000 sorghum accessions in the US National Plant Germplasm System (USDA-ARS National Genetic Resources Program, 2006), additional information is needed to aid in the identification of regions associated with anthracnose resistance. Therefore, a random selection of sorghum germplasm representing 28 countries was evaluated for resistance to anthracnose to determine if resistant germplasm was more frequently observed for specific countries of origin in order to identify additional collections for evaluation.
MATERIALS AND METHODS
A subset of 97 accessions was randomly selected from the sorghum collection maintained by the US National Plant Germplasm System in Griffin, Georgia (Table 1). Only sorghum accessions with complete characterization information were included in the subset representing 28 countries. Molecular marker data was also available for 93 of the 97 sorghum accessions included in the evaluation. Seed samples for the germplasm evaluation subset were obtained from the USDA-ARS Plant Genetic Resources Conservation Unit in Griffin, Georgia. The anthracnose disease evaluation was conducted at the USDA-ARS Tropical Agriculture Research Station in Isabela, Puerto Rico during the wet growing season in 2004. The first evaluation was planted 2 August 2004 with the second evaluation planted 21 October 2004. Weather data for the two disease evaluations is presented in Table 2. Each sorghum accession was planted in a single 1.8 m length row with 0.9 m row spacing using a partially balanced lattice design and three replications. Border rows of anthracnose susceptible genotypes were planted around each experiment. A single application of fertilizer at a rate 560 kg ha-1 (15-5-10 NPK) was conducted at planting. Lorsban 15G (O,O-diethyl O-(3,5,6-trichloro-2-pyridinyl) phosphorothioate) granular insecticide (Dow Agro Sciences, Indianapolis, IN) was applied at a rate of 8 kg ha-1 during planting to prevent seed loss from fire ants. For the first evaluation, three applications of supplemental irrigation were conducted for stand establishment prior to anthracnose inoculation with a single application of supplemental irrigation following inoculation. Supplemental irrigation was applied once for the second evaluation seven days after planting. Mechanical tillage and hand hoeing were used for weed control.
| Table 1: | Country of origin, disease rating and average percentage of infected leaf area for the 97 sorghum accessions and 11 control samples included in the two anthracnose disease evaluations conducted in Isabela, Puerto Rico during the wet growing season of 20041 |
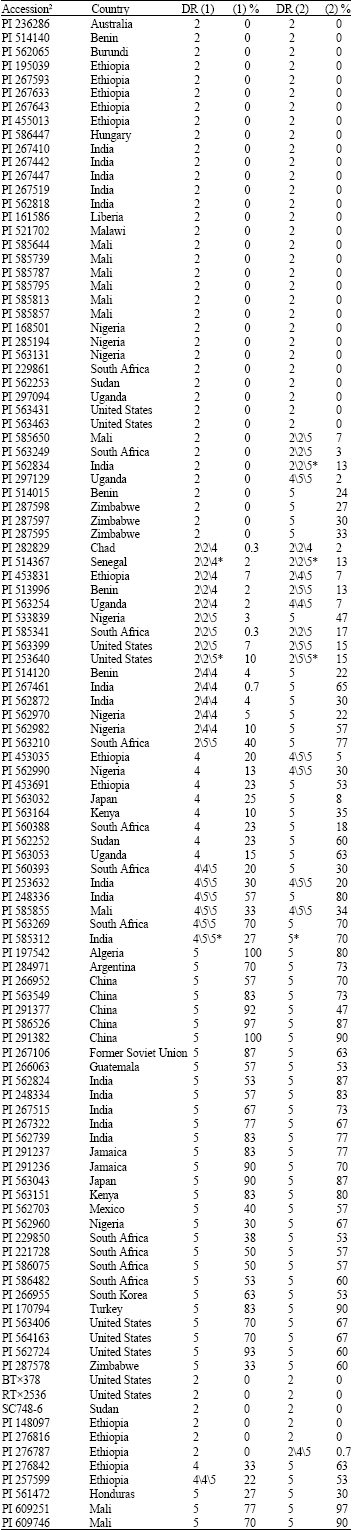 | |
| 1DR (1) = Disease rating for the first evaluation planted 2 August 2004. DR (2) = Disease rating for the second evaluation planted 21 October 2004. The disease rating is based on a 1-5 scale (Erpelding and Prom, 2006). Data is presented for the three replications when variation was observed between replications. * = Indicates within accession variation was observed for the germplasm line with the most predominant disease phenotype reported. % (1) = Percentage of infected leaf area for the first evaluation with a LSD(0.05) = 19.4. % (2) = Percentage of infected leaf area for the second evaluation with a LSD(0.05) = 20.7. The percentage of infected leaf area is an average of the three replications for each evaluation, 2Plant introduction number and country of origin for the sorghum accessions include in the evaluation (USDA-ARS National Genetic Resources Program, 2006). Accessions are arranged by anthracnose infection response from resistant to susceptible. Control samples = BTx378, RTx2536, SC748-6, PI 148097, PI 276816, PI 276787, PI 276842, PI 257599, PI 561472, PI 609251 and PI 609746 | |
Eleven control samples were included in the anthracnose evaluation to determine variation in infection response within and between experiments. Two accessions from Mali, West Africa (PI 609251 and PI 609746) were included as anthracnose susceptible controls.
| Table 2: | Total rainfall, number of days with rain, temperature and relative humidity for the two anthracnose evaluations conducted in Isabela, Puerto Rico during the 2004 wet growing season |
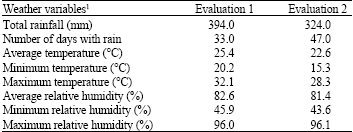 | |
| 1Weather data was summarized after disease inoculation until the final anthracnose reading was conducted for the evaluations. Anthracnose disease evaluations for the 97 sorghum accessions and 11 control samples were conducted from 3 October 2004 to 30 October 2004 for the first evaluation and from 27 December 2004 to 9 February 2005 for the second evaluation | |
Three resistant (PI 148097, PI 276787 and PI 276816) and two susceptible (PI 257599 and PI 276842) accessions from Ethiopia were included as controls. The anthracnose susceptible control PI 561472 (Sureño) is an inbred variety developed in Honduras (Meckenstock et al., 1993). BTx378, RTx2536 and SC748-6 are breeding lines included in the evaluation as anthracnose resistant controls. BTx378, RTx2536 and SC748-6 are also resistant to anthracnose pathotypes present in the continental United States with SC748-6 conferring resistant to numerous pathotypes found in the United States and worldwide.
Preparation of anthracnose cultures, inoculation of sorghum plants and evaluation of infection response were as described by Erpelding and Prom (2006). Briefly, anthracnose cultures were established from infected leaf tissue and these cultures were used to colonize sorghum seed for inoculation. Plants were inoculated approximately 30 days after planting during the 7-10 leaf stage by placing anthracnose colonized seed in the leaf whorl. The plant host response to anthracnose infection was evaluated 13, 34, 45 and 61 days after inoculation for the first evaluation and 34, 47 and 77 days after inoculation for the second evaluation using a 1-5 rating scale. The percent infected leaf area for the susceptible accessions was estimated during the final evaluation with the exception of the highly susceptible accessions where the disease resulted in rapid leaf senescence. For these accessions, infected leaf area was estimated during the evaluation conducted approximately 45 days after inoculation prior to plant death. The disease response from the final rating and the percent infected leaf area were used for statistical analysis. Nonparamentric statistical analysis of infected leaf area means was used to conduct comparisons for evaluations with unequal samples sizes using a t-test (p = 0.05).
RESULTS
The variation in anthracnose infection response (Table 1) was highly significant (p = 0.01) for the 97 sorghum accessions (data not shown). For the first evaluation, 38 accessions conferred a resistant response. A susceptible response was observed for 44 accessions with 30 accessions rated as highly susceptible. The infection response varied between replications for 15 accessions during the first evaluation and these accessions were considered susceptible to anthracnose. The disease response was evaluated 13 days after inoculation for the first evaluation and 11 of 44 susceptible accessions showed development of anthracnose lesions on inoculated leaves across the three replications (data not shown). Thirty-three of the anthracnose susceptible accessions showed a susceptible disease response 34 days after inoculation and a susceptible response was observed for 43 accessions within 45 days after inoculation. For the second evaluation, 30 accessions were rated as resistant to anthracnose. A susceptible response was observed for 57 accessions and all susceptible accessions in the second evaluation were rated as highly susceptible. A susceptible response was observed for 45 of the 57 accessions rated as susceptible 34 days after inoculation with a consistent susceptible response observed for 54 accessions 47 days after inoculation (data not shown). The disease response varied between replications for 10 accessions during the second evaluation and these accessions were considered susceptible. Overall, 30 of the 97 accessions evaluated showed a resistant response to anthracnose infection over the two evaluation periods with 67 accessions susceptible to the disease. No variation in disease response within and between experiments was observed for 60 accessions. Seven accessions that varied from a resistant to susceptible response between replications for the first evaluation also showed variation for infection response in the second evaluation. The 59 accessions considered susceptible to anthracnose in the first evaluation were also rated as susceptible in the second evaluation. Eight accessions rated as resistant during the first evaluation showed a susceptible response for the second evaluation. The response to anthracnose infection was similar between experiments for the 11 control samples included in the evaluation with the exception of PI 276787, which show a variable response in the second evaluation (Table 1). Anthracnose infection was severe for both evaluations resulting in leaf senescence 45 days after inoculation for the highly susceptible accessions. Plant death prior to maturity was observed for several accessions resulting in poor seed development and lodging.
| Table 3: | Country of origin, number of resistant and susceptible sorghum accessions and the average percentage of infected leaf area for the 97 sorghum accession included in the anthracnose evaluation conducted in Isabela, Puerto Rico during 2004 |
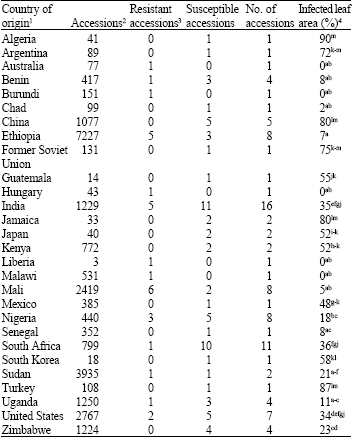 | |
| 1Sorghum germplasm accessions from 28 countries were included in the anthracnose disease evaluation (USDA-ARS National Genetic Resources Program, 2006), 2Total number of sorghum accessions in the US National Plant Germplasm System for each of the 28 countries included in the subset of 97 accessions evaluated for anthracnose disease response (USDA-ARS National Genetic Resources Program, 2006), 3Thirty of the 97 accessions evaluation for anthracnose disease response showed a consistent resistant response for the two evaluations with 67 accessions rated as susceptible, 4Infected leaf area for the sorghum accessions was averaged over the two evaluations based on country of origin. Infected leaf area percentages followed by the same letter are not significantly different (p = 0.05) | |
Sorghum germplasm accessions from 28 countries were included in the anthracnose evaluation and the number of resistant and susceptible accessions varied based on the country of origin (Table 3).
The percentage of infected leaf area was determined as a measure of the quantitative response to anthracnose infection (Table 1). The severity of anthracnose infection based on the percentage of infected leaf area was similar between the two experiments with an average of 44% infected leaf area for the 59 susceptible accessions in the first evaluation and 47% for the 67 susceptible accessions identified in the second evaluation. The variation between experiments for the percentage of infected leaf area for the susceptible accessions was not significant (data not shown). The percentage of infected leaf area was similar for 31 of the 59 accessions that showed a susceptible response for both experiments. For 18 accessions, the percentage of infected leaf area was greater during the second evaluation; whereas, 10 accessions showed a higher percentage of leaf infection during the first evaluation. The severity of infection was generally low for the sorghum accessions that showed variation for infection response within and between experiments. In contrast, infection severity was high for the 30 highly susceptible accessions with an average of 70% infected leaf area for the two evaluations. The six anthracnose susceptible controls included in the experiment were rated as highly susceptible for the two evaluations with a greater percentage of infected leaf area observed during the second evaluation. The anthracnose infected leaf area for the susceptible controls average 46% for the first evaluation compared to an average of 66% for the second evaluation, but the difference in infected leaf area between experiments was not significant (data not shown). The percentage of infected leaf area was averaged over accessions based on country of origin and variation in anthracnose infection severity was observed for the sorghum germplasm associated with country of origin (Table 3).
DISCUSSION
A greater number of accessions were susceptible to anthracnose infection in the second evaluation suggesting climatic conditions were more favorable for disease development. Environmental conditions can significantly influence the development of anthracnose disease symptoms on the host plant (Pande et al., 1994; Thakur and Mathur, 2000; Hess et al., 2002). Pande et al. (1994) indicated that warm temperatures, high relative humidity, excessive rainfall and prolonged dew periods were favorable climatic conditions that would enhance disease development. Anthracnose evaluations conducted at several locations in Burkina Faso and Mali, West Africa indicated that disease severity was greater at the wetter locations (Neya and Le Normand, 1998; Hess et al., 2002). Variation in disease expression was also observed by Erpelding and Prom (2004) for an evaluation conducted during the wet and dry growing seasons in Isabela, Puerto Rico with greater disease severity observed during the wet season. Although less rainfall was received during the second evaluation (Table 2), the frequency was greater and may have contributed to the increase in the number of susceptible accessions and the higher disease severity observed for the evaluation. In addition, approximately 75% of the rainfall occurred prior to the disease evaluation conducted 34 days after inoculation for the first evaluation. The excessive rainfall and the irrigation of the first evaluation after inoculation did not appear to enhance disease expression as compared to the infection response observed for the second evaluation even though rainfall occurred on a similar number of days after anthracnose inoculation for each experiment prior to the disease evaluation conducted 34 days after inoculation. Disease expression for the second evaluation may have also been enhanced by lower temperature that could have prolonged the dew period. Additionally, the higher temperatures during the first evaluation could have hindered disease expression since maximum temperatures exceeded 30°C. Pande et al. (1994) considered temperatures above 30°C to be detrimental to anthracnose development. Although climatic conditions appeared to be more favorable for disease expression during the second evaluation, the disease response for over 60% of the accessions included in the evaluation was similar between experiments suggesting that variable climatic conditions during the wet growing season in Isabela, Puerto Rico are generally favorable for anthracnose disease development.
Approximately 30% of the sorghum accessions included in the anthracnose evaluation showed a resistant disease response for the two experiments. These resistant accessions were obtained from 14 countries. Phenotypic and molecular genetic data indicate genetic variation for the accessions, which may suggest possible genetic variation for anthracnose resistance. Limited information is available on the genetics of host plant resistance for anthracnose. A single dominant gene conditioning anthracnose resistance was identified from a sorghum germplasm accession from Sudan (LeBeau and Coleman, 1950) whereas, a single recessive gene for anthracnose resistance was identified from a germplasm line from Ethiopia (Boora et al., 1998). This would suggest possible genetic variation for host plant resistant between regions. In addition, the pathogen is considered highly variable within and between regions (Ali and Warren, 1987; Cardwell et al., 1989; Pande et al., 1991; Guthrie et al., 1992; Valerio et al., 2005), which could contribute to the evolution of host plant resistance. Thus, the anthracnose resistant germplasm identified from this evaluation may provide additional sources of resistance for sorghum improvement.
Molecular genetic evaluation of the sorghum accessions included in this study indicated that accessions cluster by country of origin (Wang et al., 2006). The response to anthracnose infection also showed an association with country of origin. Although the number of accessions from individual countries is low, the results would suggest that anthracnose resistant sorghum germplasm may occur at a higher frequency for certain countries of origin. Mali and Ethiopia appear to be regions associated with anthracnose resistant sorghum germplasm. Erpelding and Prom (2004) evaluated a collection of sorghum germplasm from Mali, West Africa and showed that anthracnose resistant germplasm occurred at a high frequency in the collection. Presently, no information is available on the genetic diversity of host plant resistance for the sorghum germplasm from these regions to suggest that the higher frequency of resistant germplasm is associated with greater genetic diversity for anthracnose resistance. Therefore, evaluation of sorghum germplasm collections from other regions would be desirable for the identification of anthracnose resistant germplasm to enhance the genetic diversity of host plant resistance available for sorghum improvement. The results of this study would indicate that sorghum collections from other African nations such as Burundi, Liberia and Malawi could also be a source of anthracnose resistant germplasm. Although the frequency of anthracnose resistant germplasm appears to be lower for the sorghum collections from Benin, Nigeria, Sudan, South Africa and Uganda, evaluation of the sorghum collections from these regions should result in the identification of anthracnose resistant germplasm. The anthracnose resistant control sample, SC748-6, used in this study, was developed from a sorghum accession from Sudan. In addition, data presented by Erpelding et al. (2005) would suggest that Sudanese sorghum germplasm could be a source of genetic diversity for anthracnose resistance. Although no anthracnose resistant accessions were identified for the sorghum germplasm from Chad, Senegal and Zimbabwe, the average percentage of infected leaf area for the accessions evaluated was low suggesting resistant germplasm could be present in the germplasm collections from these regions. In contrast, the germplasm collections from Algeria, Argentina, China, Jamaica and Turkey may lack sources of anthracnose resistance, since the average percentage of infected leaf area for the accessions evaluated was high. Only a limited number of sorghum accessions were evaluated from these regions and it is possible that anthracnose resistance germplasm may be present in these collections at a low frequency. For 14 countries, only one accession was evaluated in this study and therefore additional disease characterization would be essential to verify specific regions associated with anthracnose resistance.
Large scale evaluations would be useful to enhance disease characterization of the sorghum germplasm collection maintained by the US National Plant Germplasm System. As suggested by Erpelding et al. (2005), large scale evaluations can successfully result in the identification of anthracnose resistant germplasm. The germplasm collections from eight countries included in this study have more than 1,000 accessions (Table 3) and large scale evaluations would be necessary to determine disease response for the accessions from these collections. The anthracnose resistant germplasm identified from these evaluations can then be more extensively evaluated to identify germplasm for sorghum improvement and genetic characterization. Climatic conditions are highly favorable during the wet growing season in Isabela, Puerto Rico for anthracnose disease development. In general, the wet growing season occurs from May to December. The cooler temperatures during the winter months of the wet season appear to enhance expression of anthracnose disease symptoms and large scale evaluations during this period could be successfully conducted to identify resistant germplasm and reduce potential environmental interactions between the pathogen and plant host.
Disclaimer: Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendations or endorsement by the U.S. Department of Agriculture.
REFERENCES
- Ali, M.E.K. and H.L. Warren, 1987. Physiological races of Colletotrichum graminicola on sorghum. Plant Dis., 71: 402-404.
Direct Link - Ali, M.E.K., H.L. Warren and R.X. Latin, 1987. Relationship between anthracnose leaf blight and losses in grain yield of sorghum. Plant Dis., 71: 803-806.
CrossRefDirect Link - Boora, K.S., R. Frederiksen and C. Magill, 1998. DNA-based markers for a recessive gene conferring anthracnose resistance in sorghum. Crop Sci., 38: 1708-1709.
Direct Link - Erpelding, J.E. and L.K. Prom, 2004. Evaluation of malian sorghum germplasm for resistance against anthracnose. Plant Pathol. J., 3: 65-71.
CrossRefDirect Link - Erpelding, J.E., L.K. Prom and W.L. Rooney, 2005. Variation in anthracnose resistance within the Sudanese sorghum germplasm collection. Plant Genet. Resour. Newslett., 141: 11-14.
Direct Link - Erpelding, J.E. and L.K. Prom, 2006. Variation for anthracnose resistance within the sorghum germplasm collection from Mozambique, Africa. Plant Pathol. J., 5: 28-34.
CrossRefDirect Link - Guthrie, P.A.I., C.W. Magill, R.A. Frederiksen and G.N. Odvody, 1992. Random amplified polymorphic DNA markers: A system for identifying and differentiating isolates of C. graminicola. Phytopathology, 82: 832-835.
Direct Link - Harris, H.B., B.J. Johnson, J.W. Dobson Jr. and E.S. Luttrell, 1964. Evaluation of anthracnose on grain sorghum. Crop Sci., 4: 460-462.
Direct Link - Hess, D.E., R. Bandyopadhyay and I. Sissoko, 2002. Pattern analysis of sorghum genotype H environment interaction for leaf, panicle and grain anthracnose in Mali. Plant Dis., 86: 1374-1382.
CrossRefDirect Link - Meckenstock, D.H., F. Gomez, D.T. Rosenow and V. Guiragossian, 1993. Registration of sureno sorghum. Crop Sci., 33: 213-213.
Direct Link - Marley, P.S., 2004. Effects of integrating host resistance with time of planting or fungicides on anthracnose and grain mould and yield of sorghum (Sorghum bicolor) in the Nigerian Northern Guinea Savanna. J. Agric. Sci., 142: 345-350.
Direct Link - Neya, A. and M. le Normand, 1998. Responses of sorghum genotypes to leaf anthracnose (Colletotrichum graminicola) under field conditions in Burkina Faso. Crop Prot., 17: 47-53.
CrossRefDirect Link - Ngugi, H.K., S.B. King, G.O. Abayo and Y.V.R. Reddy, 2002. Prevalence, incidence and severity of sorghum diseases in Western Kenya. Plant Dis., 86: 65-70.
CrossRefDirect Link - Pande, S., L.K. Mughogho, R. Bandyopadhyay and R.I. Karunakar, 1991. Variation in pathogenicity and cultural characteristics of sorghum isolates of Colletotrichum graminicola in India. Plant Dis., 75: 778-783.
Direct Link - Pande, S., R.P. Thakur, R.I. Karunakar, R. Bandyopadhyay and B.V.S. Reddy, 1994. Development of screening methods and identification of stable resistance to anthracnose in sorghum. Field Crops Res., 38: 157-166.
CrossRefDirect Link - Rosewich, U.L., R.E. Pettway, B.A. McDonald, R.R. Duncan and R.A. Frederiksen, 1998. Genetic structure and temporal dynamics of a Colletotrichum graminicola population in a sorghum disease nursery. Phytopathology, 88: 1087-1093.
CrossRefDirect Link - Thomas, M.D., I. Sissoko and M. Sacko, 1996. Development of leaf anthracnose and its effect on yield and grain weight of sorghum in West Africa. Plant Dis., 80: 151-153.
Direct Link - Valerio, H.M., M.A. Resende, R.C.B. Weikert-Oliveira and C.R. Casela, 2005. Virulence and molecular diversity in Collectotrichum graminicola from Brazil. Mycopathologia, 159: 449-459.
CrossRefDirect Link - Wang, M.L., R. Dean, J.E. Erpelding and G.A. Pederson, 2006. Molecular genetic evaluation of sorghum germplasm differing in response to fungal diseases: Rust (Puccinia purpurea) and anthracnose (Collectotrichum graminicola). Euphytica, 148: 319-330.
Direct Link