
G. Moravvej
Faculty of Agriculture, Ferdowsi University of Mashad, Mashad, Iran
S. Abbar
Faculty of Agriculture, Ferdowsi University of Mashad, Mashad, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 1 | Page No.: 48-54







ABSTRACT
In the present study, the effects of volatile components of Citrus paradisi, C. aurantium, C. limonium and C. sinensis peel essential oils were investigated on the cowpea adult bruchid, Callosobruchus maculatus (F.). The oils were extracted from the fruit peels using hydrodistillation. The results indicated that the citrus oils had high fumigant activity against adult beetles. The mortality of 1-2 day-old adults increased with concentration and exposure time from 3 to 24 h. The oil of C. paradisi was more effective than those of C. aurantium and C. limonium (The LC50 values were 125, 145 and 235 &Amp;plusmn;1 L-1 at 24 h exposure, respectively). The oil of C. sinensis proved to be least toxic (LC50 = 269 &Amp;plusmn;1 L-1). The results suggested that citrus peel oils can be used as potential control measure against cowpea beetles.
PDF Abstract XML References Citation
How to cite this article
G. Moravvej and S. Abbar, 2008. Fumigant Toxicity of Citrus Oils Against Cowpea Seed Beetle Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). Pakistan Journal of Biological Sciences, 11: 48-54.
DOI: 10.3923/pjbs.2008.48.54
URL: https://scialert.net/abstract/?doi=pjbs.2008.48.54
DOI: 10.3923/pjbs.2008.48.54
URL: https://scialert.net/abstract/?doi=pjbs.2008.48.54
INTRODUCTION
The cowpea seed beetle, Callosobruchus maculatus (F.) is a field-to-store pest of pulses and the level of field infestation is a major factor that influences the bionomics of this bruchid in storage (Olubayo and Port, 1997; Nahdy et al., 1998). According to Rees (2004), loss of seed material is considerable-each adult Callosobruchus emerging from a cowpea (Vigna unguiculata) would have consumed about 25% of the seed from which it emerged. Damaged seed often does not germinate or germinate well. Heavy infestations of bruchids can cause heating of commodity which results in quality loss and mould growth. Currently, only two fumigants, methyl bromide and phosphine, are widely used against stored product insect pests (Perry et al., 1998). The use of methyl bromide is being restricted because of its potential to damage the ozone layer (Butler and Rodriguez, 1996; MBTOC, 1998). The future use of phosphine could be threatened by the further development of resistant strains of pests (Bell and Wilson, 1995; Daglish and Collins, 1999). Therefore, there is an urgent need to develop safe alternatives to conventional insecticides and fumigants to protect stored-products from insect infestations.
In the search for alternatives to conventional fumigants, essential oils extracted from aromatic plants have been widely investigated and their toxicity to stored-product insects has been of special interest during the last decade (Shaaya et al., 1993; Ho et al., 1997; Obeng-Ofori and Reichmuth, 1997; Obeng-Ofori et al., 1997; Huang et al., 2002). The fruit peels of some Citrus species have been reported to have insecticidal properties against insect pests (Don-Pedro, 1985; Onu and Sulyman, 1997; Elhag, 2000). The major active component of citrus oils is limonene. Insecticidal activity of limonene has been successfully applied for the control of insect parasitoids of pet animals. (+)-limonene was toxic to malation-resistant fleas (Collart and Hink, 1986) and to all life stages of the cat flea, Ctenocephalides felis (Hink and Fee, 1986). Weekly application of (+)-limonene reduced flea and tick infestation by 80% in dog and cat, with no adverse effects on blood composition or liver or kidney function (Tonelli, 1987). Limonene-treated water had adverse effect on oviposition of female mosquitoes (Kassir et al., 1989).
The toxicity of powdered sun-dried orange and grapefruit peels to C. maculatus has been demonstrated (Don-Pedro, 1985). In another studies, the essential oils of citrus peels proved to reduce oviposition or larval emergence through parental adult mortality (Don-Pedro, 1996; Elhag, 2000). Studying on the toxicity of essential oils from various plant species, Papachristos and Stamopoulos (2002) demonstrated the fumigant activity of Citrus sinensis for males and females of the bruchid, Acanthoscelides obtectus (Say).
The above studies were limited only to one or two Citrus species and conducted mainly using powdered fruit peels. In the present study, the activities of volatile fractions of essential oils extracted from the fruit peels of four different Citrus species on the cowpea adult bruchid, C. maculatus were investigated.
MATERIALS AND METHODS
Rearing of test insects: The study was undertaken in Entomology laboratory of Dept. of Plant Protection, Ferdowsi University of Mashad, Iran. A culture of non-flight form of the cowpea weevil, C. maculatus was established on the seeds of unshelled brown cowpea, V. unguiculata in wide-mouthed plastic jars under laboratory conditions (32±6°C, 45±5% RH and 12-12 h light-dark cycle). Adding new seeds and removing very old ones to and from the main culture monthly prevented crowding of the bruchids and so no flight form was developed. One to two days old adults were used for all bioassays.
Extraction of essential oil: The citrus fruits were collected from the central local market of Mashad, Iran during November and December 2006. The essential oils were extracted from fresh rind tissue (albedo and flavedo) of fruits by water steam distillation using a Clevenger apparatus. The Citrus species were C. sinensis Osbeck., C. aurantium Risso., C. limonium Risso. and C. paradisi Macf. (Rutaceae). About 1.5 mL oil was extracted per 100 g fresh peel. Extracted oils were stored at 5°C until the onset of bioassays.
Bioassays: The bioassays were conducted following the preparation of essential oils as described by Rahman and Schmidt (1999) and Keita et al. (2001) with slight modifications. The 2 cm diameter pieces of filter paper (Whatman No. 1) were impregnated with six different oil concentrations (including 0 μl L-1 as control) to give the equivalent concentrations in air of 148-555 μl L-1 for C. limonium and C. sinensis, 74-370 μl L-1 for C. aurantium and 74-444 μl L-1 for C. paradisi. The range of concentrations had been chosen on the basis of a number of preliminary trials. The filter paper was attached to the undersurface of the screw cap of a glass vial (volume 27 mL). The cap was screwed tightly onto the vial containing 5 pairs of one-to 2 day-old C. maculatus adults. Six replicates of each control and treatment were set up. Mortality was recorded after 3, 6, 9, 12 and 24 h from the commencement of exposure. When no leg or antennal movements was observed, insects were considered dead.
Data analysis: Mortality data of adults at 24 h exposure time for each plant species were analysed with the probit model (Finney, 1971) using a Maximum Likelihood Program (POLO-PC, LeOra Software, Berkeley, California). The program POLO-PC calculates a theoretical natural response for each experiment, based on the pattern of mortality at all concentration levels, including controls. The results include estimate of the LC50 (and other LCs if required) and the 95% confidence limits, slope and intercept of probit mortality regression and the relevant statistical tests (such as t-ratio, g factor and heterogeneity). For comparison of the probit mortality lines of treatments, the program also provides the likelihood ratio tests of equality and parallelism. The data obtained from repeated experiments were pooled for probit analysis, after acceptance of the hypothesis of equality following likelihood ratio tests (Russel et al., 1977). Estimated median lethal concentration to kill 50% of aphids was expressed as LC50 (μL oil per unit air liter). The resistance ratio and 95% confidence limits of this ratio were determined between adults from different treatments and comparisons were made based on the procedure described by Robertson and Preisler (1992). The estimates of parameters needed for computing confidence limits of the resistance ratio were provided by individual probit analysis in the POLO-PC output.
RESULTS
The essential oil vapors of the peel of four Citrus species showed variable toxicity to adults of C. maculatus, depending on concentration and exposure period (Table 1). There was no mortality in the presence of 148 μl L-1 of C. sinensis oil at 3 and 6 h exposure. At 3 h exposure, the maximum rates of mortality, 24, 29 and 40%, were achieved in the presence of 400, 370 and 444 μl L-1 of C. limonium, C. aurantium and C. paradisi oils, respectively. At 12 h exposure, the minimum rates of mortality of 5 and 25% in adults were attained by the presence of 148 μl L-1 of C. sinensis and C. limonium oils, respectively. The essential oils of C. paradisi and C. aurantium achieved the mortalities lower than 16% in the presence of 74 μl L-1 for 12 h exposure. The essential oils of C. sinensis and C. limonium achieved respectively the mortalities of 96 and 91% against the adults, at the highest concentration and exposure period investigated (556 μl L-1 air, 24 h). The essential oils of C. paradisi and C. aurantium were more active and achieved, respectively 96 and 84% mortalities after 24 h exposure at much lower concentrations (444 and 370 μl L-1 air, respectively). In all bioassays, the control mortality did not exceed 1.7% at 3 h and 3.3% at 24 h exposure. These control mortalities were quite below the recommendation (30%) made by the FAO (Anonymous, 1970).
Mortality data were adjusted for natural mortality in controls according to Abbott (1925) formula and within each oil concentration, were plotted against exposure periods. The results showed that there were positive and linear significant relationships between percent mortality of adults and duration of exposure to the
| Table 1: | Mean percent (±SE) mortality in the adults of C. maculatus exposed for various periods to four citrus essential oils at different concentrations (n = 6)A |
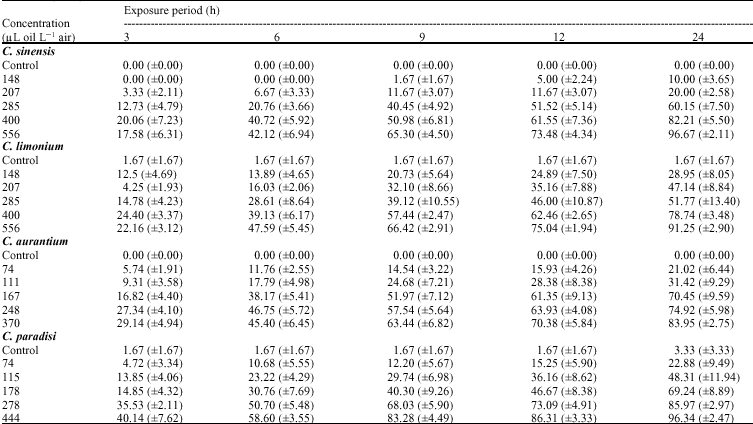 | |
A: Oils were applied to 2 cm filter papers held in 27 mL vials | |
| Table 2: | Analysis results of linear regressions of C. maculatus mortality data on exposure periods A in various concentrations of four citrus essential oilsB |
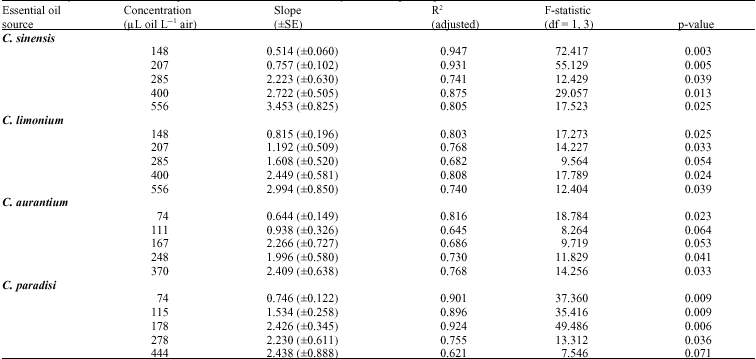 | |
A: Mortality data at each exposure period was a mean of six replicates, B: Oil applied to 2 cm filter papers held in 27 mL vials. Exposure periods were 3, 6, 9, 12 and 24 h | |
essential oil vapors within all concentration levels and plant species (p<0.05), although not significantly within concentrations of 285 μl L-1 of C. limonium (p = 0.054); 111 and 167 μL L-1 of C. aurantium (p = 0.064 and 0.053, respectively) and 444 μl L-1 of C. paradisi (p = 0.071). Coefficients of determination (R2) indicated that between 62-95% of the variation in the rates of adult mortality was explained by duration of exposure to essential oils. Within each essential oil, the slopes of regressions of mortality rates on exposure times were smaller in low concentrations than those in high concentrations (Table 2).
| Table 3: | Fumigant toxicity of four citrus essential oil vapors to C. maculatus adults A |
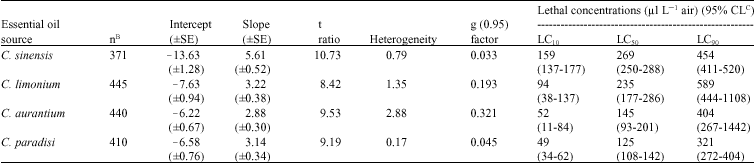 | |
A: Oil applied to 2 cm filter papers held in 27 mL vials. Exposure period was 24 h, B: n = Total number of one-to 2 day old adult insects tested (including control), C: CL = Confidence Limits | |
However, as different concentrations were applied for the four citrus oils, more appropriate comparisons could be obtained using probit analyses of data. The dose-mortality responses of C. maculatus adults to citrus essential oils were compared in terms of differences in slope and/or intercept of probit regressions and LC50s and LC90s. The slope values in fumigant toxicity were in the range of 2.88-5.61. The heterogeneity factors less than 1.0 for C. sinensis and C. paradisi oils indicated that the results were found to be within the 95% confidence limits, so no correction factor (g) was required. The heterogeneity factors more than 1.0 for C. limonium (1.35) and C. aurantium (2.88) required to use g factor for correcting of the respective LC50s values. The regression test (t ratio) was greater than 1.96 for all essential oils and the potency estimation test (g factor) was less than 0.5 at all probability levels for all essential oils, except at 99% level for C. limonium (0.65) and C. aurantium (1.08) (Table 3).
The slopes of the four probit morality regressions for the essential oils were significantly different, as revealed by rejecting the likelihood ratio test of parallelism (χ2 = 24.42, df = 3, p<0.001), suggesting that at least the slope of one of the probit mortality regressions would be significantly different from the others. Further likelihood ratio tests between the paired combinations concerned revealed that the slope of the probit mortality line for the oil of C. limonium was not significantly different from those of C. aurantium (χ2 = 0.496, df = 1, p = 0.481) and C. paradisi (χ2 = 0.025, df = 1, p = 0.876). Similar result was obtained when comparing the slopes of the probit mortality lines between the two latter oils (χ2 = 0.334, df = 1, p = 0.563). In contrast, the slope of the probit mortality line for the oil of C. sinensis was significantly greater than those of C. limonium (χ2 = 14.44, df = 1, p< 0.001), C. aurantium (χ2 = 22.30, df = 1, p<0.001) and C. paradisi (χ2 = 16.62, df = 1, p<0.001) (Table 3).
The intercepts of probit morality regressions for the four essential oils were significantly different, as revealed by rejecting the likelihood ratio test of equality (χ2 = 135.97, df = 6, p<0.001), suggesting that at least the
| Table 4: | LC90s ratios and their 95% confidence limitsA calculated for comparing fumigant toxicity of citrus essential oils to the adults of C. maculatusB |
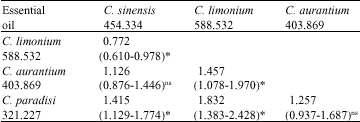 | |
A: The lower and upper 95% CL were calculated according to the procedure described by Robertson and Preisler (1992), B: Italic, bold and bracket-enclosed values show LC90 (μl L-1 air), the ratio of LC90s concerned and its 95% confidence limits, respectively, *: Significant difference between crossed LC90s at 5%; ns: Non-significant | |
| Table 5: | LC50s ratios and their 95% confidence limitsA calculated for comparing fumigant toxicity of citrus essential oils to the adults of C. maculatusB |
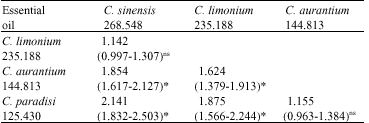 | |
A: Lower and upper 95% CL were calculated according to the procedure described by Robertson and Preisler (1992), B: Italic, bold and bracket-enclosed values show LC50 (μl L-1 air), the ratio of LC50s concerned and its 95% confidence limits, respectively, *: Significant difference between crossed LC50s at 5%; ns: Non-significant | |
intercept of one of the probit mortality regressions would be significantly different from the others. Further likelihood ratio tests of equality indicated that the intercepts between all possible paired combinations significantly differed from each other (p<0.001), except those of C. aurantium and C. paradisi (χ2 = 4.06, df = 2, p = 0.131) (Table 3).
The above differences in parameters of the probit mortality regressions between experimental treatments were reflected in the LC90 or LC50 estimates (Table 3-5). Based on the LC90 values, the essential oil of C. limonium showed the least and that of C. paradisi the highest fumigant toxicity to the adults of C. maculatus. However, comparisons of LC90s among the four essential oils using their ratios and 95% confidence limits indicated that the toxicity of C. sinensis oil did not significantly differ from the toxicity of C. aurantium oil. Also the toxicity of the latter was not significantly different from that of C. paradisi oil Comparison between all other paired oils showed significant differences in their toxicity to C. maculatus adults (Table 4). In contrast, based on the LC50 values, the essential oil of C. sinensis showed the least fumigant toxicity to the adults of C. maculatus, although not significantly different from the oil of C. limonium. Moreover, the fumigant toxicity of C. sinensis oil was significantly lower than that of C. aurantium oil. The results of comparison between the other paired combinations of essential oils were similar to those obtained by LC90 values (Table 5).
DISCUSSION
Over 120 plants and plant products have been shown to have insecticidal or deterrent activity against stored product pests (Dale, 1996). Currently many farmers in parts of Africa and Asia use some of these botanicals to protect their legumes from attack by bruchids, with varying degrees of success (Don-Pedro, 1990; Singh, 1990; Dharmasena, 1995; Dharmasena et al., 1998). However, the number and quality of plants used by farmers is often limited by their availability (Dharmasena, 1995).
Rutaceae is a large family containing 130 genera in seven subfamilies, with many important fruits and essential oil products. Lemon essential oil has the highest value of all essential oils imported to the USA and is widely used as flavoring agent in bakery, as fragrance in perfumery and also for pharmaceutical applications (Weiss, 1997). The essential oils extracted from Citrus genus contain a high percentage of monoterpenes. Its major component is limonene. Analysis of the toxicity data in the present study showed that the essential oil vapors from citrus peels exhibited a variable toxic action against the adult stage of C. maculatus, depending on plant species, concentration and exposure period. Studying on thirteen essential oils related to seven plant families, Papachristos and Stamopoulos (2002) observed varying toxicities among different plant species to the bruchid, A. obtectus. A similar positive relationship between the rate of mortality and exposure time obtained in our study has also been demonstrated in bioassays of many insects with various toxicants (Robertson and Preisler, 1992; Perry et al., 1998; Tunc et al., 2000; Sanon et al., 2002). The essential oil vapor of C. paradisi exhibited the most toxicity to the cowpea adult beetles as demonstrated by the lowest of both LC50 and LC90 estimates at 24 h exposure. Based on the LC50 values, the fumigant toxicity of the essential oils in descending order was C. paradisi, C. aurantium, C. limonium and C. sinensis. Similar results were obtained based on the LC90 values, except for the last two oils in which C. sinensis oil was significantly more toxic than C. limonium oil.
Studying on the effects of essential oils from various plant species on the bruchid, A. obtectus, Papachristos and Stamopoulos (2002) reported that fumigant LC50s values of C. sinensis at 24 h exposure for males and females were 11.4 and 19.5 μl L-1 air, respectively. These values were much lower than the LC50 value 269 μl L-1 of the same plant source obtained in the present study for C. maculatus adults. The observed difference between our results and those of Papachristos and Stamopoulos (2002) seems to be reasonable because of different species and size of insects and/or methodology of oil extraction concerned, as shown in similar experiments with various stored product pests and essential oil vapors (Huang et al., 2000; Sanon et al., 2002). Don-Pedro (1996) attributed the mortality of C. maculatus adults on citrus peel-treated grains to the fumigant activity of the vapor released by peels. He indicated that grains treated by 7 mL oil kg-1 seed caused 100% mortality 1 h after application. LD50 values of orange and grapefruit peels admixed with cowpea grains on adult C. maculatus were 4 and 5.62 g peel per 100 g cowpea, respectively. The LC50 values of C. maculatus adults for citrus essential oil vapors (125-269 μl L-1) in the present study were much lower than the value reported by Sanon et al. (2002) for the oil of Bosica senegalensis Lam. (Capparaceae) (480 μg L-1).
The slope value of probit mortality regression in descending order was C. sinensis, C. limonium, C. paradisi and C. aurantium (Table 3). However, the likelihood ratio tests showed that only was the slope for the essential oil of C. sinensis significantly greater than others and that the slopes for the last three essential oils were similar. A steep slope value indicates that there is a large increase in the mortality of insects with relatively small increase in the concentration of toxicant (Robertson and Preisler, 1992; Perry et al., 1998; Tiwari and Singh, 2004) that was true for the fumigant toxicity of C. sinensis essential oil within the experimental conditions of present study. The lower toxicities (the higher LC50s) of C. sinensis and C. limonium compared to two other species were due to their range of higher concentrations needed for calculations of probit regressions. However, the steep slope of probit mortality regression of C. sinensis was reflected in its higher toxicity to adults when comparison was made in terms of LC90 values, being significantly lower in C. sinensis (454 μl L-1 air) than in C. limonium (589 μl L-1 air) (Table 4). When the value of t-ratio is greater than 1.96, the regression is significant.
Values of heterogeneity factor less than 1.0 denote that in the replicate tests of random samples, the concentration response line would fall within 95% confidence limits and thus the model fits the data adequately, the case that was not true for C. limonium and C. aurantium oils (Table 3). The index of significance of potency estimation g indicates that the value of the LC50 or LC90 was within the limits at 95% probability level (Table 3) as it is less than 0.5 (Robertson and Preisler, 1992; Tiwari and Singh, 2004).
According to the report of Regnault-Roger (1997), the essential oils represent a market estimated at $700 million and a total world production of 45000 t. Nearly 90% of this production is focused on 15 products, particularly mints (Mentha piperita, M. arvensis and M. spicata) and citrus (orange, lemon and lime). The advantages of using citrus peel oil as a grain protectant are: It can be easily extracted from peels by water steam distillation, it may have very low toxicity to mammals since citrus oil is one of the popular flavorings or fragrances (as food ingredients) and consumed by people in various parts of the world, as the essential oil is volatile, it can potentially be used as a fumigant and it is cost-effective and its application is easy. However, as the essential oils are intended to be used like fumigants to disinfest commodities, they should have the ability to kill all stages of insects. Moreover, aromatic plants contain, in general, essential oils at concentrations of 1-3% (Cakir, 1992) so large quantities of plant material have to be processed in order to obtain essential oils in quantities sufficient for commercial scale tests. The observed fumigant activity in the present study shows that essential oils are sources of biologically active vapor that are potentially efficient insecticides. Consequently, the possibility of employing these natural fumigants to control insects in stored products can be worthy of further investigations.
ACKNOWLEDGMENTS
Thanks are due to the Faculty of Agriculture for assistance to conduct these experiments as part of M.Sc. studies of S. Abbar. We thank M. Azizi-Arani and S. Hatefi for their technical support and the anonymous reviewers for their constructive comments on the manuscript.
REFERENCES
- Abbott, W.S., 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol., 18: 265-267.
CrossRefDirect Link - Bell, C.H. and S.M. Wilson, 1995. Phosphine tolerance and resistance in Trogoderma granarium Everts (Coleoptera: Dermestidae). J. Stored Prod. Res., 31: 199-205.
CrossRefDirect Link - Collart, M.G. and W.F. Hink, 1986. Development of resistance to malathion in cat flea (Siphonaptera: Pulicidae). J. Econ. Entomol., 79: 1570-1572.
PubMed - Dharmasena, C.M.D., M.S.J. Simmonds and W.M. Blaney, 1998. Insecticidal activity of eight plant species on egg laying, larval development and adult emergence of Callosobruchus maculatus (f.) in cowpea. Trop. Agric. Res. Ext., 1: 67-69.
Direct Link - Don-Pedro, K.N., 1985. Toxicity of some citrus peels to Dermestes maculatus Deg. and Callosobruchus maculatus (F). J. Stored Prod. Res., 21: 31-34.
Direct Link - Don-Pedro, K.N., 1990. Insecticidal activity of fatty acid constituents of fixed vegetable oils against Callosobruchus maculatus (F.) on cowpea. Pest. Sci., 30: 295-302.
Direct Link - Don-Pedro, K.N., 1996. Fumigant toxicity is the major route of insecticidal activity of citrus peel essential oils. Pest. Sci., 46: 71-78.
Direct Link - Elhag, E.A., 2000. Deterrent effects of some botanical products on oviposition of the cowpea bruchid Callosobruchus maculatus(F.) (Coleoptera: Bruchidae). Int. J. Pest. Manage., 46: 109-113.
Direct Link - Finney, D.J., 1971. Probit Analysis. 3rd Edn., Cambridge University Press, London, UK., pp: 76-80.
CrossRefDirect Link - Hink, W.F. and B.J. Fee, 1986. Toxicity of D-limonene,the major component of citrus peel oil, to all life stages of cat flea, Ctenocephalides felis (Siphonaptera: Pulicidae). J. Med. Entomol., 23: 400-404.
Direct Link - Ho, S.H., Y. Ma and Y. Huang, 1997. Anethole, a potential insecticide fromIllicium verum Hook F., against two stored-product insects. Int. Pest Control, 39: 50-51.
Direct Link - Huang, Y., S.L. Lam and S.H. Ho, 2000. Bioactivities of essential oil from Elletaia cardamomum (L.) Maton. to Sitophilus zeamais Motschusky and Tribolium castaneum (Herbst). J. Stored Prod. Res., 36: 107-117.
CrossRef - Huang, Y., S.H. Ho, H.C. Lee and Y.L. Yap, 2002. Insecticidal properties of eugenol, isoeugenol and methyleugenol and their effects on nutrition of Sitophilus zeamais Motsch. (Coleoptera: Curculionidae) and Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). J. Stored Prod. Res., 38: 403-412.
CrossRefDirect Link - Kassir, J.T., Z.H. Mohsen and N. Mehdi, 1989. Toxic effects of limonene against Culex quinquefasciatus say larvae and its interference with oviposition. Anzeiger fur Schadlingskunde, 62: 19-21.
CrossRef - Keita, S.M., C. Vincent, J.P. Schmit, J.T. Arnason and A. Belanger, 2001. Efficacy of essential oil of Ocimum basilicum L. and O. gratissimum L. applied as an insecticidal fumigant and powder to control Callosobruchus maculatus (Fab.) [Coleoptera: Bruchidae]. J. Stored Prod. Res., 37: 339-349.
CrossRefDirect Link - Nahdy, M.S., R.H. Ellis, S.N. Silim and J. Smith, 1998. Field infestation of pigeonpea Cajanus cajan (L.) Millsp. by Callosobruchus chinensis (L.) in Uganda. J. Stored Prod. Res., 34: 207-216.
Direct Link - Obeng-Ofori, D. and C. Reichmuth, 1997. Bioactivity of eugenol, a major component of essential oil of Ocimum suave (Wild.) against four species of stored-product Coleoptera. Int. J. Pest Manage., 43: 89-94.
CrossRef - Obeng-Ofori, D., C.H. Reichmuth, J. Bekele and A. Hassanali, 1997. Biological activity of 1,8 cineole, a major component of essential oil of Ocimum kenyense (Ayobaugira) against stored product beetles. J. Applied Entomol., 121: 237-243.
CrossRefDirect Link - Olubayo, F.M. and G.R. Port, 1997. The efficacy of harvest time modification and intercropping as methods of reducing the field infestation of cowpeas by storage bruchids in Kenya. J. Stored Prod. Res., 33: 271-276.
CrossRefDirect Link - Onu, I. and A. Sulyman, 1997. Effect of powdered peels of citrus fruits on damage: Seeds by Callosobruchus maculatus(F.) to cowpea. J. Sustain Agric. Environ., 9: 85-92.
Direct Link - Papachristos, D.P. and D.C. Stamopoulos, 2002. Repellent, toxic and reproduction inhibitory effects of essential oil vapours on Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae). J. Stored Prod. Res., 38: 117-128.
CrossRefDirect Link - Rahman, M.M. and G.H. Schmidt, 1999. Effect of Acorus calamus (L.) (Araceae) essential oil vapours from various origins on Callosobruchus phaseoli (Gyllenhal) (Coleoptera: Bruchidae). J. Stored Prod. Res., 35: 285-295.
CrossRefDirect Link - Regnault-Roger, C., 1997. The potential of botanical essential oils for insect pest control. Integ. Pest. Manage. Rev., 2: 25-34.
CrossRefDirect Link - Russell, R.M., J.L. Robertson and N.E. Savin, 1977. POLO: A new computer program for probit analysis. Bull. Entomol. Soc. Am., 23: 209-213.
CrossRefDirect Link - Sanon, A., M. Garba, J. Auger and J. Huignard, 2002. Analysis of the insecticidal activity of methylisothiocyanate on Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) and its parasitoid Dinarmus basalis (Rondani) (Hymenoptera: Pteromalidae). J. Stored Prod. Res., 38: 129-138.
CrossRefDirect Link - Shaaya, E., U. Ravid, N. Paster, M. Kostjukovsky, M. Menasherov and S. Plotkin, 1993. Essential oils and their components as active fumigants against several species of stored-product insects and fungi. Acta Hortic., 344: 131-137.
Direct Link - Tiwari, S. and A. Singh, 2004. Piscicidal activity of alcoholic extract of Nerium indicum leaf and their biochemical stress response on fish metabolism. Afr. J. Tradit. Complementary Alter. Med., 1: 15-29.
Direct Link - Tunc, I., B.M. Berger, F. Erler and F. Dagli, 2000. Ovicidal activity of essential oils from five plants against two stored-product insects. J. Stored Prod. Res., 36: 161-168.
CrossRef